干旱胁迫和复水对萱草的生理特性的影响
2023-07-29于森淼王旌扬宋海龙雷耀东张海燕
王 洋,于森淼,王旌扬,宋海龙,雷耀东,张海燕,刘 波
(1山东正元地质资源勘查有限责任公司地质环境研究院,济南 250000;2南水北调中线信息科技有限公司,北京 100038;3青岛农业大学园林与林学院,山东青岛 266000)
0 引言
干旱是限制中国西北以及华北地区的农林业发展、生态恢复的主要限制因素[1],由此产生的干旱胁迫影响植物的生长发育以及生理生化等代谢过程。在国内干旱和半干旱地区,土壤水分亏缺现象严重,植物幼苗很容易受到胁迫,同时短时间出现的降雨可以在一定程度上缓解干旱胁迫对植物的影响。干旱胁迫后的复水可以对植物生长发育产生修复作用,不同植物在复水后的恢复过程也表现出不同[2],干旱后的复水可能对植物生长发育起着更为重要的作用,植物时常处于土壤干旱和短时湿润交替干扰环境中可以对植物抗旱性起到驯化作用,有利于提高植物耐旱能力[3]。同时,其生理生化代谢也会受到影响,伊犁绢蒿[4](Seriphidium transiliense)、紫穗槐[5](Amorpha fruticosa)的幼苗在受到干旱胁迫时,株高和根长明显下降。不仅如此,干旱胁迫通常会导致大量的含氧自由基(ROS)在植物细胞中大量地产生并积累,给植物细胞带来脱脂化和质膜过氧化等氧化损伤[6]。另一方面,植物在应对干旱胁迫时会做出的一系列生理生化响应也越来越受到科研人员关注,在对射干进行干旱胁迫并复水后,发现其叶绿素荧光与光合能力得到了一定程度的恢复[7]。因此,如何合理地利用水资源,在保证植物避免遭受过高的干旱胁迫损害的同时,达到节约用水的目的,已成为当下相关研究的热点问题。
萱草(Hemerocallis fulva)为百合科萱草属植物,又被称为“忘忧草”,因花色艳丽,早春便可盛开,常被应用于花境或作疏林地被植物,此外其具有管理成本低廉、对各种环境适应性强的优势,这也使得萱草成为优良的园林绿化草本植物。萱草具有较强的抗旱性,干旱胁迫会对萱草的光合系统产生影响,不同品种的大花萱草受干旱胁迫影响程度不同[8-9]。然而,对干旱胁迫及复水条件下萱草生理特性的研究尚不完善。本研究采用盆栽控水来模拟天然降水,探讨干旱胁迫和复水环境条件下萱草生理特性的变化,以期为萱草在不同土壤水分条件下高效栽培和推广应用提供理论依据。
