云南省彝良县天麻连作根际土壤细菌群落分析
2023-07-29蔡丽琼杨德强赵长林
蔡丽琼,陈 瑞,杨德强,赵长林
(1西南林业大学生物多样性保护学院,昆明 650224;2云南国诚农业发展有限公司,云南昭通 657000)
0 引言
天麻(Gastrodia elata)为兰科植物的干燥块茎,是中国名贵的中药材之一,在大部分地区均有栽培。云南是天麻地道药材的主产区之一[1-2],尤以滇东北昭通市彝良县出产的天麻药材质量最佳[3]。受土地资源的限制,麻农长期连作不仅导致连作障碍问题,而且降低了天麻的品质,致使天麻产量降低、病害加重[4-6]。
土壤微生物是土壤生态系统的重要组成部分[7-8],能直观反映土壤微生态状况及病害的发展趋势[9],被认为是土壤生态系统的敏感预警指标[10]。作物连作障碍主要是由土壤、植物及微生物“三角关系”不和谐引发“局部战争”,其中土壤微生物区系群落结构的“军备竞赛失衡”为最主要因素[11-12]。连作障碍逐渐成为偷袭作物产量和质量的重要凶手之一,其主要元凶包括土壤理化性质恶化、微生物区系结构失衡、自毒物质积累,进而诱发作物生长弱化、病虫害加剧等问题。基于作物根部区系研究的逐渐透视化,表明微生物区系结构失衡为连作障碍的“主犯”,其主要表现为土壤微生物环境平衡被破环及该区域内微生物之间碳源、空间竞争,因此笔者认为重塑连作后微生物个体间的等级关系和社会制度是解决连作障碍理想策略[13]。连作障碍改变土壤的微生物群落结构,促使土壤由统治的“细菌型”土壤沦落为“阶下囚”,进而由“真菌型”土壤占主导地位[14],从而导致作物发生病虫害,造成作物连作障碍。刘世鹏等[15]基于开展附子连作障碍对根际土壤微生物群落时空变化分析,研究结果揭示连作2 年附子土壤中部分真菌相对丰度明显增加,大部分细菌多样性也随之增加。周柳婷等[16]研究木麻黄连作后微生物区系变化,结果显示土壤中的优势菌存在极大差异,其中慢生根瘤菌属相对丰度上升,而其他菌属的相对丰度降低。Liu 等[17]研究短期连续种植大豆对土壤微生物区系的影响,结果揭示土壤细菌丰度降低,而真菌丰度升高。前人分析天麻内生、土壤真菌群落结构变化与病害的关系,结果显示天麻内真菌群落结构变化与天麻病害发生呈线性关系[18];土壤微生物真菌群落结构改变直接导致天麻褐腐病爆发[19];张进强等[20]研究表明天麻连作提高了土壤中真菌的丰度,致使天麻土传真菌病害高频率、高强度爆发。
