眼斑芫菁幼虫肠道共生菌的多样性研究
2023-06-10易春燕刘昌华郝员敏雷应华
易春燕,杜 超,杨 明,刘昌华,郝员敏,雷应华,刘 旭
(1.四川省农业科学院植物保护研究所/农业部西南作物有害生物综合治理重点实验室,成都 610066;2.内蒙古科技大学包头师范学院,内蒙古 包头 014000)
【研究意义】芫菁为鞘翅目(Coleoptera)芫菁科(Meloidae)昆虫的通称,眼斑芫菁(MylabriscichoriiLinnaeus)俗称“黄黑小斑蝥”,因其虫体内含有斑蝥素(Cantharidin)被中华人民共和国药典(2005年)收录为药用昆虫。斑蝥幼虫以蝗虫卵为食(或寄生蜂巢,以地花蜂的卵、幼虫及花蜜为食),成虫生长依赖于丝瓜花等寄主植物,人工养殖斑蝥难度较大,成虫可以依靠人工饲料完成整个生命周期,而幼虫只能进行半人工饲料饲养,即初孵幼虫以蝗虫卵饲养,待幼虫生长到3龄时转移至人工饲料,直至化蛹。昆虫携带共生菌是一种普遍存在的自然现象,在长期的协同进化过程中,昆虫与其内共生菌之间形成了稳定的互惠共生关系[1-3]。昆虫和其肠道微生物处在一个共生的关系中,大多数共生菌对昆虫的生长发育、食物消化、免疫等具有一定的影响。肠道作为昆虫消化系统的重要组成部分,其中的共生菌对昆虫食性的选择和食物的降解代谢具有重要作用[4-10]。因此,揭示眼斑芫菁幼虫肠道多样性与食物之间的关系,对眼斑芫菁的人工饲养具有重要意义。【前人研究进展】斑蝥素的应用范围日趋广泛,对斑蝥素资源的需求愈来愈大,但不合理采集导致野生斑蝥资源日渐减少。芫菁科昆虫资源匮乏已成为制约进一步开发利用斑蝥素的“瓶颈”,人工饲养斑蝥已成为必然趋势。20世纪90年代,科研人员就对斑蝥的人工饲养进行了研究,明确了斑蝥生长的最适温度、湿度、光照条件、饲养密度等[11-12],汪会荣[13]研制了斑蝥成虫和幼虫的人工饲料配方,目前大多数研究相对集中在斑蝥素的合成、应用等,同时也开展了芫菁与外寄生菌的研究,周治成等[14]基于扫描电镜观察和分子生物学试验对一株扁角豆芫菁(EpicautaimpressicornisPic )寄生菌进行了分离鉴定,结果显示为饲养过程中感染的真菌黄曲霉菌;李晓飞等[15]对芫菁寄生菌降解斑蝥素进行了研究,发现球孢白僵菌可以有效降解斑蝥素,而曲霉不能降解斑蝥素。