樟叶越桔肌动蛋白Actin7基因克隆与组织表达
2023-06-10蒲芝雨杨玉菊张安勉
蒲芝雨,杨玉菊,张安勉,赵 平,丁 勇
(1. 西南林业大学云南省高校林木生物技术重点实验室,昆明 650224; 2. 西南林业大学西南地区林业生物质资源高效利用国家林业与草原局重点实验室,昆明 650224)
【研究意义】樟叶越桔(Vacciniumdunalianum)为杜鹃花科(Ericaceae)越橘属植物[1],主要分布于中国西南地区,其干燥叶芽是云南省特色茶品“雀嘴茶”,富含氨基酸、Vc等多种营养成分[2]。同时樟叶越桔含有丰富的酚酸类次生代谢产物[3-4],具有重要经济价值,因而围绕活性成分生物合成相关功能基因的研究得以重视[5-6]。肌动蛋白(Actin)广泛存在于真核细胞中且进化高度保守,在真核生物细胞生理活动中发挥重要作用。研究樟叶越桔Actin基因的序列特征和组织表达情况,有利于探究樟叶越桔Actin家族基因调控机制,促进民族资源植物的开发利用。【前人研究进展】肌动蛋白(Actin)高度保守,其蛋白分子螺旋聚合成微丝,是细胞骨架的主要组分,由多基因家族编码,存在着不同的肌动蛋白异形体,也行使着不同的生理功能[7-8]。根据异形体的时空表达模式,Actin可分为生殖型和营养型[9]。在拟南芥(Arabidopsisthaliana)中ACT2能够使根毛在正确表皮细胞上形成突起[10]。ACT7可使叶绿体集聚以及正常形成愈伤组织[11-12]。随着分子生物学的发展,较多生物的Actin基因被相继克隆,如土人参(Talinumpaniculatum)[13]、山楂(Crataeguspinnatifida)[14]和红芪(HedysariRadix)[15]等。Actin家族基因中有的成员在不同组织和发育时期表达相对稳定,而被广泛作为内参基因用于其他功能基因的表达研究[16-17],也有部分Actin基因成员的表达存在组织特异性[18],因而Actin基因家族因成员不同存在着不同的组织和时空表达差异性,并发挥着不同的生物学功能。樟叶越桔富含酚酸类物质,其中以6′-O-咖啡酰熊果苷(6′-O-caffeoylarbutin,CA)含量最高[19],能有效抑制黑色素生成[20],且对α-葡萄糖苷酶和胰脂肪酶具有明显抑制作用[21]。【本研究切入点】樟叶越桔有望成为新型的抗糖尿、抗血脂及美白植物资源。而Actin蛋白作为生物体的重要组成成分,其基因及基因表达研究在樟叶越桔中未见报道。【拟解决的关键问题】 本研究基于转录组数据库从樟叶越桔中克隆了Actin7基因cDNA和组DNA序列,分析了该基因结构特征、密码子偏好性和组织表达特征,为深入研究樟叶越桔Actin家族基因和调控机制奠定基础。
1 材料与方法
1.1 试验材料
本试验植物材料选用云南省武定县大麦地村(25°45′ N;102°17′ E)的野生樟叶越桔植株,随机选取6棵植株,采取植株上幼嫩叶片、成熟叶片、花芽、花、绿色果实、红色果实、绿色果梗和红色果梗8种组织样品,6棵植株的每种组织混合放置于液氮中速冻,并带回实验室-80 ℃冰箱保存。
1.2 试验方法
1.2.1 引物设计与合成 以西南林业大学樟叶越桔资源创新与利用课题组前期获得的樟叶越桔三代转录组数据库为依据,筛选获得Nr、SwissProt和KEGG数据库均注释为Actin基因的cDNA序列,以编号VdSWFU1_c35521序列为基础,采用NCBI-Blast-Primer在线工具设计引物对,目的基因cDNA和组DNA的PCR扩增引物对为VdActin7-F1(5′-TTCCGTGAAATTCCGGTTCC-3′)和VdActin7-R1(5′-TCAGACTGCGAA CCACATCG-3′),qRT-PCR引物对为VdActin7-F2(5′-TTGATCTTGCTGGGCGTGAT-3′)和VdActin7-R2(5′-ACTTCCGGGCATCTGAATCG-3′),内参基因60S-2qRT-PCR引物对为60S-2-F1(5′-AGCACCTGAACCTGGATT-3′)和60S-2-F2(5′-GATAGCAGCG GTGGACTT-3′),上述引物委托生工生物(上海)有限公司合成。
