褪黑素对模拟干旱胁迫下罗勒幼苗光合性能及抗氧化酶活性的影响
2024-04-08郄亚微贺涛
郄亚微 贺涛

摘要:为了研究干旱胁迫下施加褪黑素对罗勒幼苗光合性能及抗氧化酶活性的影响,以罗勒幼苗为试验材料,设置对照(CK)、干旱胁迫(DR)、褪黑素预处理+干旱胁迫(ME+DR)3种处理,在干旱胁迫7 d后测定叶片的光合生理参数、根系形态参数、抗氧化酶活性。结果表明,DR处理显著抑制罗勒幼苗的光合作用,影响根系形态,降低光合色素含量;同时,DR处理下罗勒幼苗的可溶性糖、脯氨酸、过氧化氢、丙二醛、超氧阴离子含量均有所提高。与DR处理相比,褪黑素可通过恢复叶绿素含量、根系结构、气体交换参数,改善DR处理造成的不利影响。此外,外源褪黑素可增强超氧化物歧化酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的活性,降低相对电导率,通过调节超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、谷光甘肽S-转移酶、脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的相对基因表达,对罗勒幼苗进行膜质过氧化水平的稳定和渗透调节。综上所述,褪黑素可以有效改善罗勒幼苗在干旱胁迫下的根系特征、叶片光合作用、抗氧化机制,从而提高幼苗对干旱胁迫的适应性。
关键词:罗勒;光合作用;根系生长;氧化损伤;褪黑素;干旱
中图分类号:S636.901 文献标志码:A
文章编号:1002-1302(2024)03-0179-05
我国的干旱、半干旱地区约占国土面积的26.73%,干旱已经成为限制植物生长、产量和品质的重要因素之一[1]。研究表明,干旱胁迫会导致植物色素含量降低、离子稳态失衡、蒸腾作用降低、气孔关闭、细胞增大减小,最终导致植物死亡[2-4]。在缺水条件下,植物活性氧的过度积累会引起氧化损伤,从而导致细胞器严重损伤[5]。活性氧分子包括H2O2、O-2·等,与细胞膜系统相互作用,主要破坏细胞中存在的大分子[6]。活性氧过量积累,导致电子传递链氧化损伤,叶绿体、线粒体脂质过氧化增强,酶活性、蛋白质、核酸失活,最终降低作物的光合作用和产量[7]。抗氧化酶、非酶抗氧化剂分子有效减少活性氧的积累,从而平衡活性氧的合成和信号,增强植物的耐干旱胁迫能力[8]。此外,渗透物的积累也有助于抵抗干旱胁迫,防止植物的氧化膜损伤[9]。全球气候变暖显著影响了作物产量,其中干旱胁迫是主要因素之一。为了有效避免干旱所造成的危害,在农业生产中通常使用植物激素或生物刺激剂等农艺措施来提高植物的耐旱性,促进生理、生化和分子过程,对植物的生长调节和恢复力产生积极影响。因此,研究植物激素的作用机制,对提高植物抗逆性具有重要意义。
褪黑素是一种天然、多功能、无毒、具有调节性的吲哚类激素物质[10]。迄今为止,褪黑素被证明可以增强植物对不同非生物胁迫的抗性,如干旱、热、冷、盐、重金属、化学物质、病原体[11-13]等。此外,褪黑素在细胞和组织水平上促进植物根系形态、种子萌发、光合作用、幼苗生长,延缓叶片衰老,提高抗氧化效率,促进果实成熟[11-13]。具体而言,活性氧稳态和抗氧化能力的增强,与干旱胁迫下幼苗中较高的褪黑素含量有关[14-15]。前人研究表明,在干旱胁迫下,外源褪黑素可增强小麦的植株生长属性[16],改善生菜的根结构系统[15],提高菊花的光合效率[13],降低大豆的脂质过氧化和活性氧积累[17],改善燕麦的抗氧化酶系统[12]。