【本研究切入点】近年来,昆虫肠道共生菌和寄主昆虫的互作逐渐成为国内外关注的热点[16],取食人工饲料后,斑蝥肠道共生菌可能在提供营养方面起着重要作用,进而影响其新陈代谢和生长发育,但斑蝥肠道菌群的相关研究尚未见报道。【拟解决的关键问题】本研究以眼斑芫菁为研究对象,通过高通量测序技术分析其肠道内共生菌的多样性,初步探索斑蝥肠道内共生菌的组成差异以及功能,为后续针对眼斑芫菁或其它鞘翅目昆虫肠道微生物的研究提供依据。
1 材料与方法
1.1 眼斑芫菁试虫的采集与实验室饲养
供试的眼斑芫菁成虫采自贵阳省毕节县,采回实验室内用丝瓜花饲养,接初孵幼虫以蝗卵饲养,至2龄时,采集样品作对照;一部分用蝗卵继续饲养至4龄采样;另一部分2龄幼虫转接至人工饲料饲养,至4龄采集样本,样品信息如表1所示。

表1 样品信息Table 1 Information of the sample
1.2 眼斑芫菁试虫的饥饿处理与表面杀菌
将收集的眼斑芫菁试虫转移至新的离心管中,在饲养环境下饥饿处理24 h,使眼斑芫菁试虫体内的肠道菌群达到相对稳定的水平。用无菌水清洗饥饿处理后的试虫,再用75%乙醇浸泡2 min,再次用无菌水冲洗去除残留的乙醇。
1.3 眼斑芫菁试虫的解剖与肠道收集
经过饥饿处理和表面杀菌后,在体视显微镜下用尖头镊子夹住眼斑芫菁试虫的虫体,用眼科剪沿着头口器下方的腹部,顺着胸足、腹足、尾足之间轻轻剪开,然后换另外一把镊子展开腹部,完整取出肠道,置于无菌离心管中,用液氮速冻, -80 ℃冰箱保存用于DNA提取。
1.4 眼斑芫菁试虫肠道共生菌群16S rDNA扩增子测序
采用CTAB法提取眼斑芫菁试虫肠道菌的基因组DNA,测定DNA浓度并用琼脂糖凝胶电泳检测DNA的完整性。PCR全长扩增,产物纯化,构建SMART bel文库,PacBio上机进行高通量测序。