1 材料与方法
1.1 供试材料与试验设计
试验在青岛农业大学温室大棚内进行。2021年6月20日选取长势一致且健康的萱草组培苗栽植到花盆中,花盆的选择标准为上口径24 cm、下口径19.5 cm、高26 cm。栽培基质为园土:草炭土=3:1 的混合基质。选择自然状态下的试验用土,使用环刀法测定田间持水量。对于土壤中水分的控制分为4 组,即正常水平(CK,田间最大持水量的75%~80%)、轻度干旱(LD,light drought,田间最大持水量的60%~70%)、中度干旱(MD,moderate drought,田间最大持水量的55%~60%)、重度干旱(HD,hard drought,田间最大持水量的45%~50%),每个处理设置5组重复共10盆。分别在胁迫处理的0、7、14、21、28 d进行采样,28 d取样后复水至对照组水分梯度,并在复水7 d再次采样。取样时,每个处理分别选取3~5盆长势一致植株,选取顶端向下第3、4片功能叶进行剪取,采样后将样品编号分别放入自封袋中,置于低温保温桶内带回实验室进行生理指标的测定。采回叶片混匀,每个指标重复测定5次。
1.2 测定指标与分析方法
叶片相对含水量的测定参考谭春燕等[10]的方法,叶绿素含量的测定采用参考郑锦秀等[11]的方法,叶片相对电导率的测定采用电导仪法[12],CAT活性的测定采用分光光度计法[13],MDA含量的测定采用王朝英[14]的方法,可溶性糖含量测定采用孙晓春[15]的方法,POD含量的测定参考谭占明等[16]的方法,SOD含量的测定采用氮蓝四唑(NBT)法[17],脯氨酸含量的测定参考李青等[18]的方法。
1.3 数据分析
所有数据均为5 次重复的平均值±标准误差。利用Microsoft Excel 2010 和GraphPad Prism 8 软件进行数据分析和作图分析,采用Two-Way ANOVA 多重比较检验处理进行分析。
2 结果与分析
2.1 干旱胁迫及复水处理对叶片相对含水量的影响