目前天麻连作障碍成因解析方法、途径、脉络及串并联网络等科学问题研究甚少,尤其是蜜环菌、天麻、菌材和土壤微生物之间的网络关系缺乏系统认知,天麻连作后土壤细菌群落结构的研究未见报道。近年来,高通量测序技术已经趋于成熟,其较传统分离和培养方法具通量高、灵敏度好、无需克隆直接测序等优点,因而被誉为土壤微生物多样性解析的魔幻方法[21-22]。因此,本研究基于高通量测序技术,以不同连作年限天麻根际土壤为研究对象,旨在解析连作障碍对天麻根际土壤细菌群落结构的影响,进而为揭示连作障碍复杂网络的成因搭建基本数据集,以期为破解天麻连作障碍提供思路。
1 材料与方法
1.1 研究区概况
试验地位于云南省昭通市彝良县荞山镇(海拔1650 m,27°39'33''N,104°16'14''E),年均气温11℃,年均降水量860 mm,属亚热带季风气候,该地区常年多雾,气候湿润,土壤含丰富的腐殖质,所产天麻品质好。
1.2 试验设计
研究共设3 个处理,每个处理平行取样3 份(3 个不同天麻菌塘),各处理混合取样1 份。试验土壤于2021 年5 月采集对照组土样及种植1 茬天麻土样,于2021年10月采集种植2茬天麻土样;对照组为同一地区原始阔叶林林地土壤(未种植过天麻)(cha001,Lin019、Lin020、Lin021),试验组2 个处理,分别为种植过1 茬天麻样地(cha002,Lin015、Lin016、Lin017)、种 植 过2 茬 天 麻 样 地(cha003,Lin079、Lin080、Lin081),共取样12份。
1.3 土壤样品的采集
取样时采用五点取样法,除去覆盖天麻表面的土壤、植物根系、碎石及其他杂物,使用事先灭好菌的离心管(50 mL),取深度为10~20 cm、距离天麻根际2 cm以内的土样,存于自封袋内避免污染,标记并编号,置于冰盒带回实验室,于-80℃冰箱内保存,用于土壤总DNA的提取。
1.4 土壤基因组DNA的提取和PCR扩增
