棉花GhRAR1基因调控棉花抗黄萎病机理的研究
2023-05-30胡晓倩石志钰朱玉涛高琳颖王平王宏伟侯玉霞
胡晓倩 石志钰 朱玉涛 高琳颖 王平 王宏伟 侯玉霞


摘要 棉花黄萎病是一种严重影响棉株生长的土传维管束真菌病害。挖掘抗性基因,解析抗病机制,对创制棉花抗病种质防御棉花黄萎病具有重要意义。本研究从抗病棉花品种‘中植棉2号中获得GhRAR1基因,采用病毒诱导的基因沉默(VIGS)技术、基因过表达及qRT-PCR技术探究GhRAR1介导的棉花抗黄萎病机制。结果表明,GhRAR1的表达受棉花黄萎病菌诱导。GhRAR1沉默棉株对黄萎病菌敏感性增强,表现为植株病情指数升高、叶片萎蔫、茎部维管束组织褐变加重。GhRAR1沉默棉株中活性氧(ROS)生成基因GhRbohD表达量低,H2O2积累量少。过表达GhRAR1基因的拟南芥植株对棉花黄萎病抗性增强,AtRbohD表达量高,H2O2积累量多,发生过敏反应(HR),菌丝扩散被抑制。推测GhRAR1可能调控ROS积累和HR,从而介导棉花对黄萎病菌的抗性。
关键词 棉花黄萎病; GhRAR1; VIGS技术; 抗病
中图分类号: S 423.1
文献标识码: A
DOI: 10.16688/j.zwbh.2021639
Abstract Cotton Verticillium wilt is a soil-borne vascular fungal disease, which seriously affects the growth of cotton. Mining resistance genes and exploring disease resistance mechanism are essential for innovating cotton disease-resistant germplasms and controlling cotton Verticillium wilt. Here, we cloned GhRAR1 from the disease-resistant cultivar ‘Zhongzhimian 2, and VIGS, overexpression techniques and qRT-PCR were used to explore the potential mechanism of GhRAR1-mediating defense against Verticillium wilt. The results showed that GhRAR1 was up-regulated in cotton plants inoculated with Verticillium dahliae. When GhRAR1 was knocked down, cotton plants were more susceptible to V.dahliae than the control, with the increase of plant disease index, leaf wilting and heavy browning of vascular bundle tissue in the stem. Furthermore, GhRAR1-silenced cotton seedlings showed lower expression of GhRbohD, and less accumulation of H2O2 than the control. When GhRAR1 was overexpressed in Arabidopsis thaliana, the plants exhibited enhanced resistance to V.dahliae. GhRAR1-overexpressed A.thaliana plants showed higher expression of AtRbohD, more accumulation of H2O2 than wild-type A.thaliana, and appeared hypersensitive response (HR), thereby inhibited the spread of hyphae. Taken together, our study indicates that GhRAR1 may be involved in regulating ROS accumulation and HR production for the resistance of cotton plants to V.dahliae.
Key words cotton Verticillium wilt; GhRAR1; VIGS; disease resistance
棉花黃萎病的病原为大丽轮枝菌Verticillium dahliae,常规化学防治方法难以实现有效防治[1],培育棉花抗病品种是解决这一难题的有效途径,因此研究抗性基因及其作用机制,为抗病育种提供基因资源显得尤为重要。
植物本身已进化出了复杂的免疫系统以应对病原菌的入侵,有许多抗棉花黄萎病的基因在棉花免疫系统中发挥着作用。例如,防卫基因GhPGIP1在棉花中过表达后,PRs、EDS1、PAD4和ICS1基因表达上调,促进木质部木质化,进而提高棉花的抗病性[2];漆酶基因GhLAC15受病原菌的强烈诱导,过表达GhLAC15后,棉花木质化程度增强、细胞壁中阿拉伯糖和木糖的积累增多,对黄萎病的抗性增强[3];编码富含亮氨酸重复序列的类受体蛋白基因Gbve1在陆地棉中过表达增强了对黄萎病菌的抗性[4];编码转录因子基因GhMYB108沉默后,棉花对棉花黄萎病菌的敏感性增强[5];SA信号通路的GhSAMDC基因[6],JA通路的GhCDKE基因[7],ET通路的GbABR1基因[8]等均正向调控棉花对黄萎病的抗性。