1.5 数据分析
PacBio下机数据通过PacBio公司SMRT分析软件导出bam文件。按barcode区分样本后进行OTUs(Operational taxonomic units )聚类和物种分类分析。根据OTUs聚类结果,对每个OUT的代表序列做物种注释,得到对应的物种信息和基于物种的丰度分布情况。同时,对OTUs进行丰度、Alpha多样性计算及Venn图分析,得到样品内物种丰度和均匀度信息、不同样品或分组间的共有和特有OTUs信息等。对OTUs进行多序列比对并构建系统发生树,进一步得到不同样品和分组的群落结构差异,通过PcoA 降维图和样品聚类树进行展示。为进一步挖掘分组样品间的群落结构差异,选用LefSe、MRPP等统计方法对分组样品的物种组成和群落结构进行差异显著性检验。
2 结果与分析
2.1 眼斑芫菁幼虫肠道菌群测序质量与OTUs分布
本试验共检测12个眼斑芫菁幼虫肠道样本(表2),获得96 390条原始数据(Raw reads),最终获得81 759条有效数据(Clean reads),原始碱基数Base(nt)共118 280 001 bp,序列相似性为85.2%,有效数据比例均超过80%,说明测序数据质量满足后续分析要求。聚类共获得230个OTUs,从L.H2、L.H4和L.S4肠道菌群测序数据中获得121、106和48个OTUs。从图1可知,蝗卵饲养的眼斑芫菁幼虫的肠道细菌所含OTUs种类明显高于从2龄转接到人工饲料中的眼斑芫菁幼虫,说明蝗卵饲养的眼斑芫菁幼虫肠道细菌种类更丰富。
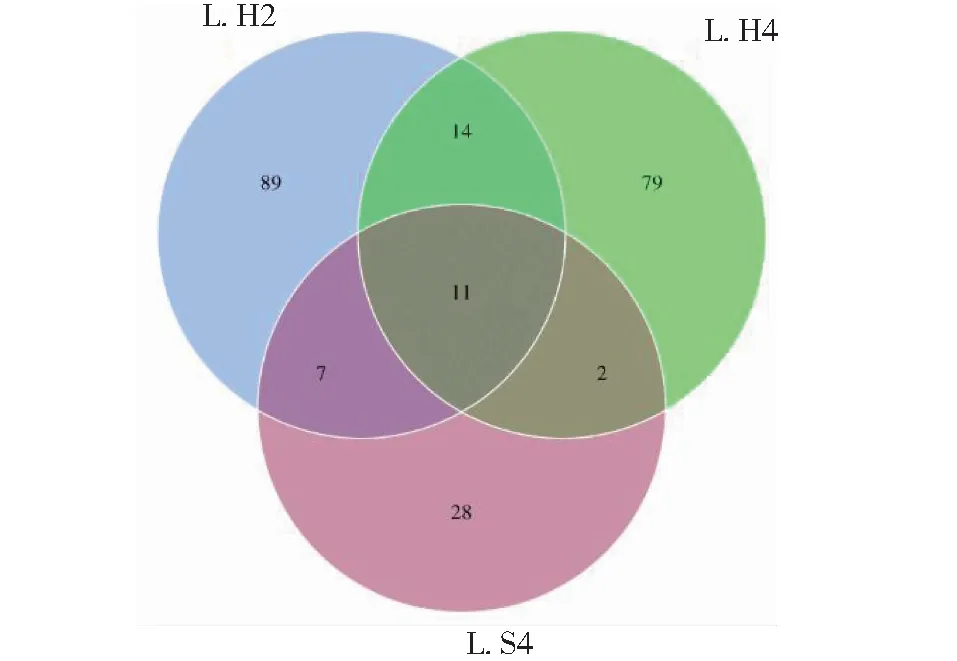
图1 不同组眼斑芫菁幼虫肠道内生菌Fig.1 Intestinal bacteria of different groups of M. cichorii Linnaeus larvae
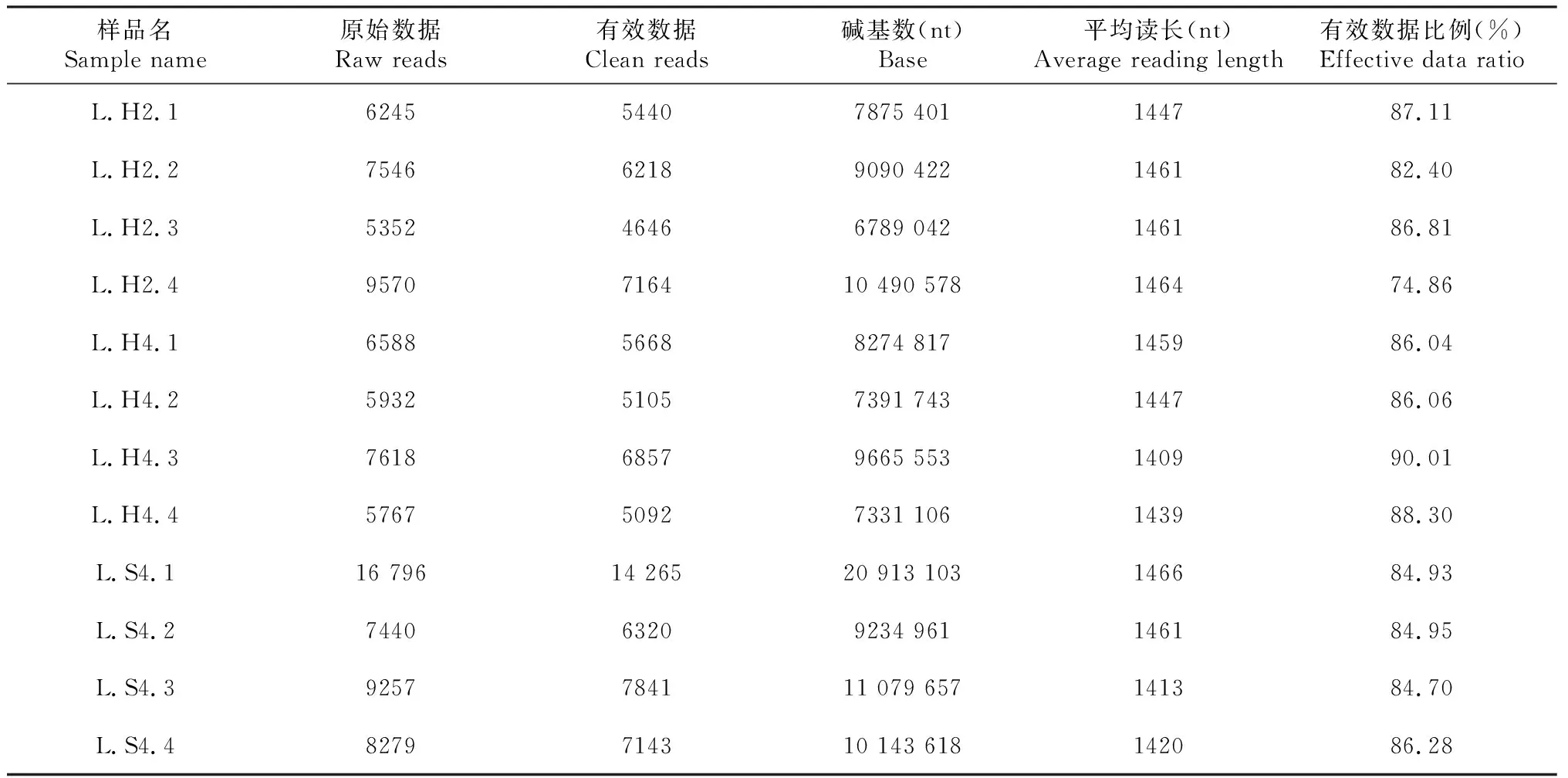