罗勒(Ocimum basilicum L.)为唇形科罗勒属药食兼用芳香植物,广泛分布于热带与亚热带地区[18]。缺水对罗勒的生理、形态、生化、解剖结构都有显著的抑制作用[19]。在水分亏缺条件下,有关褪黑素如何调节罗勒植物的生理、形态、生化的机制目前尚不清晰。本试验通过研究干旱胁迫下,褪黑素对罗勒幼苗光合性能、膜损伤、根系结构、叶绿体结构、活性氧稳态的影响,探索褪黑素在增强罗勒干旱耐受性方面的调控机制,旨在为罗勒植物耐旱性评价和旱地引种栽培提供科学参考。
1 材料与方法
1.1 试验设计
试验于2022年在中国热带农业科学院热带作物品种资源研究所南药圃进行,以绿叶罗勒 (Ocimum basilicum Improved Genovese Compact)為试材,种子经5% NaClO消毒20 min,蒸馏水洗净,置于铺有湿润滤纸的培养皿内。种子露白后,将其播于含有草炭、蛭石、珍珠岩混匀的塑料盆(内径 18 cm、高11 cm) 中。待幼苗长至3张真叶时,充分灌溉后停水(土壤绝对含水量为40%~45%),水分胁迫组为自然干旱,试验设置3个处理:CK,正常供水;DR处理,干旱胁迫,置于20% PEG-6000溶液中处理7 d;MT+DR处理,褪黑素预处理+干旱胁迫,在含100 μmol/L褪黑素培养液中预处理 72 h,置于20% PEG-6000溶液中处理7 d。每个处理重复 3 次,试验重复2次。干旱胁迫7 d后,取罗勒幼苗,进行光合生理指标的测定。
1.2 测定指标及方法
1.2.1 光合特征参数 采用可见光分光光度法分别在645、663、440 nm处测量吸光度,计算叶绿素a、叶绿素b 、类胡萝卜素含量;利用 Li-6400XT型便携式光合仪测定光合参数,包括净光合速率、胞间CO2浓度、气孔导度、蒸腾速率。
1.2.2 渗透调节物质和膜脂过氧化水平 采用酸性茚三酮比色法测定游离脯氨酸含量;采用蒽酮比色法测定可溶性糖含量;采用硫代巴比妥酸法测定丙二醛含量;用DDS-307电导率仪测量叶片电导率;过氧化氢、超氧阴离子含量的测量参照郭欣欣等的方法[20]。
1.2.3 抗氧化酶活性 参照南京建成试剂盒中的操作指南,记录超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、谷光甘肽S-转移酶、脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶分别在550、405、290、340、412、420、412、340 nm 波长处的活性。
1.2.4 根系形态参数 干旱胁迫7 d后,取3株罗勒幼苗根系,用自来水冲洗干净,使用根系扫描仪扫描根系,然后使用Win RHIZO根系分析系统得到根系形态参数,包括根长、根体积、根表面积、根尖数、根系平均直径。
1.3 统计分析
使用SPSS 22.0統计软件对数据进行单因素方差分析(ANOVA),并使用LSD(最小显著性差异)检验有无显著性(α=0.05)。
2 结果与分析
2.1 褪黑素对干旱胁迫下罗勒幼苗光合色素含量的影响
由表1可见,与CK相比,DR胁迫处理下罗勒幼苗叶片中的叶绿素a、叶绿素b、类胡萝卜素含量分别显著降低62.04%、47.54%、66.67%;ME+DR处理下,叶片叶绿素a、叶绿素b、类胡萝卜素含量较CK分别显著降低21.30%、19.67%、29.73%,但比DR处理分别显著提高107.32%、53.13%、110.81%。