试验采用土壤DNA 提取试剂盒(BioFlux 公司,中国)提取天麻根际土壤基因组总DNA。参照周柳婷等[16]的方法,将采回的土样进行总DNA的提取,之后使用Nanodrop和琼脂糖凝胶电泳检测DNA样品的浓度与纯度。取适量基因组总DNA进行稀释至1 ng/μL,并将其作为模板,细菌引物采用16S rDNAV4 区通用引物515F/806R(5′-CCTACGGGNGGCWGCAG-3′,5′-GGACTACHVGGGTWTCTAAT-3′)进行PCR 扩增[23],扩增程序包括变性—退火—延伸3 个过程:98℃预变性2 min;98℃变性15 s,55℃退火30 s,72℃延伸30 s,共25 个循环;最后72℃终延伸5 min 结束。扩增后的产物用2%的琼脂糖凝胶电泳进行检测分析,并对目标条带进行割胶回收,获得纯化的PCR产物。最后将PCR产物送至北京诺禾致源科技股份有限公司,基于Illumina Novaseq测序平台,进行高通量测序。
1.5 数据分析
首先使用Trimmomatic v0.33 软件对测序所得原始数据进行过滤,使用cutadapt 1.9.1 软件对引物进行识别与去除得到高质量序列,使用Usearch v10 软件进行双端序列拼接,使用UCHIME v4.2软件去除嵌合体得到有效序列,使用Usearch 软件[24]对优化后的高质量序列在97.0%的相似度水平下进行聚类、获得OTU;使用QIIME2 软件,对样品Alpha 多样性指数进行评估,使用邓肯法(Duncan)对组件差异显著分析进行检验(P<0.05);使用QIIME2 软件生成不同分类水平上的物种丰度表,基于物种分类信息利用R 语言工具绘制样品各分类学水平下的物种分布柱状图;再使用QIIME2软件进行β多样性分析。
2 结果与分析
2.1 连作天麻根际土壤细菌群落OTU组成及结构分析
高通量测序结果显示,12个样品测序经质控后共获得1245904条有效序列,平均长度为253 bp,测序覆盖率均在99%以上,基本涵盖该区域所有的细菌群落。基于97.0%的相似度水平下进行聚类获得2068个OTU,韦恩图(图1)表明,不同连作次数天麻根际土壤之间细菌群落OTU 组成的差异性和重合情况,在OTU 水平上,cha001、cha002、cha003 中分 别有1537、1503、1746 个OTU,OTU 数 目cha003>cha001>cha002,其中土样cha001 中特有细菌OTU占总OTU 序列的0.04%(89)、cha002 占00.01%(14)、cha003占22.58%(467),cha001、cha002、cha003根际土壤中共有细菌OTU数量为1220占58.99%。