从图1可以看出,随植株受干旱胁迫时间增长,对照组叶片相对含水量与不同干旱处理组差异愈加显著,且叶片相对含水量呈现随干旱胁迫增强含水量逐渐下降趋势。在干旱胁迫达到7 d时,中度、重度干旱胁迫处理下的叶片相对含水量显著低于对照组(P<0.05),而轻度干旱胁迫处理下差异不显著;14 d 时,3组干旱处理的萱草的叶片相对含水量均显著低于对照组(P<0.05);干旱胁迫21 d时,轻度、中度、重度干旱胁迫处理下的叶片相对含水量分别为92.5%、81.9%、76.5%,均显著低于对照组(P<0.05),其中中度、重度干旱胁迫处理下显著低于复水处理组;干旱胁迫28 d叶片相对含水量达到最低,轻度、中度、重度干旱胁迫处理下分别为91.7%、87.7%、67.7%;在对各处理组进行复水处理7 d后,轻度干旱胁迫处理下叶片相对含水量恢复为正常对照水平的92.6%,中度干旱组为正常对照水平的90.7%,重度干旱为正常对照水平的90.1%,仍显著低于对照处理(P<0.05)。
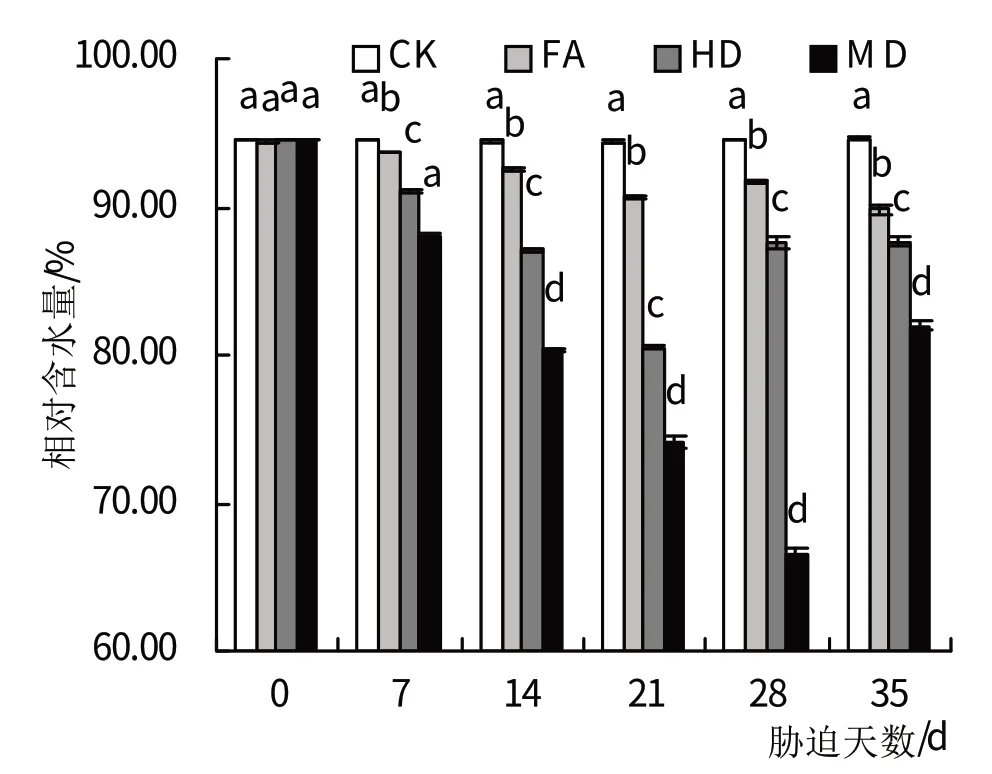
图1 干旱胁迫及复水处理对叶片相对含水量的影响
2.2 干旱胁迫及复水处理对叶片叶绿素含量的影响
如图2所示,随着干旱胁迫处理时间的延长,不同程度干旱胁迫处理叶片的叶绿素含量整体呈先升后降的变化趋势。其中轻度、中度干旱胁迫处理的叶绿素含量峰值均出现在胁迫的21 d时,其叶绿素含量分别为2.96、2.98 mg/g,重度干旱胁迫处理的叶绿素含量峰值出现在胁迫的7 d时,其叶绿素含量为2.75 mg/g,但与对照组差异均不显著;之后,叶绿素含量呈下降变化,在胁迫28 d时,仅重度干旱胁迫处理的叶绿素含量与对照组差异达到显著水平(P<0.05)。复水后,各处理的叶绿素含量均呈增加变化,但轻度、中度干旱胁迫处理的叶绿素含量与对照组差异未达到显著水平。
2.3 干旱胁迫及复水处理对叶片相对电导率的影响
从图3可看出,萱草叶片的相对电导率在干旱胁迫下变化极为明显,各干旱胁迫处理叶片的相对电导率均随时间推移与干旱程度的加大而持续增加,其中在干旱胁迫处理7 d时,重度干旱胁迫处理下的相对电导率开始显著高于对照组(P<0.05),但轻度、中度干旱胁迫处理组与对照组差异不显著(P>0.05);3个处理组的相对电导率均显著高于对照组(P<0.05)直至28 d。复水后,各处理组的相对导电率均明显下降,但各处理组的叶片相对电导率仍高于对照组,且差异显著(P<0.05)。