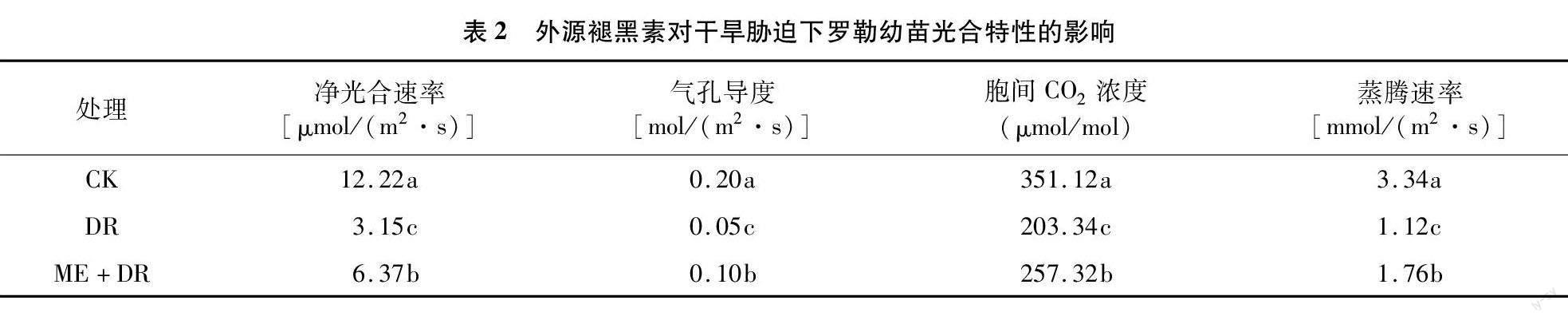
2.2 褪黑素对干旱胁迫下罗勒幼苗光合参数的影响
由表2可见,与CK相比,DR胁迫处理下罗勒幼苗叶片净光合速率、气孔导度、胞间CO2浓度、蒸腾速率分别显著降低74.22%、75.00%、42.09%、66.47%;ME+DR处理下,上述参数分别比CK降低47.87%、50.00%、26.71%、47.31%,但比DR处理分别显著提高102.22%、99.98%、26.55%、57.14%。
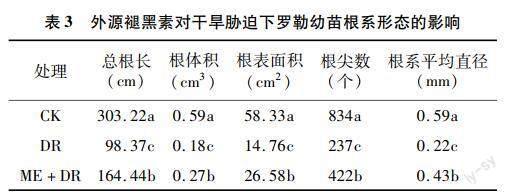
2.3 褪黑素对干旱胁迫下罗勒幼苗根系形态的影响
由表3可见,与CK相比,DR处理显著降低罗勒幼苗根系形态的参数值,总根长、根体积、根表面积、根尖数、根系平均直径分别显著降低67.56%、69.49%、74.70%、71.58%、62.71%;ME+DR处理下,上述根系形态参数分别比CK降低45.77%、54.24%、54.43%、49.40%、27.12%,但与DR处理相比分别显著增加67.16%、50.00%、80.08%、78.06%、95.45%。
2.4 褪黑素对干旱胁迫下罗勒幼苗渗透调节物质的影响
由图1可见,与CK相比,DR处理下叶片的脯氨酸、可溶性糖含量分别显著增加130.54%、49.55%;ME+DR处理下,脯氨酸、可溶性糖含量分别比CK增加85.74%、21.50%,但与DR处理分别显著降低19.44%、18.76%。
2.5 褪黑素对干旱胁迫下罗勒幼苗膜脂过氧化物的影响
由表4可见,与CK相比,DR处理下罗勒幼苗过氧化氢含量、超氧阴离子含量、丙二醛含量、相对电导率分别显著增加131.36%、142.11%、122.28%、31.13百分点;ME+DR处理下,上述参数分别比CK增加76.58%、102.63%、68.27%、17.88百分点,但比DR处理分别显著降低23.68%、16.30%、24.30%、13.25百分点。
2.6 褪黑素对干旱胁迫下罗勒幼苗抗氧化酶活性的影响
由表5可见,在DR胁迫下,超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶的活性较CK分别提高56.08%、83.72%、87.89%、8.97%、8.77%,而谷光甘肽 S-转移酶活性较CK降低26.35%;ME+DR处理下,超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶的活性较CK分别提高93.09%、90.70%、124.55%、40.79%、148.61%,而谷光甘肽S-转移酶活性较CK降低12.72%。与DR胁迫处理相比,ME+DR处理下,超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、谷光甘肽S-转移酶的活性分别提高23.72%、3.80%、19.51%、29.21%、33.16%、18.50%。此外,与CK相比,DR胁迫处理幼苗的脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的活性分别显著提高26.81%、27.70%。与DR胁迫处理相比,ME+DR处理下脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶活性分别提高33.71%、16.94%。