1.2.2 核酸提取与检测 选用全能型植物RNA提取试剂盒[OminiPlant RNA Kit(Dnase I)]提取樟叶越桔8个样本总RNA,选用君诺德生物科技有限公司的CTAB植物基因组DNA快速提取试剂盒(离心柱型)从幼嫩叶片中提取樟叶越桔基因组DNA。总RNA和基因组DNA通过1%琼脂糖凝胶电泳和微量紫外分光光度计(NanoDrop2000)检测质量。选用反转录试剂盒(HiFiScript cDNA Synthesis Kit)将各组织的高质量总RNA (200 ng)合成单链cDNA备用。
1.2.3VdActin7基因扩增与克隆 以幼嫩叶片总RNA反转录的cDNA 2 μL为模板,与高保真酶PrimeSTAR Max Premix(2×) 25 μL、10 μmol/LVdActin7-F1和VdActin7-R1引物对各1 μL混合,用ddH2O补足至50 μL,得到VdActin7基因cDNA序列扩增的PCR体系;PCR反应程序为98 ℃变性10 s、57 ℃退火15 s、72 ℃延伸2 min,35个循环。以幼嫩叶片基因组DNA(200 ng)为模板,VdActin7-F1和VdActin7-R1为引物对进行VdActin7基因染色体DNA序列扩增,PCR反应体系和扩增条件同上。
PCR产物用1%琼脂糖凝胶电泳检测,用凝胶回收试剂盒(Gel Extraction Kit)回收纯化,用NanoDrop2000分析浓度与纯度。选用平粘末端克隆试剂盒(pClone007 Versatile Simple Vector Kit)将质量合格的PCR回收产物与克隆载体连接,42 ℃热激45 s,转化DH5α感受态细胞,涂布于含Amp抗性(50 mg/L)的LB固体培养基上,37 ℃培养16 h。随机挑选菌落于1 mL LB液体培养基(Amp+)中,37 ℃振荡培养12 h,利用菌液PCR筛选阳性菌落并送至北京擎科生物有限公司进行测序。
1.2.4 序列分析 利用NCBI(https://www.ncbi.nlm.nih.gov/)Blast程序对目的基因核苷酸、氨基酸进行同源性比对,Splign预测内含子和外显子,ORFfinder程序做开放阅读框分析,软件CondonW分析核苷酸组成,软件MAGA 11构建进化树,在线软件ProtParam(https://web.expasy.org/protparam/)分析其理化性质,SOPMA(https://npsa-prabi.ibcp.fr/)预测蛋白二级结构,SWISS-MODEL(https://swissmodel.expasy.org/)预测蛋白三级结构,SignalP 4.1 Server (http://www.cbs.dtu.dk/services/SignalP/)预测蛋白信号肽;软件CodonW和EMBOSS分析密码子偏好性、有效密码子数(Effective number of codons, ENC)、密码子适应性指数(Codon adaption index, CAI)和相对同义密码子使用度(Relative synonymous codon usage, RSCU)。