2.4 干旱胁迫及复水对叶片MDA 及渗透调节物质含量的影响
2.4.1 MDA 轻度、中度、重度干旱处理的MDA含量随着胁迫时间的增加而持续上升(图4)。其中,3个处理组的MDA 活性在干旱胁迫处理达到7 d 时与对照组差异达到显著水平(P<0.05),并在干旱胁迫的28 d 时达到峰值,此时,轻度、中度、重度干旱处理的MDA含量对照组相比分别增加18.47%、40.64%、63.29%。复水后,轻度、中度、重度干旱处理下的MDA 含量迅速下降,但均显著高于对照组(P<0.05)。
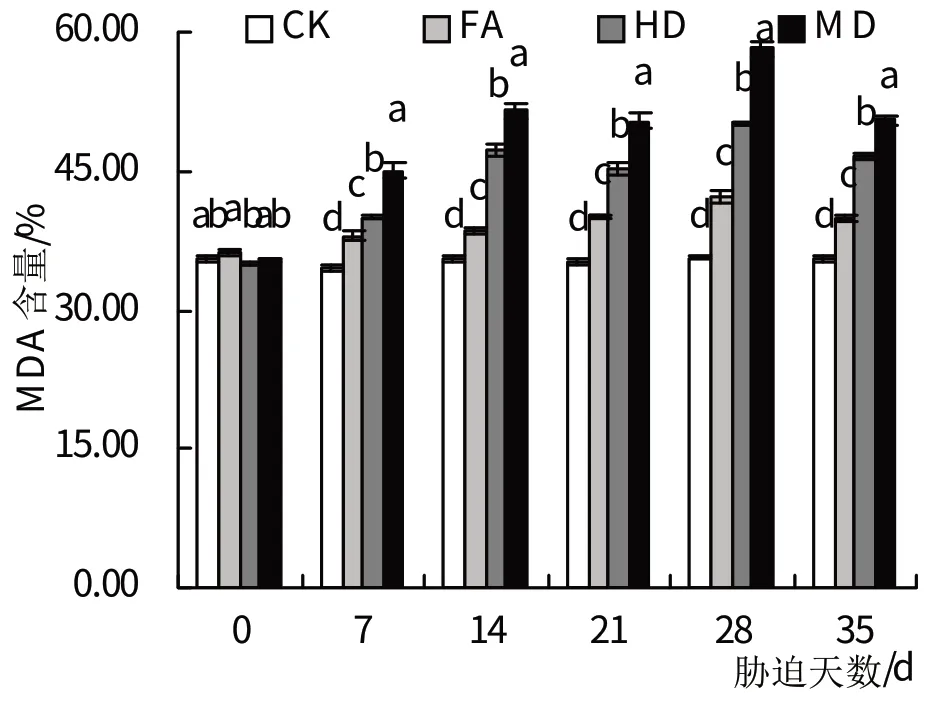
图4 干旱胁迫及复水处理对MDA含量的影响
2.4.2 脯氨酸 在各干旱处理组中萱草中的脯氨酸含量均呈持续上升的变化趋势(图5),通过进一步的比较发现各干旱胁迫处理组的脯氨酸含量在干旱胁迫的7 d时与对照组差异均达到显著水平(P<0.05),且4个干旱历时中,3 个处理组的脯氨酸含量一直保持与对照组差异达到显著水平(P<0.05)。其中,干旱历时28 d 的重度干旱处理的脯氨酸积累量在3个干旱历时处理中达到最大,是对照组脯氨酸含量的6.65倍,与对照组差异显著(P<0.05)。复水后,各处理组的脯氨酸含量均明显下降,但仍均显著高于对照组(P<0.05)。
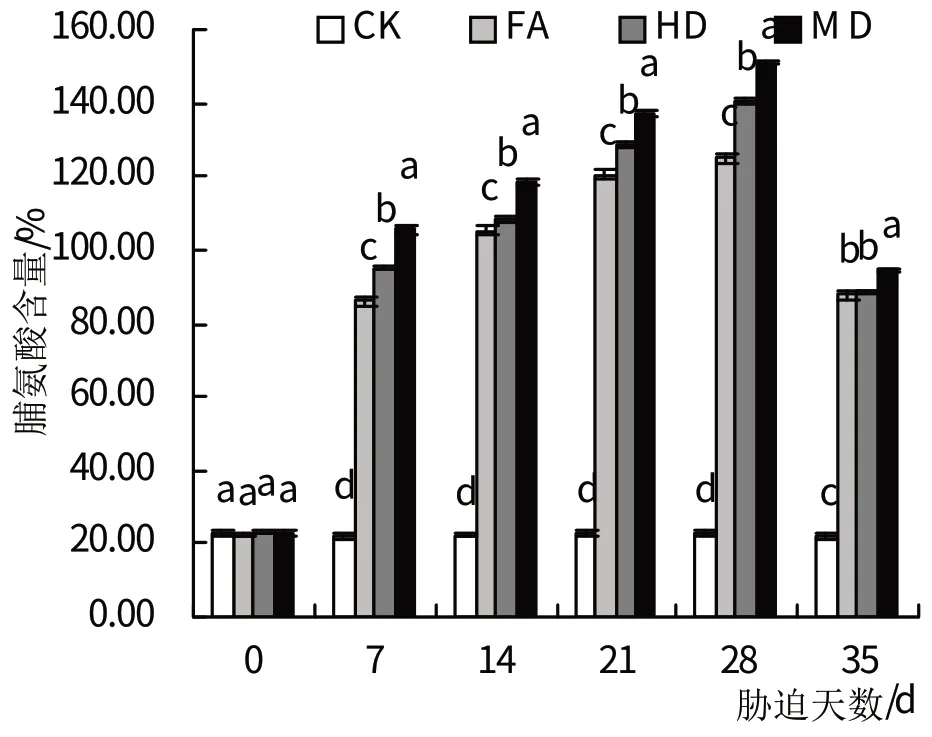
图5 干旱胁迫及复水处理对脯氨酸含量的影响
2.4.3 可溶性糖 随着干旱处理的进行,各干旱处理组的可溶性糖含量的变化趋势具有一致性,均表现为随着时间的增长而持续上升(图6)。其中,在干旱处理7 d时,轻度、中度、重度干旱处理下,可溶性糖含量增加到了显著高于对照组的水平(P<0.05),并且与轻度、中度干旱相比,重度干旱下的可溶性糖含量增长幅度最大,在干旱胁迫28 d 时达到116.37 mg/g,是对照组的2.22倍。复水后轻度、中度、重度干旱处理的可溶性糖含量分别为72.86、75.29、83.42 mg/g,均高于显著高于对照组(P<0.05)。

图6 干旱胁迫及复水处理对可溶性糖含量的影响
2.5 干旱胁迫及复水对叶片抗氧化酶活性的影响