2.7 褪黑素对干旱胁迫下罗勒幼苗抗氧化酶基因相对表达量的影响
由表6可见,在DR处理下,超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶的相对基因表达量较CK分别降低了8.31%、53.21%、36.42%、0.99%、7.03%,而谷光甘肽S-转移酶的相对基因表达量较CK降低了74.75%。与DR胁处理相比,ME+DR处理下,超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、谷光甘肽S-转移酶的相对基因表达量分别提高144.30%、94.12%、73.37%、114.57%、65.55%、73.99%。此外,与CK相比,DR胁迫处理下,幼苗的脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的相对基因表达量分别显著提高59.60%、3.50%。与DR胁迫处理相比,ME+DR处理下,脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的相对基因表达量分别提高103.80%、126.09%。
3 讨论
光合作用是对干旱胁迫高度敏感的生理过程之一,干旱对光合作用的生物能过程有不利影响[2-5]。本研究结果表明,在干旱胁迫下,经褪黑素预处理的罗勒幼苗,其叶片净光合速率、气孔导度、胞间CO2浓度、蒸腾速率均显著得到改善;这与前人在大豆[2]、银杏[4]、燕麦[5]等植物上的结论相一致。植物受到干旱胁迫时,会降低叶片气孔导度,从而减少水分散失,同时CO2吸收也受到影响,最终导致光合速率降低[21-22]。而经褪黑素预处理后,植物可以对干旱环境进行较快的响应,以减少干旱对植物光合系统及其功能造成的损伤[16,23]。光合色素是植物进行光合作用过程所必需的元素[24]。本研究结果表明,DR处理使罗勒幼苗叶片中的叶绿素a、叶绿素b、类胡萝卜素显著降低;施用褪黑素显著提高了罗勒幼苗的光合色素含量,这与前人在小麦上的研究结果[16]一致。由于光合作用能力得到增强,植物获得了更多的能量,这使它们能够应对干旱所造成的的环境压力[25]。
根系不仅为植物的地上部分提供结构支撑,而且还提供营养和水分[26]。因此,植物的生存取决于其适当的生长发育和根系功能。本研究结果表明,干旱胁迫显著降低了罗勒幼苗的根表面积、根体积、根长度、根尖数、根系平均直径;经褪黑素处理的罗勒幼苗根系形态参数明显增强,这与前人在番茄上的研究结果[27]一致。其主要原因是,经褪黑素预处理的植物修复了被破坏的根系,从而维持了根系的正常功能。同时,外源褪黑素的施用可以改善非生物胁迫所造成的根系损伤[26]。
在干旱胁迫下,脯氨酸、可溶性糖作为重要的渗透调节物质,可以提高植物的抗逆性[9]。前人研究表明,干旱胁迫下番茄[28]、山核桃[29]叶片含水量降低,导致脯氨酸、可溶性糖含量增加,本研究结果与之是一致的,说明干旱胁迫条件下,作物通过增加渗透调节物质来维持细胞渗透势平衡。逆境条件下,活性氧过度积累,导致脂质过氧化,损害细胞膜结构,从而危害植物的生长[7]。在本研究中,干旱胁迫下罗勒幼苗的氧化损伤水平(过氧化物含量、氧离子含量、丙二醛含量、电导率)显著增加;外源褪黑素可以有效保护植物细胞免受氧化损伤,这与前人在冬小麦上的研究结果[7]一致。由此可见,褪黑素可以通过平衡活性氧过度积累,来减少DR引起的氧化损伤,并修复受损细胞膜。
