三江源地区高寒草地群落特征研究
2010-05-30杜岩功崔骁勇葛劲松赵旭东王勇
杜岩功,崔骁勇,葛劲松,赵旭东,任 杰,王勇
(1.中国科学院研究生院资源与环境学院,北京100049;2.青海省环境监测中心站,青海 西宁810007)
生物多样性与全球变化是目前各国面临的主要生态、环境问题[1]。生物多样性研究主要集中于物种多样性[2],它是群落生态组织水平的主要特征之一,也是群落生态环境优劣的重要指标[3],物种多样性较高,系统受到外界干扰时有较大的反馈调节能力,生态系统稳定性高。
高寒草地是青藏高原的主体类型之一,近年来由于长期超载放牧,造成高寒草地严重退化、沙化,生物多样性降低,草地产草力下降,草地生态环境日趋恶化[4]。因此,青藏高原高寒草地群落特征研究、高寒草地退化和恢复治理研究尤为迫切。目前有关江河源区高寒草地多样性研究主要集中在荒漠化草地和紫花针茅草原群落特征[5-6]及高寒草地不同放牧强度下演替过程、特征。结果认为:高寒草甸草地生产力高,牧草品质优良,物种多样性和丰富度指数较高;物种多样性受海拔、纬度、温度和湿度等因素影响[1];随着放牧干扰强度增加,物种多样性和丰富度指数逐渐降低,草地生物量降低[6-7]。
本研究在三江源地区选择典型的高寒草原和高寒草甸样地,通过对其生物多样性和分布格局研究,深入分析该地区群落特征,为该地区生物多样性保护和生态恢复提供科学依据。
1 材料与方法
1.1 试验区自然概况 三江源地区平均海拔4 000 m以上,气候寒冷属高寒大陆性气候,只有冷暖两季,没有四季之分,冷季持续7~8个月,全年无绝对无霜期。年降水量420~560 mm,多集中在5-10月,植物生长季雨热同期。草地类型主要以针茅草原和高寒草甸为主,土壤类型主要为高山草甸土、高山草原土[7]。
优势物种为高山嵩草Kobresia pygmaea、短花针茅 Stipabrevi f lora、紫花针茅S.purpurea、青海固沙草Orinus kokonorica,伴生种主要有二裂委陵菜Potentilla bi f urca、早熟禾Poa indattenuata、苔草Carex ivanovae、矮火绒草 Leontopodium nanum 、棘豆 Oxytropisochrocephala(见表1)。
1.2 试验设计与方法 2008年8月在三江源区选择针茅草原和高山嵩草草甸的6处典型样地,进行植物群落调查、土壤湿度测定。地上生物量采用标准收获法,群落调查样方面积1 m×1 m;0~10 cm土壤湿度采用TDR(时域反射仪)测定,3次重复。α多样性用丰富度指数、物种多样性指数和均匀度指数3类多样性指数表征[8]。
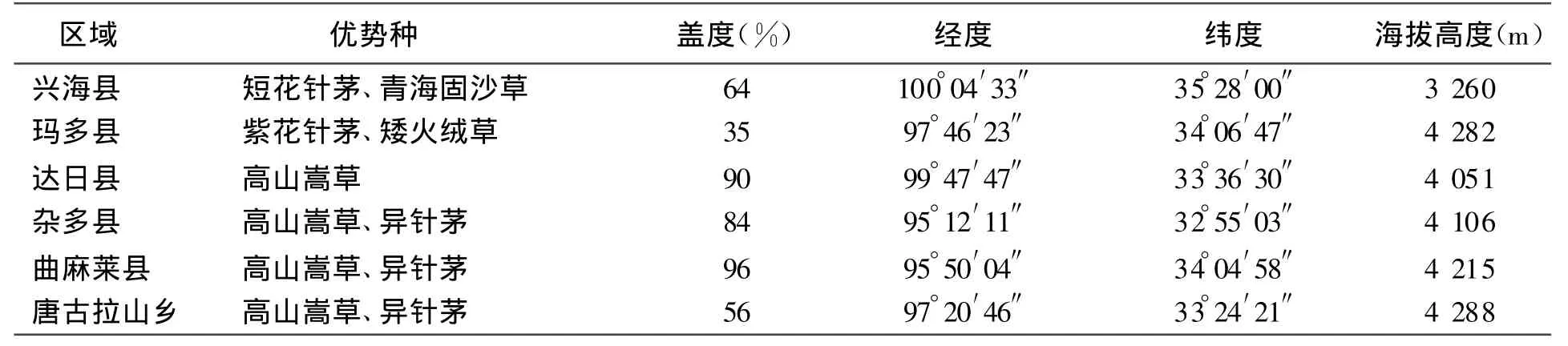
表1 样地基本情况
丰富度指数R=S
物种多样性指数计算采用 Shannon-Wiener指数:

式中,Pi为种i的相对重要值:Pi=(相对高度+相对盖度)/2;S为种i所在样方的物种总数。