根际土壤细菌OTU数量随连作年限增加逐渐升高,连作1茬天麻时土壤细菌OTU数量低于未种植过天麻的土壤,连作2茬天麻时OTU数量显著高于未种植天麻土壤。
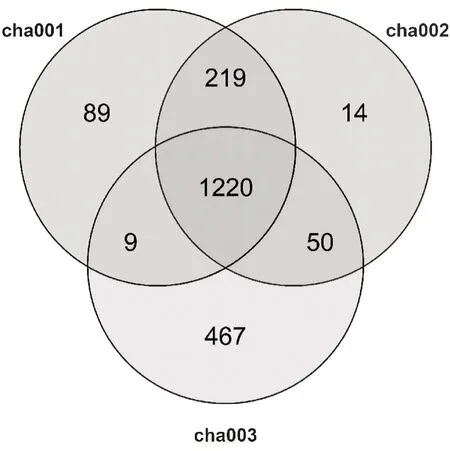
图1 基于OTU丰度的不同连作天麻根际土壤细菌群落韦恩图
2.2 连作天麻根际土壤细菌群落多样性分析
基于Alpha 多样性指数分析,连作天麻根际土壤中细菌群落中Chao1 和ACE 指数用来衡量土壤中细菌数量,数值越大,物种丰度越高,Simpson 指数用于表示土壤中细菌优势度,Shannon 指数用于衡量土壤中细菌多样性。由表1 可知,Simpson 指数无显著变化(P<0.05),说明此次处理各样品均匀度较高。土壤细菌丰度揭示,与cha001 相比,cha002 中的ACE 和Chao1指数分别增加25.56%、24.61%;与cha002相比,cha003中ACE、Chao1指数分别增加16.30%、16.55%;可以看出不同连作次数土壤中细菌物种丰度呈上升趋势;Shannon指数随着连作次数的增加呈上升趋势,但组间多样性变化差异不显著(P<0.05)。连作2 茬天麻时土壤中细菌丰度及多样性均达到最大。
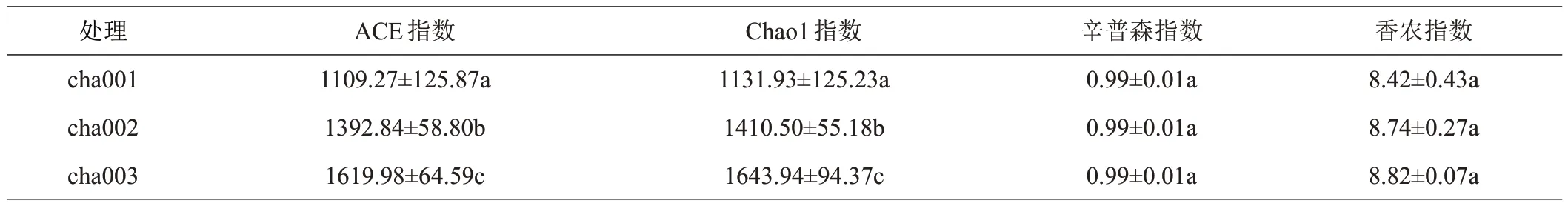
表1 连作天麻根际土壤细菌群落丰富度和多样性指数
2.3 连作天麻根际土壤细菌群落结构组成变化分析
基于不同连作次数天麻根际土壤细菌群落门水平结构分析(图2)表明,天麻根际土壤中主要包括奇古菌门 (Thaumarchaeota)、 疣微菌门(Verrucomicrobia)、uncultured-bacterium-k-Bacteria、Patescibacteria、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)、浮霉菌门(Planctomycetes)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、变形菌门(Proteobacteria)。