1.2.5VdActin7基因组织表达分析 以8种组织的cDNA分别为模板,选用TaKaRa的TB Green试剂盒配制qRT-PCR反应体系:TB Green Premix Ex Taq II(Tli RNaseH Plus)(2×)12.5 μL、10 μmol/LVdActin7-F2和VdActin7-R2各1 μL、cDNA 2 μL,ddH2O补至25 μL。反应程序为95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火30 s,72 ℃延伸30 s,40个循环;65~95 ℃(以0.5 ℃/5 s升温速度至95 ℃),升温过程中以每升高0.5 ℃采集荧光信号1次。内参基因为60S-2。每个样品设置3个生物学重复。以幼嫩叶片组织cDNA为模板进行qRT-PCR引物(VdActin7-F2和VdActin7-R2)的扩增效率分析。将cDNA依次稀释成5个梯度,其浓度分别为1、1/5、1/25、1/125和1/265倍。qRT-PCR反应体系和反应程序同上。
1.3 数据统计与分析
根据所得CT值绘制标准曲线,依据公式E(%)=(10-1/K-1)×100 获得引物的扩增效率。采用 2-ΔΔCT方法计算相对基因表达量,利用软件Microsoft Excel 2007 和 IBM SPASS 21.0分析目的基因表达情况和显著性并绘图。
2 结果与分析
2.1 总RNA及基因组DNA质量分析
樟叶越桔8种组织样品总RNA条带均清晰、无弥散(图1),提取的幼嫩叶片基因组DNA条带单一、明亮整齐(图2),表明核酸样品完整性好、无降解。总RNA和基因组DNA样品的A260nm/A280nm比值均在1.8~2.0内,证明核酸纯度好、无水解。试验提取的核酸满足后续试验要求。

1:绿色果梗;2:红色果梗;3:幼嫩叶片;4:成熟叶片;5:花芽;6:花;7:绿色果实;8:红色果实。1:Green fruit stem; 2:Red fruit stem; 3:Tender leaves; 4:Mature leaves; 5:Flower buds; 6:Flowers; 7:Green fruit; 8:Red fruit.图1 樟叶越桔8种组织总RNA琼脂糖凝胶电泳图Fig.1 Agrose gel electrophoresis of total RNA extracted from 8 tissues of V. dunalianum

M: DL15000 Marker; 1:基因组DNA。M: DL15000 Marker; 1: Genomic DNA.图2 樟叶越桔DNA琼脂糖凝胶电泳Fig.2 Agrose gel electrophoresis of DNA extracted from V. dunalianum
2.2 VdActin7基因克隆与分析
试验扩增获得目的基因约1400 bp cDNA和1600 bp DNA条带(图3),经克隆测序为1387 bp cDNA和1657 bp DNA序列。该基因DNA包含4个外显子和3个内含子,内含子1、2、3的长度分别为93、89、88 bp,均符合GU-AG内含子结构特征,其cDNA包括完整的ORF阅读框,长度为1134 bp,编码含有377个氨基酸的蛋白质。将氨基酸序列在NCBI数据库做同源性搜索,发现其与大多数植物的Actin7氨基酸序列相似性均在89%~100%,表明所克隆序列为Actin基因,将该基因命名为VdActin7(VacciniumdunalianumActin7)基因,并在GenBank注册,cDNA和DNA登录号分别为OM033722和OM069739。