抗氧化酶在植物抵御生物、非生物胁迫的防御系统中起着重要作用[30-32]。本研究结果表明,DR胁迫显著提高了罗勒幼苗叶片的超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的活性及其相关基因表达,这与DR胁迫对黑麦草、苜蓿抗氧化酶的的影响结果[30]类似。褪黑素被认为是一种普遍的抗氧化剂,它能够通过清除过量的活性氧和维持氧化还原稳态来降低逆境胁迫对植物的损伤[10,33]。在本研究中,褪黑素预处理的罗勒幼苗,其超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶、谷胱甘肽还原酶、脱氢抗坏血酸还原酶、单脱氢抗坏血酸还原酶的活性和相对基因表达量上调,说明褪黑素可显著增强干旱胁迫下各种植物抗氧化酶的活性。此外,褪黑素的应用可提高抗氧化系统的性能,激发抗氧化酶的活性[34]。
4 结论
干旱胁迫下,由于罗勒幼苗光合性能受损,膜脂过氧化产物过量积累,渗透失衡,根系形态受损,导致抗旱能力显著下降。相反,外源褪黑素能有效缓解干旱胁迫,减少膜脂过氧化损伤,提高渗透调节能力,增强根系结构,提高光合性能。此外,干旱胁迫下,经褪黑素预处理的罗勒幼苗的可溶性糖、脯氨酸含量显著下降,说明外源褪黑素是一种有效保护剂,可以通过增强抗氧化酶活性和减少氧化损伤,来提高罗勒幼苗的耐旱能力。
参考文献:
[1]王 硕,贾潇倩,何 璐,等. 作物对干旱胁迫的响应机制及提高作物抗旱能力的调控措施研究进展[J]. 中国农学通报,2022,38(29):31-44.
[2]王兴荣,张彦军,陈光荣,等. 干旱胁迫对大豆光合、产量及品质的影响[J]. 干旱地区农业研究,2023,41(2):150-159.
[3]穆若兰,刘旻霞,徐 璐,等. 黄土高原半干旱区典型植物资源利用效率及光合日变化探析[J]. 植物生理学报,2022,58(7):1381-1391.
[4]肖石红,孙红斌,张卫强,等. 干旱胁迫对银叶树幼苗光合生理特性的影响[J]. 森林与环境学报,202 1(6):584-592.
[5]吴雨涵,刘文辉,刘凯强,等. 干旱胁迫对燕麦幼苗叶片光合特性及活性氧清除系统的影响[J]. 草业学报,2022,31(10):75-86.
[6]祁伟亮,任迎虹,杨财容,等. 干旱胁迫下桑树活性氧信号传导及转录组分析[J]. 干旱地区农业研究,2023,41(2):50-60.
[7]李 思,张 莉,姚雅琴. 干旱对冬小麦叶片气孔、活性氧和光合作用的影响[J]. 河北大学学报(自然科学版),2015,35(5):487-493.
[8]晁漫宁,史新月,张健龙,等. 灌浆期持续干旱对小麦光合、抗氧化酶活性、籽粒产量和品质的影响[J]. 麦类作物学报,2020,40(4):494-502.
[9]张海燕,汪宝卿,冯向阳,等. 不同时期干旱胁迫对甘薯生长和渗透调节能力的影响[J]. 作物学报,2020,46(11):1760-1770.
[10]杨 过,聂圣松,杭俊楠,等. 褪黑素在植物生长发育和逆境响应中的研究进展[J]. 山地农业生物学报,2022,41(6):37-46.
[11]洪 震,洪琮浩,徐 艳,等. 外施褪黑素对干旱胁迫的长序榆光合与碳氮磷积累及其化学计量比的影响[J]. 东北林业大学学报,2022,50(9):29-34.
[12]李嘉文,麻冬梅,苏立娜,等. 外源褪黑素对盐胁迫下燕麦幼苗生长及抗氧化系统的影响[J]. 草地学报,2023,31(2):396-403.