1.3 数据分析 不同样地土壤湿度、物种数差异采用单因素方差分析;物种多样性和物种数采用回归分析;群落特征进行聚类分析;可食用鲜草产量与土壤湿度、物种数目、物种多样性和海拔采用相关性分析(SPSS 16.0)。
2 结果与分析
2.1 不同样地土壤湿度 高寒草地的水分来源主要依靠降水[9],土壤湿度是影响植物生长的重要环境因子。高寒草甸土壤湿度显著高于高寒草原土壤(兴海县、玛多县,P<0.05)。曲麻莱县土壤湿度最高,达到42.82%,唐古拉山乡最低,为21.27%(P<0.05),杂多县和达日县土壤湿度一致(P>0.05);兴海县短花针茅草原土壤湿度为12.63%,而玛多县紫花针茅草原土壤湿度仅为7.80%,二者之间达到显著性差异水平(P<0.05,图1)。这可能是因为高寒嵩草草甸致密的草皮层具有较大的水分保蓄能力[10]。贺金生等[11]认为从大尺度来看,水热条件及其组合是决定植物群落分布的主要因素。
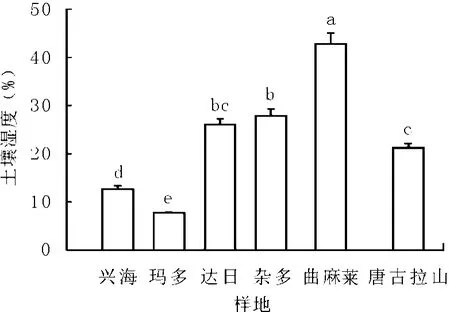
图1 高寒草地土壤湿度比较
2.2 不同植物群落的α多样性特征 物种丰富度是群落多样性的最基本特征。达日县和曲麻莱县高寒草甸物种数较高,分别为20种和15种,二者之间达到显著性差异水平(P<0.05);兴海县、玛多县和唐古拉山乡草地物种数较少,均是8种,杂多县物种数最低,仅有5种(P>0.05,图2)。物种数与海拔和土壤湿度之间相关性不显著(P>0.05)。杂多县物种数最低,但其海拔不是最高,土壤湿度也不是最低,并且纬度最低,这说明物种丰富度可能是受温度、降水、海拔和放牧利用强度等因素共同作用影响。岳鹏鹏等[5]2005年调查结果认为玛多县紫花针茅草原物种数为12~26,物种数高于本研究,这可能是由于不同年份和采样地点受放牧干扰造成。李凯辉等[12]发现新疆天山高寒草原和草甸植物物种约有11~16种,这与本研究结果一致。
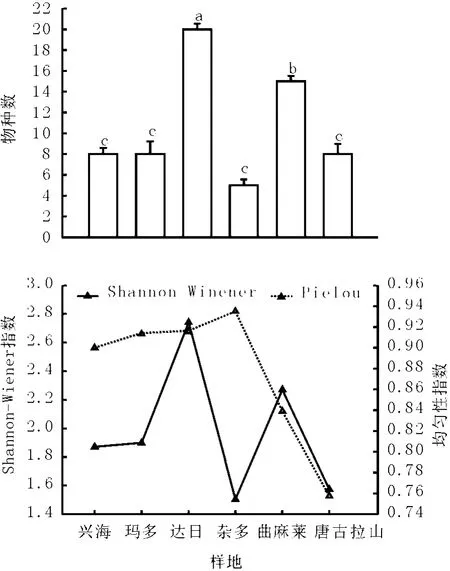
图2 高寒草地物种丰富度和物种多样性指数
物种多样性是生物多样性的核心内容之一,其在生物多样性保护中具有重要意义[1]。达日县和曲麻莱县高寒草甸物种多样性最高,分别为2.74和2.27;唐古拉山乡和杂多县最低,约为1.5;兴海县和玛多县针茅草原物种多样性约为1.87。均匀度指数唐古拉山乡和曲麻莱县稍低,分别为0.76和0.84,其余各县均匀度指数均较高(图2)。杨元武等[1]在祁连山高寒草甸研究结果认为多样性为0.65~3.37,而Wang等[8]在果洛州高寒草甸中发现物种多样性为2.8~3.5;岳鹏鹏等[5]认为紫花针茅草原物种多样性为2.2左右。