表2 眼斑芫菁幼虫肠道菌的测序预处理统计及质控信息Table 2 Sequencing preprocessing statistics and quality control information of gut microorganisms from larvae of M. cichorii
2.2 眼斑芫菁幼虫肠道内生菌的组成与结构分析
在门水平上,眼斑芫菁幼虫的肠道内生菌共注释到17个门(图2),主要为变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、拟杆菌门(Bacter-oidetes)、厚壁菌门(Firmicutes)等。3个处理组中,变形菌门(Proteobacteria)为肠道菌优势菌群的绝对优势菌门,L.H2、L.H4、L.S4的相对丰度分别为89.2%、92.7%、99.6%。
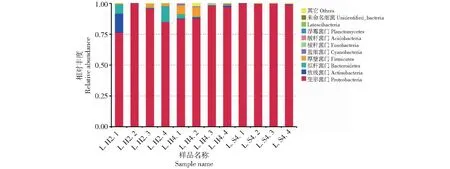
图2 眼斑芫菁幼虫肠道菌群门水平相对丰度前10的物种组成Fig.2 Bacterial composition of the top 10 relative abundances at the phylum level of M.cichorii Linnaeus larvae
在属水平上,眼斑芫菁幼虫的肠道内生菌优势属菌共注释到109个属,主要为劳尔氏菌属(Ralstonia)、甲基杆菌属(Methylobacterium),还包括沙雷氏菌属(Serratia)、Pluralibacter属、克雷伯菌属(Klebsiella)、不动杆菌属(Acinetobacter)、寡养单胞菌属(Stenotrophomonas)、鞘氨醇杆菌属(Sphingobacterium)、魏斯氏菌属(Weissella)、微杆菌属(Microbacterium)(图3)。各分组样品的优势菌属不同,L.H2肠道细菌样品中的优势菌属是克雷伯菌属(26.0%)、劳尔氏菌属(25.0%)、沙雷氏菌属(18.8%);L.H4肠道细菌样品中的优势菌属是甲基杆菌属(42.1%)、劳尔氏菌属(29.0%);L.S4肠道细菌样品中的优势菌属是劳尔氏菌属(24.8%)、Pluralibacter(18.7%)。