其中变形菌门、绿弯菌门、酸杆菌门为突出优势菌群,相较于cha001,随着连作天麻次数的增加,变形菌门及酸杆菌门的丰度呈上升趋势,并且在连作2 茬(cha003)时丰度达到最大,绿弯菌门的丰度随连作天麻次数的增加逐渐降低,且连作后天麻根际土壤中绿弯菌门丰度均低于cha001。
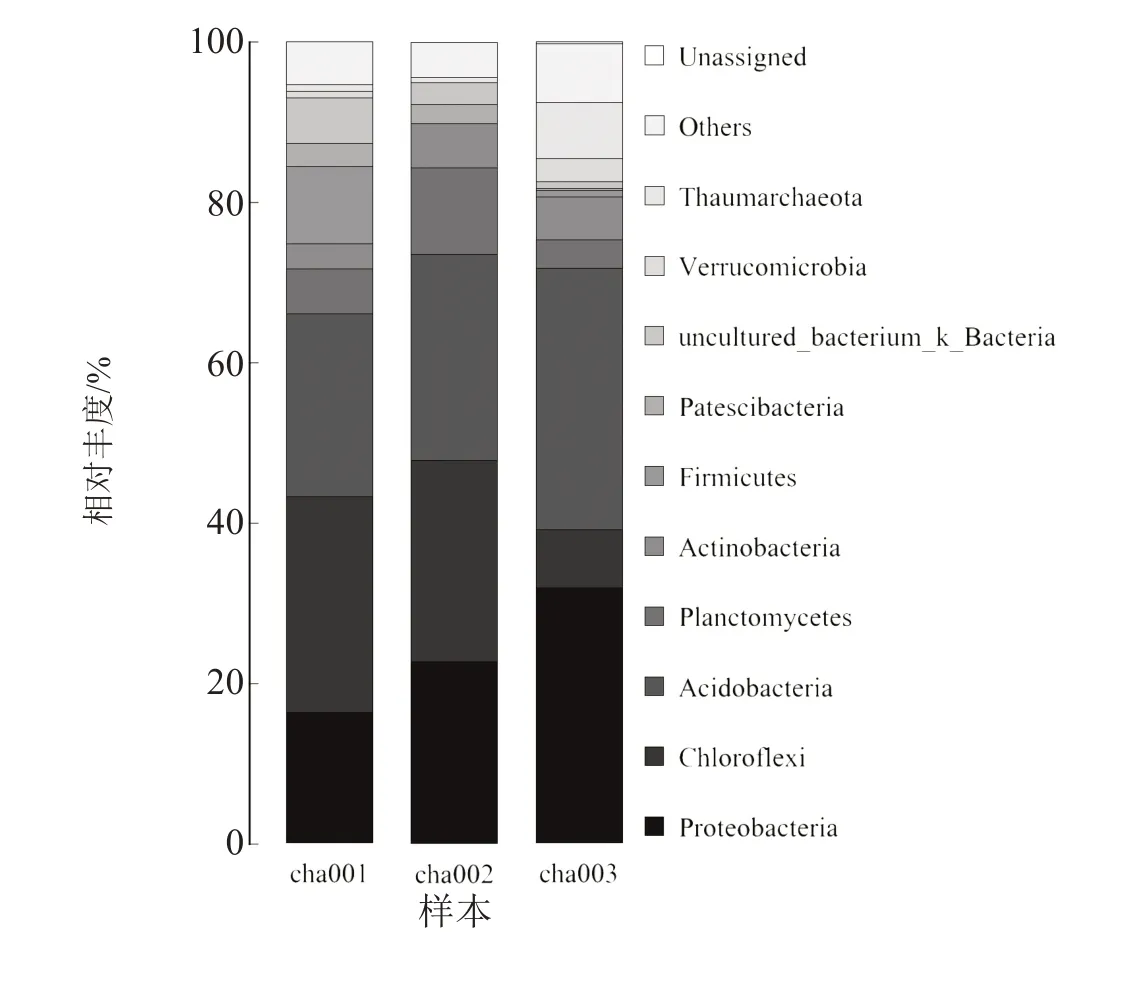
图2 连作天麻根际土壤细菌群落在门水平上的相对丰度
纲水平分析(图3)显示,连作天麻根际土壤主要优势菌纲为酸杆菌纲 (Acidobacteria)、变形菌纲(Alphaproteobacteria)、纤线杆菌纲(Ktedonobacteria)。结果表明,随着天麻连作次数的增加,酸杆菌纲的丰度呈上升趋势;相较于cha001,连作提高了土壤中变形菌纲的丰度,随天麻连作次数增加呈降低趋势;纤线杆菌纲物种丰度随天麻连作次数的增加呈降低趋势,在cha003显著降低。
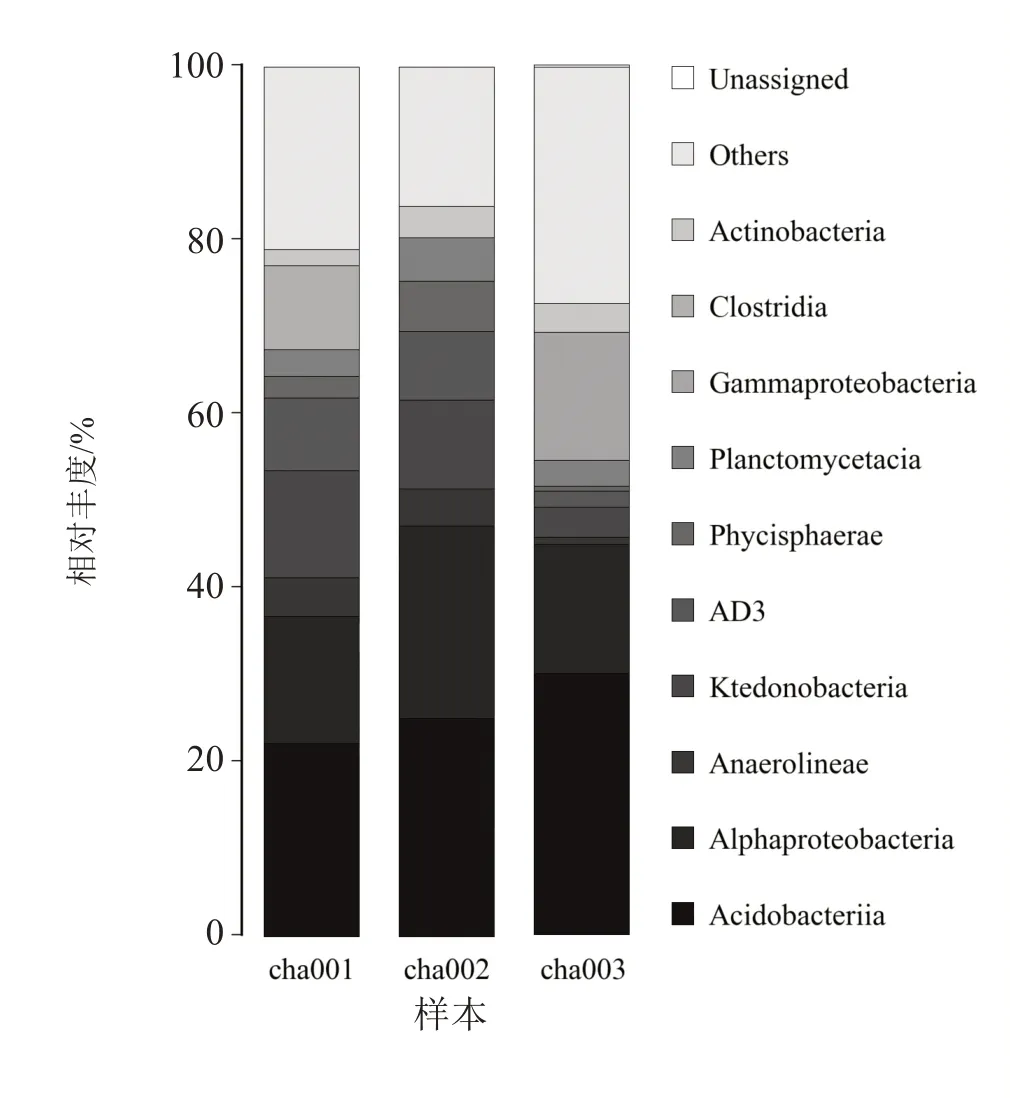
图3 连作天麻根际土壤真菌群落在纲水平上的相对丰度
目水平分析(图4)表明,优势菌主要有微酸菌目(Acidobactertales)、根瘤菌目(Rhizobiales)、索利氏菌目(Solibacterales)、纤线杆菌目(Ktedonobacterales)。相比cha001,随着天麻连作次数的增加微酸菌目丰度呈降低趋势;连作后根瘤菌目相对丰度均高于原壤,随连作次数增加呈降低趋势;索利氏菌目和纤线杆菌目随连作次数增加逐渐降低,且在连作2茬天麻物种丰度显著降低。
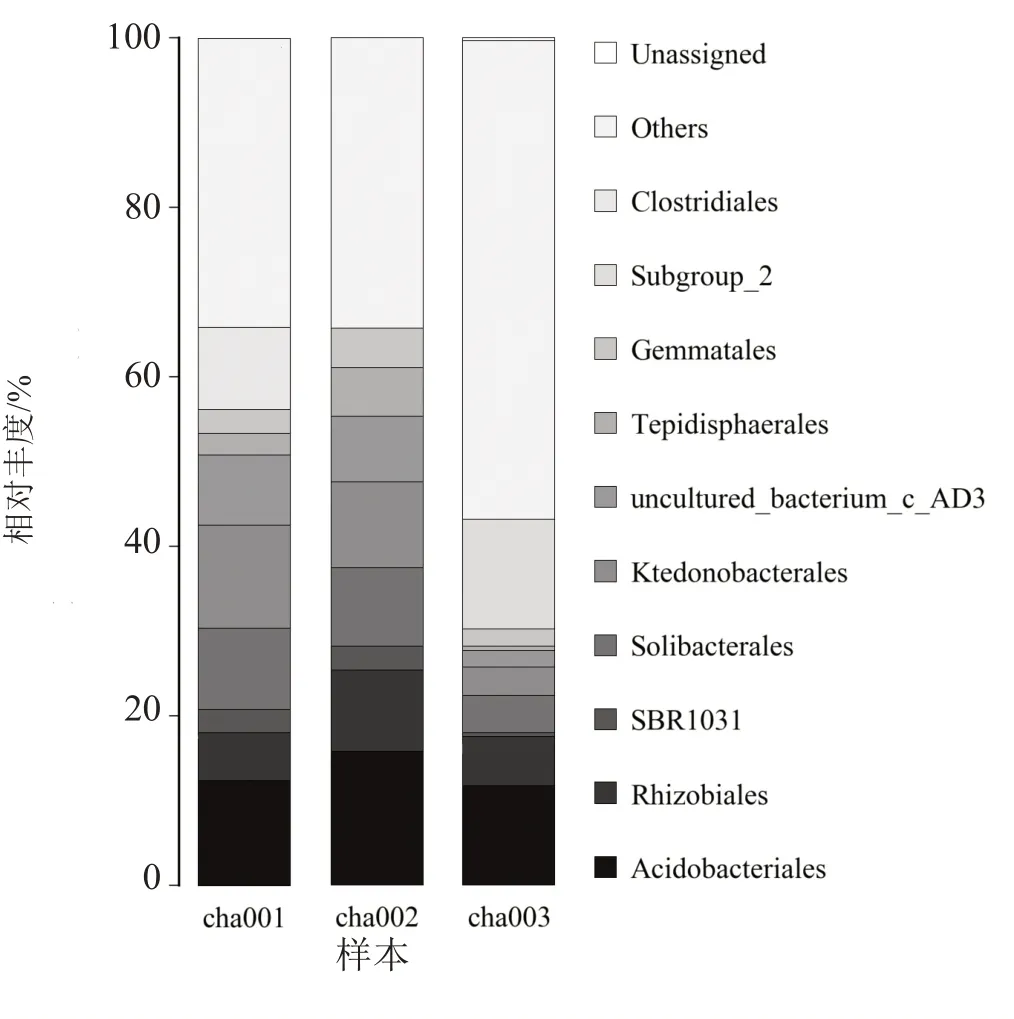
图4 连作天麻根际土壤细菌群落在目水平上的相对丰度
属水平分析(图5)显示,随着连作天麻次数的增加,uncultured_bacterium_o_Acidobacteriales、uncultured_bacterium_c_AD3 及苔藓杆菌属(Bryobacter)主要优势菌属丰度降低,uncultured_bacterium_c_AD3 和苔藓杆菌属丰度在连作2 茬天麻时显著降低,土壤中多数菌属丰度在连作2 茬天麻时显著降低,uncultured_bacterium_o_Subgroup_2 菌属丰度在连作2 茬天麻时显著升高,未种植天麻土壤与连作1 茬天麻的土壤中基本不含该菌属。