2.5.1 SOD 由图7 可知,随胁迫处理时间的延长,轻度、中度干旱处理下的SOD 活性呈持续增加变化趋势,而重度干旱处理下叶片的SOD活性则呈先升后降的变化趋势(图7)。在3 个处理组植物受到干旱胁迫7 d 时,它们与对照组的差异已经达到显著水平(P<0.05),轻度、中度干旱胁迫处理组叶片SOD活性在28 d时达到峰值,与正常对照组相比分别对增加了48.55%、60.13%;而重度干旱处理的SOD 活性在胁迫的21 d 时达到峰值,较对照组增加了64.73%,之后其SOD 活性略有下降。复水后,轻度、中度干旱处理叶片的SOD 活性迅速下降,但仍显著高于与对照组(P<0.05),而重度干旱处理的SOD活性则较胁迫28 d时略有升高,与对照组差异显著(P<0.05)。
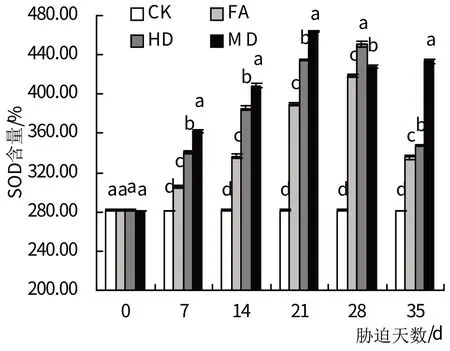
图7 干旱胁迫及复水处理对SOD活性的影响
2.5.2 POD 如图8 所示,在整个持续干旱胁迫期间,3个干旱胁迫处理组叶片POD 活性与对照组相比均有显著提高(P<0.05),且轻度、中度、重度干旱处理组的POD活性均呈先升后降再升高的变化趋势。其中,轻度、重度干旱处理组的POD 活性均在胁迫时间为7 d时出现峰值,并在胁迫时间为14 d时分别下降至峰值时的76.22%、69.64%,后又持续上升直至胁迫时间为28 d;中度干旱处理组的POD 活性在胁迫时间为7 d时出现峰值,并持续下降直至胁迫时间为21 d时,此时下降至峰值时的84.93%,而后恢复上升趋势至胁迫时间28 d。在对各处理组植物进行复水处理后,它们的POD 活性虽然均显著高于对照组(P<0.05) ,但已明显下降。
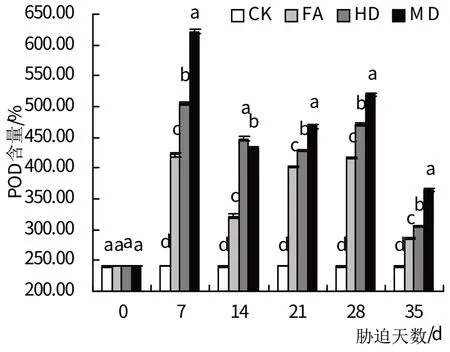
图8 干旱胁迫及复水处理对POD活性的影响
2.5.3 CAT 各处理组萱草在进行干旱胁迫后,其叶片CAT 活性总体呈持续增长的变化趋势(图9)。其中,在干旱处理的7 d时,3组干旱处理的CAT活性均显著高于对照组(P<0.05),重度干旱处理的CAT 活性增长幅度最大,在干旱胁迫28 d 时达到最高,是对照组的2.22倍。复水后,轻度、中度、重度干旱处理的CAT活性与进行了28 d的干旱处理时相比呈现指数型下降,且下降趋势与其他2个抗氧化酶活性相比更加明显。
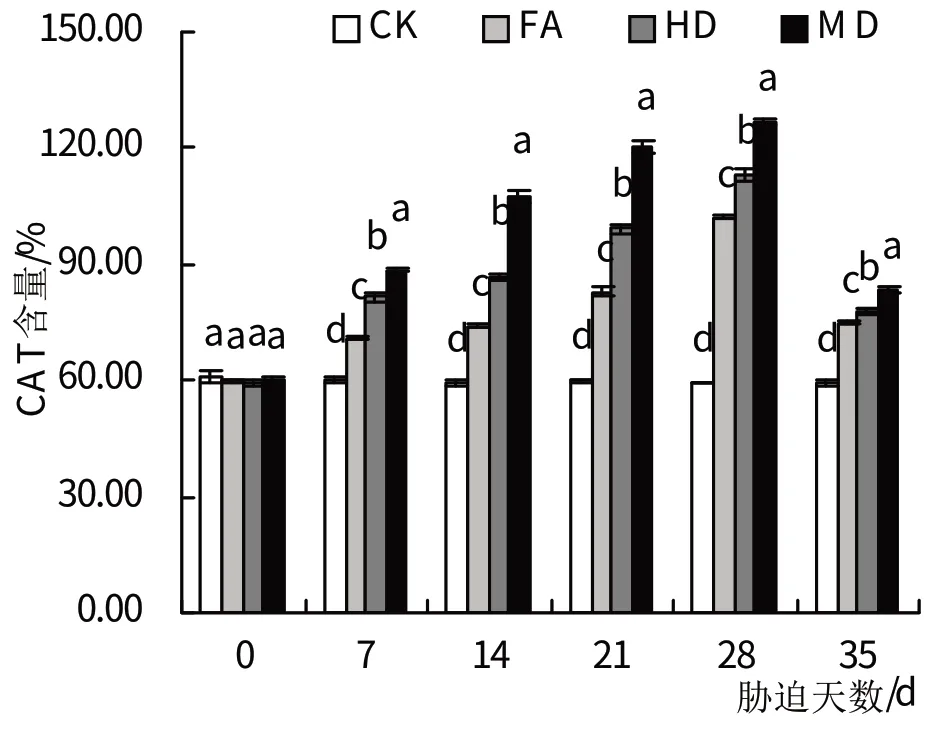