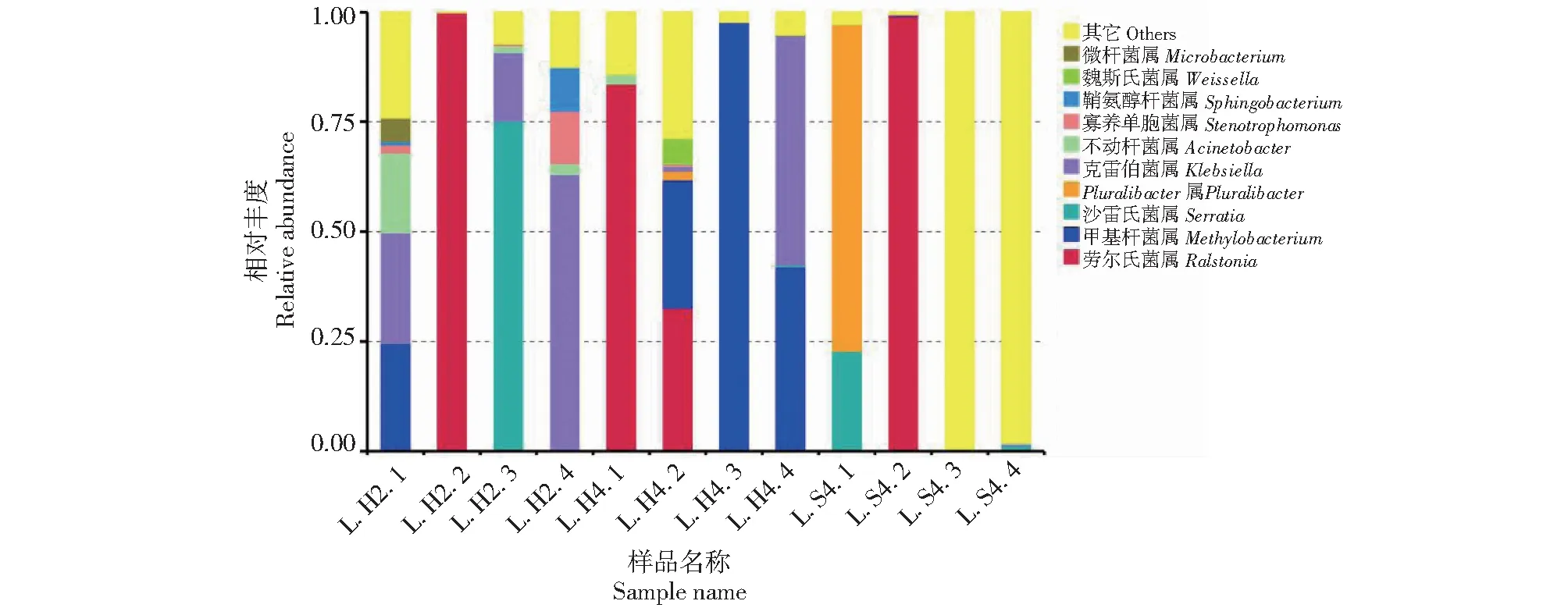
图3 眼斑芫菁幼虫肠道菌群属水平相对丰度前10的物种组成Fig.3 Bacterial composition of the top 10 relative abundances at the genus level of M.cichorii Linnaeus larvae
2.3 眼斑芫菁幼虫肠道菌群多样性分析
从表3可知,L.H2、 L.H4的Chao1指数分别为48.52、34.63,均比L.S4的(24.24)高,说明蝗卵饲养的眼斑芫菁幼虫的肠道菌群丰度比人工饲料饲养的高。L.H2、L.H4的Shannon指数(1.80和1.47)比L.S4的(0.45)高;且L.H2、L.H4的Simpson指数(0.46和0.42)高于L.S4(0.14),表明用蝗卵饲养的眼斑芫菁幼虫的肠道菌群多样性高于人工饲料饲养的幼虫。说明,L.H2和L.H4具有较高的多样性指数和丰度指数及优势度指数,表明这2组样本中的细菌种类更丰富。
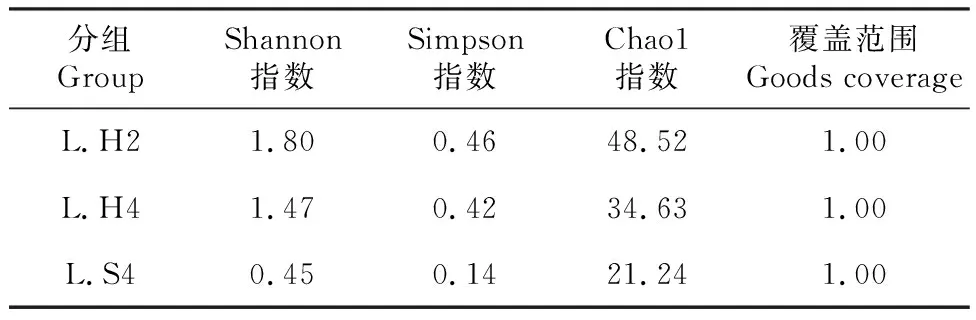
表3 眼斑芫菁幼虫肠道细菌α多样性指数Table 3 α diversity index of intestinal bacteria in M.cichorii larvae
利用Beta多样性分析中的MRPP分析配合PcoA降维分析显示,各组肠道菌群落结构组内差异大,但不同组的肠道细菌群落结构差异不显著(P>0.05),这与Alpha多样性分析结果一致。
2.4 眼斑芫菁幼虫肠道微生物群组成的差异分析
根据眼斑芫菁幼虫肠道微生物群在属水平的物种注释及丰度信息,选取丰度排名前35的属,根据其在每个样品中的丰度信息,从物种和样品2个层面进行聚类,绘制成优势菌属热图(图4),各处理眼斑芫菁幼虫肠道菌群的优势菌属差异明显。Pluralibacter菌属是转移到人工饲喂中的4龄眼斑芫菁幼虫肠道微生物群的主要组成部分,相对丰度高于其他组。