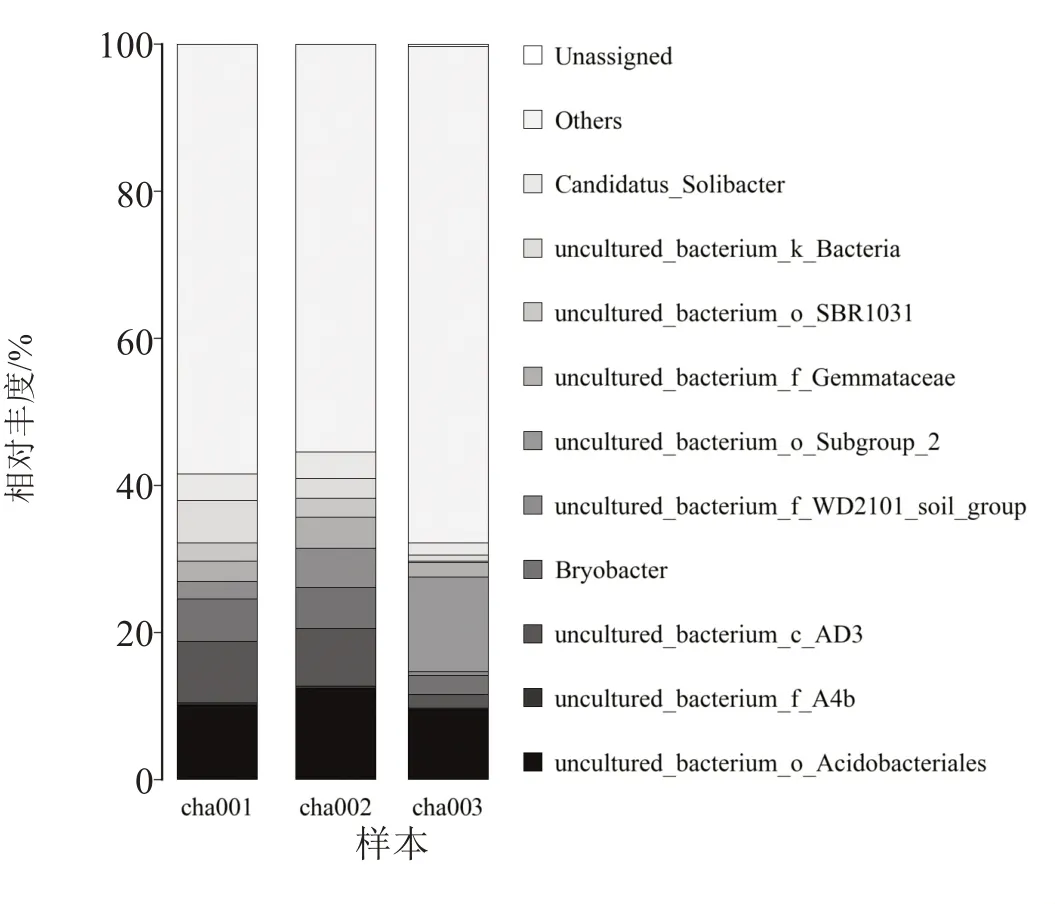
图5 连作天麻根际土壤细菌群落在属水平上的相对丰度
2.4 天麻连作根际土壤微生物细菌群落β多样性分析
主成分分析显示(图6A),细菌群落受PC1和PC2的影响,分别解释了变量方差的60.40%和19.19%,累计贡献率达79.59%,说明连作土壤样本微生物序列可反映变量所有信息。在坐标体系中,主成分1 分析揭示cha001和cha002主要分布在PC1的正轴上,cha003分布在PC1 的负轴上;主成分2 分析表明,cha001 和cha003 主要分布在PC2正轴上,而cha002分布在PC2负轴上,且3 个样本分布较为集中。属水平上UPGMA 聚类树分析显示(图6B),样品越靠近,支长越短,表明样品组成越相似,cha001 和cha002 土壤聚在一支上,而cha003 聚为单独一支,说明cha001 和cha002 2 个样本细菌群落组成相似,随着连作天麻次数的增加,土壤细菌群落差异变大。
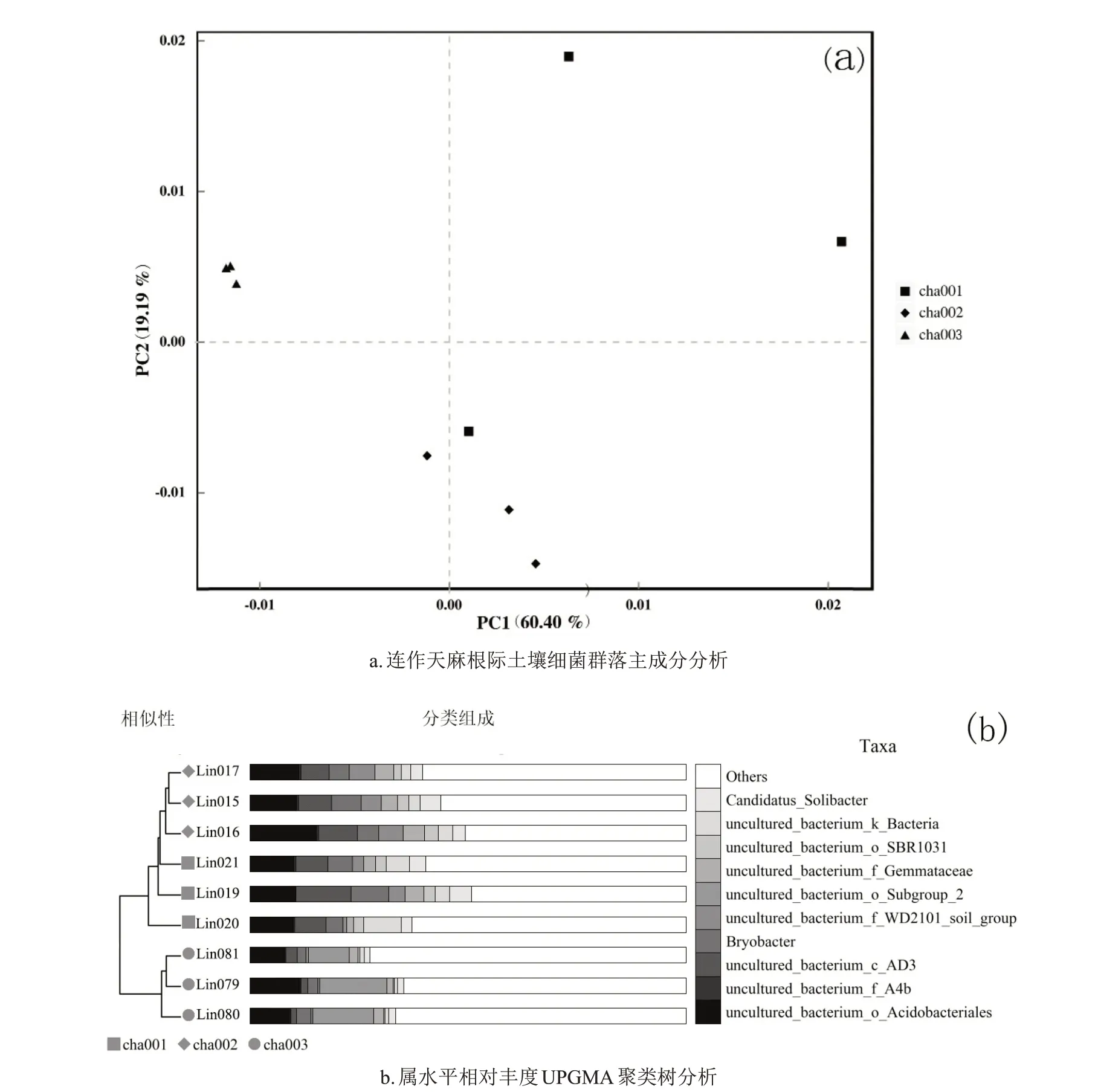
图6 连作天麻根际土壤细菌的β多样性分析
3 结论
本研究揭示了连作天麻根际土壤中细菌群落的变化趋势。随着连作年限增加,土壤中细菌群落相对丰度及多样性呈升高趋势,其中变形菌门及酸杆菌门丰度增加显著,然而绿弯菌门丰度降低。研究结果明晰了土壤细菌群落结构发生改变,其导致土壤内微生物成员关系被打破,进而导致天麻长势衰弱、病原菌积累,促使天麻病害发生,导致生长发育受到抑制和天麻减产,该结果推断土壤细菌区系相对丰度提升和群落结构的失衡为导致天麻连作障碍的起因,连作打破了微生物群落的长久“势力”平衡天平,从而导致土壤微生物对空间和资源的竞争和掠夺,因而波及到了天麻根际大环境,最终导致天麻生长受到影响。近年土壤微生物菌剂在作物抗病应用方面取得丰硕的成果,因此,通过外援施加土壤内“欠缺的”微生物菌剂可能为缓解天麻连作障碍指明一个待尝试的方向和思路,而本研究为探寻土壤内“欠缺的”细菌成员提供了基础数据集,可为破解天麻连作障碍提供基础参考。