物种多样性的纬度格局,一般表现为随着纬度降低,物种多样性增加[11]。在针茅草原和高寒嵩草草甸中这种规律都不明显,其中杂多县纬度最低,但物种多样性也最低(图2),岳鹏鹏等[5]认为在玛多县紫花针茅草原物种多样性指数与纬度呈显著负相关关系。Currie[13]认为在美国东部物种多样性变化存在明显的纬度梯度,而在西部这种特征表观不明显,这可能是由于地形、温度差异等原因造成[11]。物种多样性随着海拔梯度变化规律很复杂[11],本研究发现物种多样性与海拔之间无显著相关性(P>0.05),随海拔增加,物种多样性变化规律不明显。这与杨元武等[1]、杨立军等[14]在祁连山和达日县高寒草地研究结果一致,这表明在低温、高海拔的地区高寒草地物种多样性可能受多种因素影响。
三江源地区平均海拔在4 000 m以上,生态系统脆弱,地形、温度和湿度差异会成为生物多样性增加的主要限制因素。
6处样地统计分析发现物种多样性和物种数极显著相关(r=0.967,P<0.01),这与 Wang等[8]和胡玉昆等[15]研究结果一致。在三江源地区高寒草地多样性指数拟合方程为y=1.130 1+0.079 5x(R2=0.935 9,P <0.01,图 3),用物种数可以较好地拟合物种多样性指数。
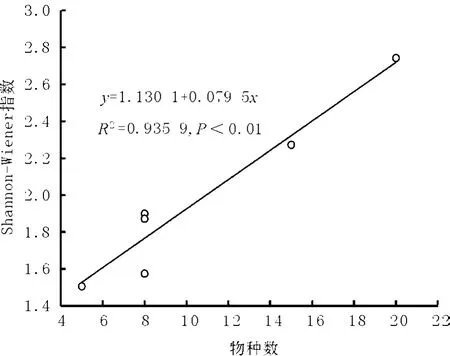
图3 高寒草地Shannon-Wiener指数与物种数目拟合方程
2.3 高寒草地鲜草量和地下生物量比较可食用鲜草量是草地承载力的重要指标,高寒草甸鲜草生产力均高于针茅草原,其中达日县高山嵩草草甸产草量最高,约为466.8 g/m2,其次是杂多县、曲麻莱县和唐古拉山乡高寒草甸,玛多县紫花针茅草原鲜草产量较低,约为71.0 g/m2(图4)。统计结果发现可食用鲜草量与土壤湿度、海拔、物种数没有显著相关性(P>0.05)。总鲜草量和地下生物量与可食用鲜草量表现出一致规律。达日县高寒草甸地下生物量最高,为3 916.7 g/m2,且鲜草产量最低的玛多针茅草原地下生物量也达到733.3 g/m2。这可以看出高寒草地植物根系发达,生物量高,这与杜岩功等[10]在海北州高寒草甸研究结果一致。杜利霞等[16]发现放牧会增加冷蒿Atemisia f rigida地下部分生物量而减少地上部分生物量,这可以使其回避被采食。董全民等[17]看到高山嵩草和早熟禾生态位较宽,而紫羊茅Festuca rubra和青海野青茅D.kokonorine生态位很窄。刘发央等[18]发现适度的放牧干扰抑制草地中优势种的竞争作用,为其他物种提供潜在生态位。高寒草甸根系致密,并能发育出其特有的草毡表层,可耐放牧家畜的践踏,是维持高寒草甸生态系统稳定性的重要生态屏障[10],这可能也是高寒草甸植物长期对环境适应的生态策略。
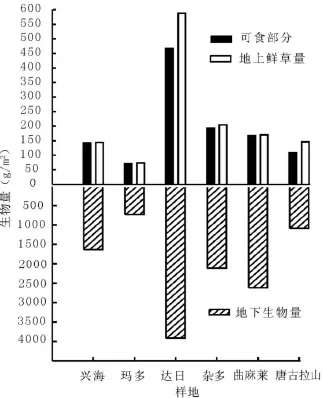
图4 高寒草地不同样地生物量
3 讨论
不同地区高寒草地群落在盖度、高度、植物种类组成、重要值和多样性等方面都变现出一定的差异。这种分布特征,可能是因为对地带性气候、地理隔离等较敏感[5]。