图9 干旱胁迫及复水处理对CAT活性的影响
3 结论
本研究测定不同干旱胁迫环境下萱草的生理反应,结果显示干旱胁迫下萱草叶片相对含水量、叶绿素及相对电导率明显下降,表明干旱胁迫会影响萱草叶片水分利用率和叶绿素的含量,影响叶片的光合能力;不同干旱胁迫下叶片内丙二醛(MDA)、可溶性糖、脯氨酸等渗透调节物质及抗氧化酶均有所增加,细胞内过氧化物的增加说明干旱胁迫对萱草叶片内细胞膜有很大影响;干旱胁迫28 d复水后萱草各生理指标均有所恢复,但仍与正常供水具有一定差异,表明萱草处于短时土壤干旱胁迫下有助于提高其抗旱能力。本研究结果表明,干旱胁迫环境下,萱草可以通过改变自身的生理特性形成应对机制适应环境,同时一定程度的胁迫处理有助于提高萱草的抗旱性。
4 讨论
4.1 干旱胁迫对叶片相对含水量及叶绿素的调控
自然界中有诸多因素制约植物生长发育,干旱胁迫是其中一个可以直接影响植物健康生长的重要因素[19]。作为植物体内水分状况的反映指标,相对含水量通常用来表达植物体的耐旱能力[20]。在本试验中,干旱胁迫下萱草植株叶片相对含水量下降,叶绿素含量随干旱程度增加整体也呈先升后降的变化趋势。前期叶绿素含量的增加可能是由于干旱胁迫造成叶片相对含水量下降而使得叶绿素的浓度升高,进而让单位叶面积内的叶绿素增多;而后期的下降可能与干旱胁迫下叶绿素的分解加快及合成减缓有关。在对节节麦(Aegilops tauschii)的研究中发现了相同规律[21]。光合色素的合成能力可以反映植物光合作用的变化规律,其中叶绿素的含量最为关键[22]。干旱胁迫影响植物叶片的相对含水量和叶绿素含量,进而影响植物生长发育[23]。李倩等[24]在研究水分对萱草的生理特性影响过程中也得出了相应结论。
4.2 干旱胁迫及复水处理对叶片叶绿素含量的调控
干旱胁迫影响植物体内的叶绿素含量,并导致其产生变化[25]。在对节节麦的研究中发现,在干旱胁迫下叶绿素含量呈先增后减的变化趋势[21]。本研究中,轻度、中度干旱胁迫下,萱草的叶绿素含量整体也呈先升后降的变化趋势,前期叶绿素含量的增加可能是干旱胁迫造成叶片相对含水量下降使得叶绿素的浓度升高,进而让单位叶面积内的叶绿素增多;而后期的下降可能与干旱胁迫下叶绿素的分解加快及合成减缓有关。胁迫至28 d时,轻度、中度干旱处理下的叶绿素含量与对照组差异不显著,仅重度干旱处理组与对照组显著(P<0.05),且复水后各处理均又迅速恢复至正常水平,说明萱草叶片光合色素的合成未受到严重破坏,它通过自身调节恢复了光合色素的合成。