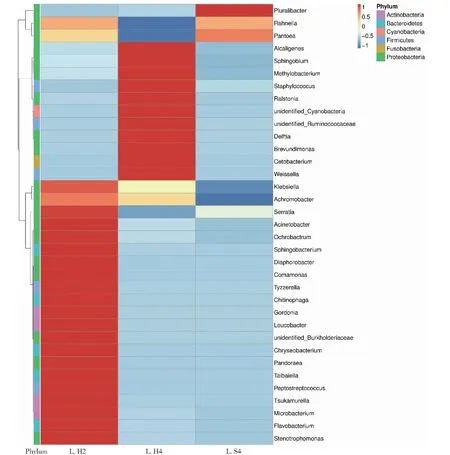
图4 眼斑芫菁肠道菌群属水平上相对丰度前35类群的聚类热图Fig.4 Cluster heat maps of the top 35 taxa with relative abundance at the genus level of M.cichorii Linnaeus larvae
LEfSe (Linear discriminant analysis effect size) 分析可以发现种间在丰度上具有显著差异的物种(图5)。L.H2肠道菌群中具有显著差异的菌群是未命名的伯克霍尔德菌属(unidentifiedBurkholderiaceae)、洋葱伯克霍尔德菌(Burkholderiacepacia)[17]和皮特不动杆菌(Acinetobacterpittii)[18-19];L.H4的肠道细菌丰度具有显著差异的为芽孢杆菌纲、乳杆菌目和鞘脂单胞菌目;L.S4的肠道菌群明显富集在α-变形菌目和醋酸杆菌科。
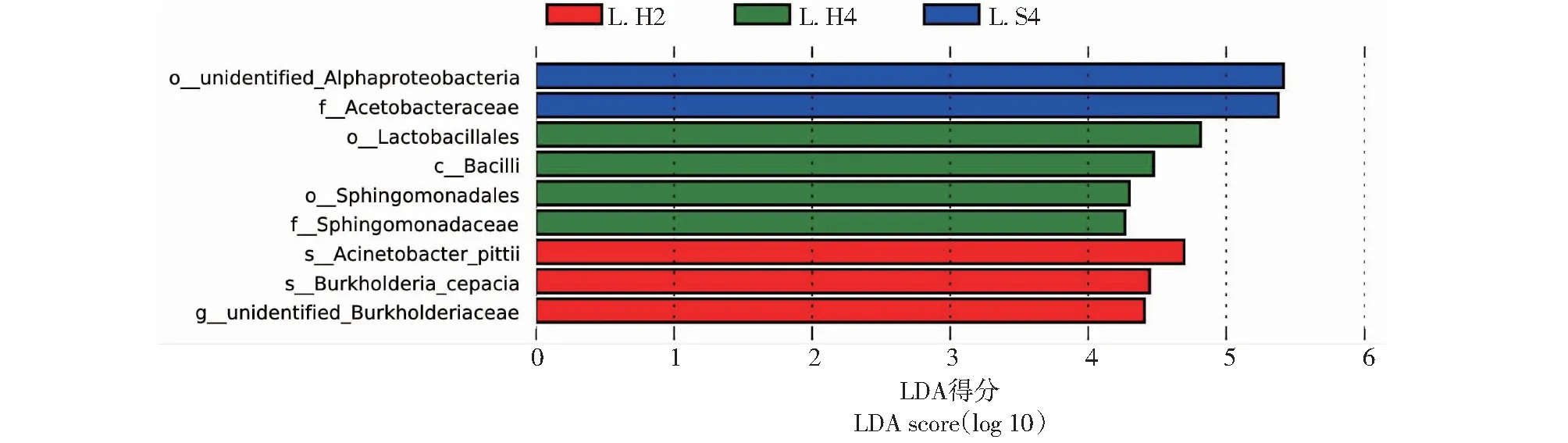
图5 肠道菌群线性判别(LDA)得分大于4的细菌类群Fig.5 Bacterial taxa with linear discriminant analysis (LDA) score greater than four in the gut microbiota
2.5 眼斑芫菁幼虫肠道菌群基因功能预测
利用PICRUSt软件对眼斑芫菁幼虫肠道菌群进行功能分析,得到不同样品细菌的功能预测信息。利用KEGG(Kyoto encyclopedia of genes and genomes)数据库,对眼斑芫菁幼虫肠道细菌基因功能预测发现,KEGG一级通路中共注释到6类生物代谢通路,由35个子功能组成,其中与新陈代谢相关的基因数量占总数的45.62%;参与环境信息处理(18.91%)、遗传信息处理(14.80%)、细胞工程(2.96%)、人类疾病(1.05%)、生物系统(0.54%)。