聚类分析是将研究样品或者变量之间相似性程度大的聚为一类,把另外一些相似性大的聚为另一类,然后继续分别聚类,最终聚为一个大类,结果采用直观的聚类树状分类图表示。重要值是反映物种在群落中重要性的一个主要指标[4],根据样方调查数据,按照群落物种重要值进行聚类分析发现,杂多县和曲麻莱县高山嵩草-异针茅草甸首先被归为一类,这二者盖度也基本一致;其次将唐古拉山乡高山嵩草草甸合为一类;然后依次是兴海县短花针茅草原、达日县高寒嵩草草甸;玛多县紫花针茅草原最后被聚合为一类,这可能是由于其物种数和盖度均较低。聚类分析的结果和样方调查数据中的优势种和草地类型分类结果一致(表1,图5)。岳鹏鹏等[5]对江河源区紫花针茅草原的不同地区样地群落进行研究也发现聚类分析结果和草地类型分类相当一致,本研究也发现该地区紫花针茅草原分布呈现出小尺度同质性,大尺度异域异质性特征。孙海松[19]对青海省21个草地类型进行聚类分析发现21类草地可以分为12类,认为用这种定量方法分类结果合理。张静等[20]采用聚类分析方法将三江源区12块退化草地分为9类,看到草地之间有较大差异,治理上应采取不同措施。
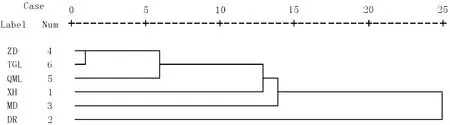
图5 高寒草地群落聚类分析
人类活动干扰和过度放牧,以及气候干暖化是造成草地退化的重要原因[21]。随着主要以根茎繁殖的莎草科植物物种数减少,草皮层坏死,冻融过程和鼠害同时加速草皮层剥蚀,最终造成土层裸露,土壤随雨水发生大量流失[20]。对处于轻度和中度干扰高寒草地,需要通过降低或者解除放牧压力,以及补播、耙松等措施,使草地得到休养生息,保持草毡表层在一定限度,既可以使草地保持较高生产力又可以缓解草地的退化[10,22-23]。而对于已经出现草皮的塌陷和斑驳等重度、极度退化高寒草地,采用建植栽培草地,短期内可实现草地植被的恢复,张耀生等[24]通过连续13年试验发现,建植栽培草地可以有效逆转高寒草地退化趋势。
4 主要结论
4.1 高寒草甸土壤湿度显著高于高寒草原土壤(P<0.05),曲麻莱县土壤湿度最高,唐古拉山乡最低(P<0.05),兴海县短花针茅草原土壤湿度约为12.63%,而玛多县紫花针茅草原土壤湿度仅为7.80%,二者之间达到显著差异(P<0.05)。
4.2 达日县和曲麻莱县高寒草甸物种数较高,分别为20种和15种,二者之间达到显著性差异水平(P<0.05)。兴海县、玛多县和唐古拉山乡草地物种数都是8种,杂多县物种数最低仅有5种(P>0.05)。物种数与海拔和土壤湿度均没有显著相关性(P>0.05)。
4.3 达日县和曲麻莱县物种多样性最高,分别为2.74和2.27;唐古拉山乡和杂多县最低,约为1.5;兴海县和玛多县针茅草原物种多样性约为1.87。均匀性指数唐古拉山乡和曲麻莱县稍低,分别为0.76和0.84,其余各县均匀性指数均较高。多样性指数随纬度和海拔变化规律不明显,但与物种数目极显著相关(r=0.967,P<0.01),拟合方程为 y=1.130 1+0.079 5x(R2=0.935 9,P<0.01)。
[1] 常学向,赵文智,赵爱芬.祁连山区不同海拔草地群落的物种多样性[J].应用生态学报,2004,15:1599-1603.