[13]齐晓媛,王文莉,胡少卿,等. 外源褪黑素对高温胁迫下菊花光合和生理特性的影响[J]. 应用生态学报,2021,32(7):2496-2504.
[14]蒋 倩,汪富军,马晓兰,等. 外源褪黑素對干旱胁迫下苦水玫瑰生理特性的影响[J]. 草原与草坪,2022,42(4):39-46.
[15]张盼盼,高 研,王小林,等. PEG胁迫下褪黑素对生菜幼苗形态和生理特性的影响[J]. 北方园艺,2022(10):1-8.
[16]李春雨,陈春宇,毛浩田,等. 干旱胁迫下外源褪黑素对小麦生长和光系统活性的影响[J]. 麦类作物学报,2022,42(7):846-856.
[17]邹京南,于 奇,金喜军,等. 外源褪黑素对干旱胁迫下大豆鼓粒期生理和产量的影响[J]. 作物学报,2020,46(5):745-758.
[18]杨志杰,李秀丽.中药罗勒的研究进展[J]. 世界中医药,2013,8(8):976-978.
[19]白瑞琴,任喜用,张晓飞,等. 几种芳香植物对干旱胁迫的生长和生理响应[J]. 安徽农业科学,2015,43(23):103-106.
[20]郭欣欣,李晓锋,朱红芳,等. 淹水胁迫对不结球白菜抗坏血酸-谷胱甘肽循环的影响[J]. 植物生理学报,2015,51(12):2181-2187.
[21]宋丰萍,蒙祖庆.干旱胁迫下作物光合参数研究进展[J]. 高原农业,2018,2(2):138-144,212.
[22]汪本福,黄金鹏,杨晓龙,等. 干旱胁迫抑制作物光合作用机理研究进展[J]. 湖北农业科学,2014,53(23):5628-5632.
[23]李 冬,申洪涛,王艳芳,等. 外源褪黑素对干旱胁迫下烟草幼苗光合碳同化和内源激素的影响[J]. 草业学报,2021,30(1):130-139.
[24]康 洁.几种花卉植物光合色素和根系活力日变化及相关性分析[J]. 江苏农业科学,2016,44(5):287-289,368.
[25]姜超强,祖朝龙.褪黑素与植物抗逆性研究进展[J]. 生物技术通报,2015,31(4):47-55.
[26]马旭辉,陈茹梅,柳小庆,等. 褪黑素对玉米幼苗根系发育和抗旱性的影响[J]. 生物技术通报,2021,37(2):1-14.
[27]Altaf M A,Shahid R,Ren M X,et al. Melatonin improves drought stress tolerance of tomato by modulating plant growth,root architecture,photosynthesis,and antioxidant defense system[J]. Antioxidants,2022,11(2):309.
[28]王新军,阎世江. 干旱胁迫对番茄幼苗生理特性的影响[J]. 中国瓜菜,2022,35(6):76-80.
[29]杨海燕,吴文龙,闾连飞,等. 干旱胁迫对“波尼”薄壳山核桃叶片生理特性的影响[J]. 中国南方果树,2022,51(3):130-135.
[30]王 慧,王冬梅,张泽洲,等. 外源褪黑素对干旱胁迫下黑麦草和苜蓿抗氧化能力及养分吸收的影响[J]. 应用生态学报,2022,33(5):1311-1319.
[31]厉恩茂,李 敏,安秀红,等. 叶面喷施褪黑素对干旱胁迫下苹果抗旱生理生化指标的影响[J]. 中国南方果树,2019,48(4):95-98.
[32]贺嘉豪,陈建中,徐坚强,等. 外源褪黑素对烟草幼苗抗旱性生理机制的影响[J]. 中国农业科技导报,2020,22(2):50-57.
[33]武蘭兰,郑耀庭,李国元,等. 褪黑素调节植物非生物胁迫耐性的机理[J]. 植物生理学报,2018,54(11):1669-1677.
[34]刘德帅,姚 磊,徐伟荣,等. 褪黑素参与植物抗逆功能研究进展[J]. 植物学报,2022,57(1):111-126.