对一级功能层分析发现,6个功能预测基因丰度无显著差异(P>0.05)。对预测基因的二级功能层分析发现,眼斑芫菁幼虫肠道菌群基因与11类消化代谢过程相关(图6),其中碳水化合物代谢和氨基酸代谢的基因在整个代谢过程中丰度最高,能量代谢以及辅助因子和维生素代谢的基因丰度也相对较高。进一步对预测基因三级功能层分析发现,L.S4的氨基酸代谢(Amino acid metabolism)比L.H4显著增强(图7,P=0.035)。
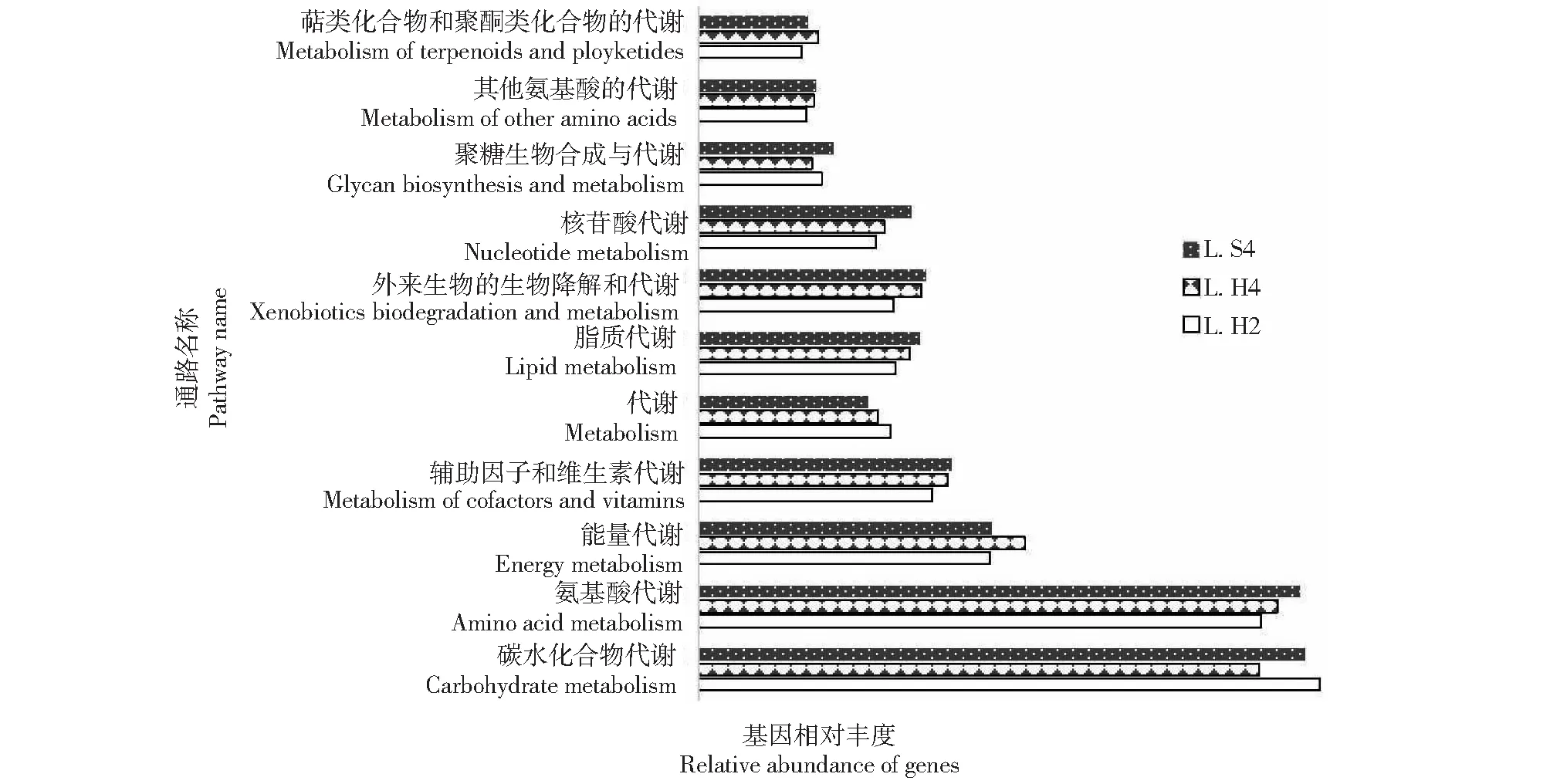
图6 Level 2水平的眼斑芫菁幼虫肠道菌群基因与代谢相关KEGG patnway注释Fig.6 KEGG patnway annotation on the correlation between intestinal flora genes and metabolism at Level 2 of M.cichorii larvae

图7 L.H4与L.S4组间差异显著的功能分析(三级功能层)Fig.7 Functional analysis of significant difference between L.H4 and L.S4 groups (Level 3)
3 讨 论
本研究对以天然食物为食的眼斑芫菁幼虫(L.H2、L.H4)及以人工饲料为食的眼斑芫菁幼虫(L.S4)的肠道菌群组成、多样性及丰度进行分析,共检测到17个门,27个纲,41个目,71个科,109个属。昆虫肠道菌结构组成的主要优势菌集中在变形菌门和厚壁菌门,尤其是鳞翅目[20-22],这一结论与本研究相同,眼斑芫菁幼虫肠道优势菌也是变形菌门(Proteobacteria),其相对丰度达到89.2%以上,因此,不同食性的昆虫肠道微生物具有相似性。