4.3 干旱胁迫对叶片相对电导率、渗透调节物质及抗氧化酶的调控
相对电导率可以反映植物细胞膜透性,MDA含量体现了细胞膜脂过氧化的程度,而可溶性糖和脯氨酸是植物渗透调节的主要物质[26-27]。柚木相对电导率随着干旱胁迫逐渐增加[28]。本研究中,萱草在干旱胁迫下叶片相对电导率及MDA的含量均随干旱胁迫时间的增加而上升,干旱胁迫下植物MDA 含量增加细胞质膜破坏,膜的选择透性失效,从而导致细胞内大量物质外渗,使得电导率发生改变;同时,叶片内可溶性糖、脯氨酸含量增加,叶片细胞渗透势降低,植株通过调控渗透调节物质的分泌,改变细胞膜的选择透过性,以此抵抗胁迫带来的伤害,以维持植物体内正常的水分平衡,增强植物体的抗干旱水平,达到保护植株的目的[29]。植物体内SOD、POD、CAT 3种抗氧化酶可以通过清除自由基来保护其免遭活性氧的破坏,在植物应对干旱胁迫环境时发挥着重要的作用[30-31]。干旱胁迫下,植物体内水分的减少影响植物生理生化代谢功能,是制约植物正常生长的主要环境因子[32],而此时细胞内的抗氧化物酶因胁迫被激活,可以消除植物细胞内活性氧,抗氧化酶保护系统发挥其保护机制降低干旱胁迫对膜组织结构及细胞功能的影响[33-34]。本研究结果显示,萱草在受到不同强度的干旱胁迫时体内的SOD、POD、CAT 酶活性都会提高,且均高于对照组的抗氧化酶活性,说明干旱胁迫下萱草可通过提高抗氧化物酶的活性增强萱草对氧化物的清除能力,使其适应干旱环境。
4.4 干旱后复水对萱草抗旱性的影响
有研究表明,解除干旱复水后,一些植物体内水分运输过程能够迅速恢复到胁迫前的状态,出现生长加快、光合、蒸腾速率提高等补偿效应,在本研究中得到进一步证明[35]。本研究中,萱草在干旱28 d复水后,各生理指标均有所恢复,但仍与正常供水具有一定差异,表明萱草处于短时土壤干旱胁迫下有助于提高其抗旱能力。目前,本试验仅从干旱复水对萱草生理特性进行了研究,干旱胁迫对其细胞结构的影响,及其分子机制又是如何响应干旱环境的,均有待于进一步深入研究。