M:DL2000 Marker;A:cDNA 扩增;B:DNA 扩增。M: DL2000 Marker; A: cDNA amplification; B: DNA amplification.图3 VdActin7基因cDNA (A)和基因组DNA (B)的PCR扩增Fig.3 cDNA (A) and genomic DNA (B) PCR amplification of VdActin7 gene
2.3 VdActin7蛋白理化性质分析
在线软件ProtParam分析结果显示,VdActin7蛋白质分子式为C1852H2920N492O559S21,分子量为41.70 kD,理论等电点(pI)为5.31,含有的20种氨基酸中Ala、Leu和Glu含量最高,占比23%。该蛋白不稳定系数为36.86(<40),表明为稳定蛋白。从亲疏水性分析结果(图4)可见,目的蛋白在第117位氨基酸,为最低峰值(-2.300);在第140位氨基酸,为最高峰值(2.200);亲水平均值为-0.163,表明其具有较强的亲水性。SignalP是信号肽及其剪切位点的预测工具[22],经SignalP预测发现,该蛋白无信号肽结构,属于非分泌蛋白质,且TMHMM预测结果显示,该蛋白不具明显的跨膜区(图5),为非膜结合蛋白。

图4 VdActin7蛋白亲疏水性预测Fig.4 Predicted hydrophilicity and hydrophobicity properties of VdActin7 protein
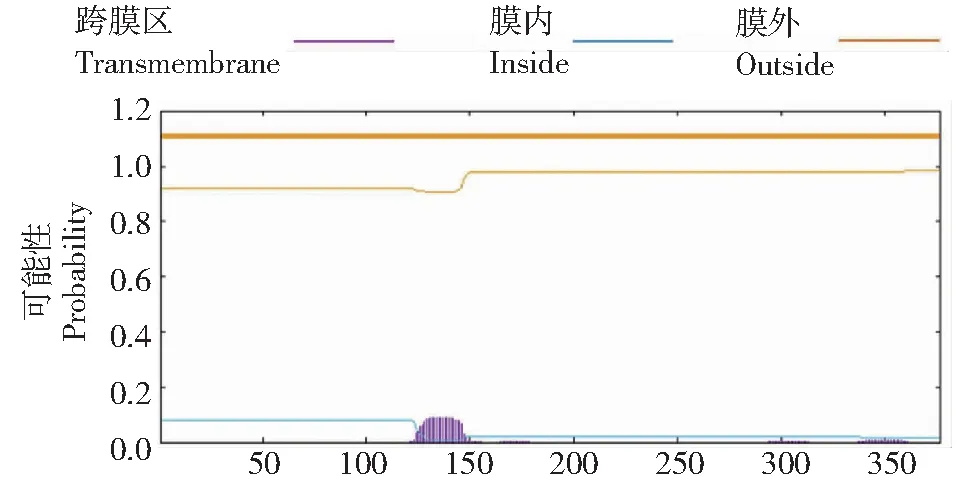
图5 VdActin7蛋白跨膜结构域预测Fig.5 Predicted transmembrane of VdActin7 protein
2.4 VdActin7 蛋白的二级和三级结构预测
二级结构预测结果(图6-A)表明,VdActin7蛋白中无规则卷曲35.81%、α-螺旋35.01%、延伸链22.02%、β-折叠7.16%,因此α-螺旋和无规则卷曲构成了该蛋白二级结构的主要骨架。保守结构域预测结果表明,其具有Actin家族典型的NBD_sugar-kinase_HSP70_Actin功能结构域特征(图6-B)。基于SWISS-MODEL同源建模的三级结构预测结果显示,VdActin7与玉米(Zeamays)花粉中F-Actin结构(6iug.1.A)具有最高的同源性(96.77%)。以6iug.1.A模型为模板建模得到VdActin7三维结构(图7),GMQE值为0.82,QMEANDisCo Global值为0.78±0.05,QMEAN Z值均大于-1.28,预测结果可信度高。

图6 VdActin7蛋白的二级结构(A)及保守结构域(B)Fig.6 Secondary structure prediction (A) and conserved domain (B) of VdActin7 protein
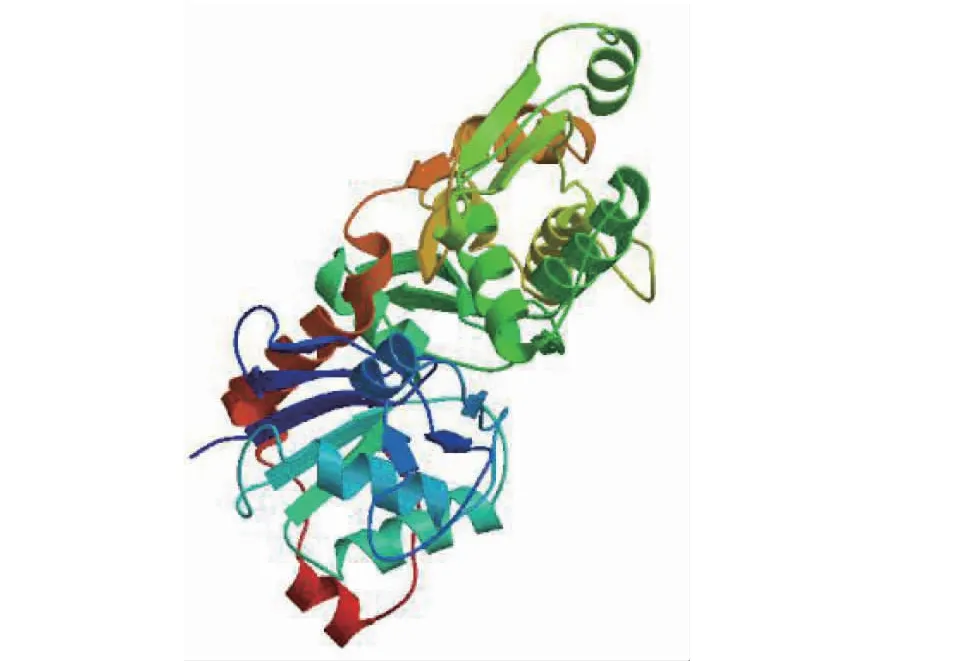
图7 VdActin7蛋白单体空间构象Fig.7 The conformation simulation of VdActin7 protein monomer
2.5 VdActin7基因同源性及系统进化树分析