食物、健康状况、环境等因素可能影响昆虫肠道共生菌的种类和数量。在不同发育阶段,昆虫肠道内菌群丰度有一定差异,不同发育龄期的内生菌多样性也有差异[23]。本研究表明,以蝗卵为食的眼斑芫菁幼虫肠道菌菌群物种丰度及多样性差异较小,但显著高于以人工饲料为食的4龄幼虫,因此,可以初步设想,眼斑芫菁幼虫肠道菌群的多样性和丰度受食物影响较大。
不同寄主植物会引起植食性昆虫肠道微环境存在差异,进而导致其肠道微生物多样性的差异[24]。陈勃生等[25]研究发现,家蚕低龄幼虫与成虫肠道和卵的表面微生物相似,随着家蚕生长发育,高龄幼虫的肠道微生物与其食物(桑叶)表面的微生物更相近。本研究显示,具有相同食物来源的眼斑芫菁2龄幼虫与4龄幼虫的优势菌群不同,且2龄幼虫肠道菌群多样性和丰度比4龄幼虫高,可能是因为4龄幼虫取食以后,其肠道菌与食物中的微生物更接近,也证实食物可以影响眼斑芫菁幼虫的肠道菌群。
肠道细菌在昆虫宿主中的作用不可或缺,其功能包括帮助消化食物、参与营养吸收、激起免疫反应等[20, 26]。研究表明,有相当比例的昆虫依赖共生菌进行代谢,例如:白蚁(Nasutitermes)肠道细菌可帮助宿主降解纤维素[27],蚜虫(Acythosiphinpisum)共生菌能够合成氨基酸[28]。本研究中,眼斑芫菁幼虫肠道菌群代谢方面的基因注释较高,说明眼斑芫菁幼虫肠道菌群在代谢过程中起重要作用,特别是碳水化合物代谢和氨基酸代谢。L.S4的氨基酸代谢(Amino acid metabolism)比L.H4显著增强。说明,从蝗卵转移至人工饲料组饲养的眼斑芫菁幼虫肠道细菌会分泌多种消化酶协助宿主进行氨基酸代谢以及营养物质的吸收利用。
4 结 论
本研究明确了眼斑芫菁幼虫肠道菌多样性和群落结构受食物影响,其肠道细菌在代谢功能上能协助宿主适应相应的食物。研究眼斑芫菁幼虫肠道细菌多样性与食物之间的关系,为研究眼斑芫菁肠道微生物变化奠定了基础。今后的研究应从眼斑芫菁各个龄期、不同虫态肠道菌群的变化规律,或者是对不同食物的连续世代适应中的变化来进行研究,并用多组学方法剖析肠道菌在宿主适应中的重要作用。