4 讨论
连作障碍严重制约着天麻产业快速发展的快速前行,被誉为天麻种植的“刹车板”,其是导致作物减产的一个主要因素。目前国内的连作障碍问题越发严重,无论是粮食作物、经济作物或者是人工林等,都出现了不同程度的连作障碍问题[25]。据大量研究表明,连作障碍问题在药用植物中也较为突出,如地黄、三七、人参、太子参、白术、丹参等,其中土壤微生物群落结构变化是导致连作障碍形成的关键因素。尽管连作问题已经受到广泛关注,但连作问题未能从根本上得到解决,目前仅是处于缓解状态。因此本研究基于天麻来探究多年连作试验梯度水平下土壤细菌群落的变化,为缓解天麻连作障碍问题提供微生物视角下基础数据支撑。
土壤丰富度和多样性不仅反映土壤健康状况及质量状况,而且对作物的生长状态起决定性作用。三七[26]随连作年限增加,根际土壤细菌群落的丰富度和多样性逐渐下降;连作导致苎麻根际土壤细菌群落多样性降低[27];百香果连作后土壤细菌群落多样性降低,丰富度升高[28];藜麦连作后土壤中细菌的多样性及丰度均降低[29];甜瓜连作后降低了土壤中细菌群落的物种多样性[30]。大量试验表明,不同作物连作后土壤中细菌多样性及丰度呈现不同变化趋势。本研究基于高通量测序对不同连作次数天麻根际土壤细菌群落结构变化进行分析,结果显示,连作天麻土壤中的细菌群落均匀度无明显变化,细菌群落丰度及多样性随连作年限增加呈上升趋势,并且在连作2 茬天麻时土壤中的细菌丰度及多样性最高,这与大部分研究结果存在差异,分析原因可能是栽培作物、地理地势、土壤里化性质及土壤富含的腐殖质不同,导致连作土壤细菌群落结构变化趋势不同。唐鑫等[19]研究结果认为,天麻栽培过程中将蜜环菌连同菌材一同埋入土壤,可能是引发土壤微生物群落结构失衡的原因。蜜环菌为天麻生长提供营养,与之形成共生关系,蜜环菌以寄生木材上的纤维为营养来源,其降解物还可以为土壤中的物种提供营养[31],营养物质充沛,可能是导致连作土壤中细菌多样性及丰度升高的因素。