将VdActin7基因完整ORF区编码的377个氨基酸序列与其他植物Actin的氨基酸序列进行比对。从图8可知,VdActin7蛋白与枣树(Ziziphusjujube,NP_001310807.1)、拟南芥(NP_196543.1)、黄瓜(Cucumissativus,XP_004147353.1)的Actin7和水稻(Oryzasativa,NP_001054419.1)Actin蛋白的同源性分别为100%、98.76%、99.73%和97.08%,且Actin蛋白均由377个氨基酸组成,表明植物的Actin蛋白在进化上具有高度保守性。

图8 樟叶越桔Actin7氨基酸序列与其他植物Actin氨基酸序列多重比对Fig.8 Multiple comparison of amino acid sequence of Actin7 from V. dunalianum and other plants
选用11个物种的Actin蛋白序列构建系统进化树。从图9可知,樟叶越桔与同为双子叶植物的兔眼越橘(Vacciniumashei)、茶树(Camelliasinensis)、枣树、大麻(Cannabissativa)、白梨(Pyrusxbretschneideri)、大豆(Glycinemax)、番茄(Solanumlycopersicum)和海岛棉(Gossypiumbarbadense)的Actin7蛋白共聚为一支。樟叶越桔与同为越橘属植物的兔眼越橘的Actin7蛋白亲缘关系最近,其次与茶树的Actin7蛋白亲缘关系较近。枣树和樟叶越桔不属于同目植物,但两者的Actin7蛋白氨基酸序列相似度为100%,故也聚为一支,说明Actin7蛋白在不同高等植物间极为保守。玉米和水稻同为单子叶植物,两者的Actin7蛋白在系统进化树中则聚为另一支,与VdActin7蛋白亲缘关系最远。

图9 樟叶越桔和其它高等植物的Actin系统进化树分析Fig.9 Phylogenetic tree of Actin from V. dunalianum and other species
2.6 VdActin7基因密码子偏好性分析
密码子具有简并性,在遗传信息的传递过程中发挥重要作用[23]。密码子偏好性分析结果显示,VdActin7基因总GC含量为48.2%,同义密码子第三位核苷酸含量(GC3s)为44.1%;CAI为0.261,ENC值为49.28%,表明密码子偏好性弱,密码子可以均匀利用。
樟叶越桔和枣树的Actin7蛋白氨基酸序列完全一致,但对应基因核苷酸的序列同源性仅有88.18%,因此进一步分析了2个基因的RSCU差异。由表1可见,2个物种Actin7基因的GC含量间存在差异,VdActin7基因的总GC含量(48.2%)高于ZjActin7(43.3%),而其GC3s含量(49.3%)则低于ZjActin7(51.1%);两者RSCU值大于1的高频密码子数分别有31和27个,2种植物Actin基因均对CUU的偏好性最强,GUU次之,但樟叶越桔对CUU的偏好性明显强于枣树。在VdActin7和ZjActin7基因中,Asp、Phe和Gln 3种氨基酸对应的简并密码子RSCU完全相同,Arg对应的简并密码子RSCU也几乎相同(除CGU),但其他具有简并性密码子的RSCU差异较大,表明简并密码子使用偏好性差异导致了编码产物完全相同的VdActin7和ZjActin7基因在核苷酸序列上的差异。
2.7 VdActin7基因表达分析
qRT-PCR试验中目的基因扩增溶解曲线呈现良好的单一信号峰,扩增效率在96.15%,线性回归方程为Y= -3.4179X+ 19.579(R2= 0.9849),表明引物特异性和扩增效率符合qRT-PCR试验要求。基因表达检测结果表明,VdActin7基因在不同组织中基因表达量不同,排序为嫩叶>成熟叶>花>红果果梗>花芽>绿果果梗>红果>绿果(图10)。该基因在叶组织、尤其是嫩叶中的表达量明显高于其他组织,暗示VdActin7蛋白在樟叶越桔叶片发育过程中可能发挥着重要作用,具体功能有待进一步研究。