[2] Tracy C,Brussard P.Preserving biodiversity:Species in landscapes[J].Ecology Application,1994(4):205-207.
[3] 刘振国,李镇清.退化草原冷蒿群落13年不同放牧强度后的植物多样性[J].生态学报,2006,26:475-480.
[4] 杜岩功,曹广民,王启兰,等.放牧对高寒草甸地表特征和土壤物理性状的影响[J].山地学报,2007,25:338-343.
[5] 岳鹏鹏,卢学峰,叶润蓉,等.江河源不同区域紫花针茅草原群落特征[J].植物生态学报,2008,32:1116-1125.
[6] 杨元武,李希来,祁盛仓.江河源地区不同荒漠化草地物种多样性研究[J].青海大学学报(自然科学版),2005,23:42-45.
[7] 张静,李希来,于海.青藏高原不同退化程度小嵩草草甸群落结构特征与土壤理化特征分析[J].草原与草坪,2008(4):5-9.
[8] Wang C,Long R,Wang Q,et al.Effects of altitude on plant-species diversity and productivity in an alpine meadow,Qinghai-Tibetan plateau[J].Australian Journal of Botany,2007,55:110-117.
[9] 曹广民,李英年,鲍新奎.高寒地区寒冻雏形土的持水特性[J].土壤,1998(1):27-31.
[10] 杜岩功,梁东营,曹广民,等.放牧强度对嵩草草甸草毡表层及草地营养和水分利用的影响[J].草业学报,2008,17(3):146-150.
[11] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17:91-99.
[12] 李凯辉,胡玉昆,王鑫,等.不同海拔梯度高寒草地地上生物量与环境因子关系[J].应用生态学报,2007,18:2019-2024.
[13] Currie D,Paquin V.Large-scale biogeographical pattern of species richness of trees[J].Nature,1987,329:326-327.
[14] 杨力军,李希来,石德军,等.青南高海拔地区高寒草甸植物群落多样性的研究[J].草原与草坪,2000(2):32-35.
[15] 胡玉昆,李凯辉,柳妍妍,等.天山南坡高寒草地海拔梯度上的植物多样性变化格局[J].生态学杂志,2007,26:182-186.
[16] 杜利霞,李青丰,董宽虎.放牧强度对短花针茅草原冷蒿繁殖特性的影响[J].草地学报,2007,15:367-370.
[17] 董全民,赵新全,马玉寿,等.放牧对高寒小嵩草草甸冷季草场主要植物种群生态位的影响[J].中国草地学报,2006,28:10-17.
[18] 刘发央,徐长林,龙瑞军.牦牛放牧强度对金露梅灌丛草地群落物种多样性的影响[J].草地学报,2008,16(6):613-618.
[19] 孙海松.应用聚类分析法对青海21个草地型分类的初步探讨[J].青海大学学报(自然科学版),2003,21(6):7-10.
[20] 张静,李希来,谢得雄.三江源地区不同退化草地聚类分析[J].草业科学,2008,25(6):8-13.
[21] 王建兵,王振国,吕虹.黄河重要水源补给区草地退化的气候背景分析[J].草业科学,2008,25(4):23-27.
[22] 曹广民,龙瑞军.三江源区黑土滩型退化草地自然恢复的瓶颈及解决途径[J].草地学报,2009,17(1):4-9.
[23] 崔庆虎,蒋志刚,刘季科,等.青藏高原草地退化原因述评[J].草业科学,2007,24(5):20-26.
[24] 张耀生,赵新全,黄德清.青藏高寒牧区多年生人工草地持续利用的研究[J].草业学报,2003,12(3):22-27.