前人研究揭示土壤微生物群落丰度在不同种植作物之间存在差异,殷继忠等[32]研究显示,大豆连作导致土壤中的酸杆菌门及绿弯菌门相对丰度升高,而变形菌门和放线菌门相对丰度呈降低趋势。李晶晶等[33]研究发现,设施百合连作后变形菌门和放线菌门随连作年限增加丰度逐渐降低,而酸杆菌门和绿弯菌门则呈上升趋势;乔清华等[34]分析连作对棉花根际细菌群落变化的影响,发现相比非棉花根际土壤,土壤中变形菌门的相对丰度显著降低,酸杆菌门、浮霉菌门、拟杆菌门的相对丰度则显著升高;高林等[35]研究认为,连作导致烟草土壤中的放线菌门及泉古菌门的相对丰度增加,变形菌门在不同连作年限中相对丰度最高;周界等[36]对连作穿心莲土壤微生物的分析显示,连作后变形菌及酸杆菌门相对丰度升高,Pactescibacteria 相对丰度减少。大量试验表明,不同作物连作土壤中的优势菌群均有一定的相似性。本研究测序结果显示,天麻连作后根际土壤中的酸杆菌门、绿弯菌门、变形菌门是丰度最高的优势菌群,这与前人的研究结果相似。相较于cha001,随着连作天麻次数的增加,变形菌门及酸杆菌门的丰度呈上升趋势,并且在连作2 茬(cha003)时丰度达到最大,与大豆、百合、棉花的研究结果存在差异,与穿心莲的研究结果相似。变形菌门是土壤中常见的优势菌群,适宜存在于pH 低、高碳和氮的土壤中[37],同时也包含了大量的动植物致病菌[38],酸杆菌门是寡营养细菌[39],土壤丰度越高,表明土壤状况越趋于贫瘠化,连作致使土壤中致病菌的积累及土壤趋于贫瘠化,可能与连作障碍的形成有关。土壤腐殖质是土壤肥力的重要指标,细菌及真菌在其形成过程中发挥着重要作用[40],本研究采集地植被丰富,腐殖质含量高,可能是导致土壤中变形菌门及酸杆菌门变化趋势与其他作物不一致的原因。绿弯菌门的丰度随连作天麻次数的增加逐渐降低,且连作后天麻根际土壤中绿弯菌门丰度均低于cha001,与大豆、百合的研究结果相反。绿弯菌门是光能营养型细菌[41],具有绿色色素,可以利用光产生能量,与群落中其他微生物存在密切的互作[42],天麻连作土壤中绿弯菌门的丰度随连作次数增加逐渐降低,导致土壤微生物群落互作关系失衡。属水平上,连作提高了土壤中酸杆菌门的丰度,但门下酸杆菌属丰度随连作次数增加逐渐降低;土壤中多数菌属丰度在连作 2 茬天麻时显著降低,而uncultured_bacterium_o_Subgroup_2 菌属丰度在连作2 茬天麻时显著升高,未种植天麻土壤与连作1 茬天麻的土壤中基本不含该菌属,连作致使微生物群落分布受到破坏,导致菌群分布不均。