不同小写字母表示差异显著(P<0.05)。Different lowercase letters indicate significant differences(P<0.05).图10 樟叶越桔Actin7基因在不同组织中的表达特征Fig.10 Expression characters of VdActin7 gene of different tissues from V. dunalianum
3 讨 论
高等植物的Actin蛋白由共同祖先进化而来[24],一般由375~377个氨基酸组成[25],在漫长的进化中其核苷酸和氨基酸水平呈现高度的保守性。本试验克隆了樟叶越桔Actin7基因全长cDNA序列和对应的DNA序列,该基因推导编码的377个氨基酸组成的Actin7蛋白具有Actin家族典型结构域NBD_sugar-kinase_HSP70_Actin,且与多种植物的Actin7蛋白具有极高的同源性。Actin蛋白在生物体内发挥重要的功能,在自然选择过程中承受着巨大的选择压力,因而氨基酸序列高度保守[26]。系统进化分析发现,樟叶越桔VdActin7与茶树CsActin7、兔眼越橘VaActin7和枣树ZjActin7聚在同一分支上,其中VdActin7与VaActin7亲缘关系最近,特别的是VdActin7和ZjActin7蛋白氨基酸序列完全一致,表明两者发挥着相同的生物学功能。但由于Actin7基因在进化过程中因简并密码子选择和使用偏好性不同,导致了该基因在樟叶越桔和枣树中的核苷酸序列的差异性。VdActin7基因密码子的总GC含量(48.2%)和GC3s含量(49.3%)符合已知双子叶植物中Actin基因的GC含量在45.0%~51.2%、GC3s含量在36.3%~53.8%的研究观点[27]。
植物中Actin基因可分为两类,一类参与细胞骨架构成,对维持细胞形态有重要作用,表达较为恒定,常常被作为内参基因使用[28];另一类则与植物生理功能密切相关,只在特定组织或特定时期表达[29]。已知研究表明,Actin7基因在拟南芥快速发育的幼嫩组织中高水平表达[30],在南方高丛蓝莓(Southern highbush blueberry)不同组织中表达不稳定[31]。本研究结果显示,Actin7基因在樟叶越桔不同组织中表达量差异显著,在叶片组织、尤其是幼嫩叶片中高水平表达,这与拟南芥和高丛蓝莓中Actin7基因的表达特征相似。因此推测Actin7基因在植物特定组织或特定时期特异性表达并发挥重要的生理学功能。细胞顶端生长的动力来自肌动蛋白和肌球蛋白的相互作用,所以在植物顶端区域存在大量肌动蛋白[32],推测Actin7蛋白作为营养型肌动蛋白在植物生长发育过程中发挥重要作用,但Actin7具体的生理功能还需进一步探究。本研究填补了樟叶越桔Actin蛋白的研究空白,丰富了高等植物Actin7蛋白信息,为进一步探究Actin7蛋白的功能、保守性以及进化关系提供了新依据。
4 结 论
本试验克隆了樟叶越桔VdActin7基因全长cDNA和DNA序列,该基因含有4个外显子和3个内含子,1134 bp的全长ORF区编码377个氨基酸组成的VdActin7蛋白。不同植物Actin7蛋白在氨基酸序列上具有极高的同源性,但基因密码子偏好性不同。VdActin7基因密码子的总GC含量和GC3s含量分别为48.2%和49.3%,对CUU的偏好性最强,GUU次之。VdActin7基因具有组织表达特异性,在叶片尤其是嫩叶组织中高水平表达。