RAR1(required for Mla12-mediated resistance, RAR1)是一种锌结合蛋白,有2个富含半胱氨酸和组氨酸的保守结构域(cysteine and histidine rich domain, CHORD),参与R基因编码蛋白TIR-NBS-LRR和CC-NBS-LRR介导的防卫信号识别与传导,是植物抗病信号通路的重要组成元件[9-10]。RAR1被植物中的抗病基因触发后,激活下游免疫反应,如活性氧(ROS)暴发、过敏反应(HR)等,发挥抗病功能[11]。其中,ROS是植物识别病原菌的早期反应中最重要的信号之一[12],除其本身具有杀菌作用外,还作为诱发HR的信号分子发挥作用[13]。HR造成侵染部位坏死,限制病原菌菌丝扩散[14]。RAR1基因的功能在大麦Hordeum vulgare中首次被鉴定,研究发现RAR1突变体在病原菌触发的全细胞H2O2积累和细胞死亡中受到损害,表明RAR1在大麦抗性途径早期具有信号传导功能[9]。在拟南芥Arabidopsis thaliana中,RAR1被证明是R基因触发早期防御反应的限速调节因子,决定了对病原体的遏制程度[10]。据报道, SiRAR1基因在抗病品种中受谷锈菌Uromyces setariae-italicae诱导后上调更显著,可能正调控谷子抗病性[15]。目前为止,RAR1在棉花抗黄萎病中的作用机制尚不清楚。
本研究基于转录组学数据从‘中植棉2号棉花中获得GhRAR1基因。通过qRT-PCR技术分析了棉花GhRAR1基因的組织特异性表达模式和对棉花黄萎病菌的响应;分别调查了GhRAR1沉默棉花植株和稳定表达GhRAR1 的拟南芥植株对棉花黄萎病的抗性水平。其中棉株中GhRAR1基因沉默是通过VIGS技术实现的,该技术以病毒为载体接种到植株中,在当代植株中即可实现转录后沉默,操作简易、快速高效[16]。初步探究了GhRAR1参与棉花抗病的机理。
1 材料与方法
1.1 植物材料和真菌培养
‘中植棉2号种子由中国农业科学院棉花研究所惠赠,种子萌发后于培养箱中24℃、L∥D=16 h∥8 h条件下培养。拟南芥Arabidopsis thaliana哥伦比亚型(Col-0)种子在4℃春化3 d后,于L∥D=16 h∥8 h,光暗对应温度22℃/18℃条件下培养。
棉花黄萎病菌Vd991菌株在PDA培养基上于25℃培养7 d,然后将菌落转移到查氏液体培养基中培养7 d,获得孢子悬浮液,调整浓度为1×107个/mL用于浸根法[17]及叶片接种试验。
1.2 基因克隆
使用EASYspin 植物 RNA提取试剂盒(博迈德,北京)提取棉花 RNA。采用FastQuant cDNA 第一链合成试剂盒(天根,北京)将RNA反转录成cDNA。以cDNA为模板,根据棉花转录组数据设计特异性引物GhRAR1 (表1),进行PCR扩增,获得GhRAR1基因,基因序列经测序验证正确。
1.3 生物信息学分析
DNA序列使用Genetyx软件进行分析。通过MEGA-X软件构建系统进化树;使用Clustal Omega在线系统(https:∥www.ebi.ac.uk/Tools/msa/clustalo/)进行氨基酸序列比对;通过ExPASy (https:∥web.expasy.org/compute_pi/)预测蛋白分子量、等电点。
1.4 qRT-PCR 分析目的基因表达特征
设计qGhRAR1、 qGhRbohD、 qAtRbohD、 qVd特异性引物(表1),以GhUBQ7 (DQ116441)和AtEF-1-α (NM_100666.4)为内参基因。qRT-PCR 使用 SYBR Premix Ex Taq (Tli RNaseH Plus) 试剂盒以 20 μL 的终体积制备反应体系:SYBR Premix Ex Taq (Tli RNaseH Plus) 10 μL,10 μmol/L上、下游引物各0.4 μL,ROX Reference DyeⅡ 0.4 μL,50 ng/μL cDNA模板2 μL,RNase-free ddH2O 6.8 μL。在ABI7500 热循环仪(Applied Biosystems)上进行扩增,采用三步法反应程序,第一步:95℃ 30 s,循环1次;第二步:95℃ 5 s,60℃ 35 s,循环40次;第三步:95℃ 15 s,60℃ 1 min,95℃ 15 s,循环1次。每处理分别设3次生物学和技术重复,相对阈值法(2-ΔΔCt)计算相对表达量。
1.5 沉默载体构建和沉默植株的获得
采用病毒诱导的基因沉默(VIGS)技术进行基因沉默[16]。
棉花GhCLA1基因控制叶绿素合成,用GhCLA1沉默棉株做阳性对照。根据GhRAR1功能域序列设计引物VIGS-GhRAR1(表1),对‘中植棉2号cDNA进行PCR扩增,得到GhRAR1沉默片段。将沉默片段与VIGS载体PYL192连接,获得阳性单克隆菌液。提取阳性单克隆质粒,构建GhCLA1和GhRAR1的沉默载体TRV∷GhCLA1和 TRV∷GhRAR1。
将TRV∷GhCLA1和TRV∷GhRAR1质粒转化到大肠杆菌中测序,然后将序列正确的重组质粒通过热激法转入根癌农杆菌GV3101菌株[18],将空载体PYL192用同样的方法转入根癌农杆菌GV3101菌株中作为对照,用TRV∷00表示。将根癌农杆菌培养物注射到‘中植棉2号2周龄幼苗的子叶中。将幼苗放回培养箱中生长 2 周,获得沉默棉株[2]。
1.6 过表达载体构建和转基因拟南芥的获得
将GhRAR1基因和pCAMBIA1300载体(含潮霉素磷酸转移酶)经Pst I和Sal I酶切后连接,构建GhRAR1过表达载体。热激法将重组载体导入根癌农杆菌 GV3101 菌株,通过蘸花序法获得转基因拟南芥植株[19],将其产生的种子在含有25 mg/L潮霉素B的MS固体培养基上筛选,显示100% 潮霉素抗性的稳定表达株系用于后续试验[14]。
1.7 组织化学染色
DAB染色用以观察H2O2积累[20]。每片叶片上滴加10 μL的1×107个/mL Vd991孢子悬浮液,72 h后,并将其浸入 DAB-HCl 溶液(pH为3.8)中染色 6~7 h,脱色后用生物显微镜(Nikon)拍照。
台盼蓝染色用以观察菌丝和坏死细胞[21]。将叶片在台盼蓝溶液中煮沸1 min,过夜染色,随后用水合氯醛溶液(250%,m/V)脱色,生物显微镜(Nikon)拍照。
1.8 H2O2含量测定
使用 FOX 试剂法[22]测定H2O2含量。FOX试剂:250 μmol/L 硫酸亚铁铵、100 μmol/L 山梨醇、100 μmol/L 二甲酚橙、25 μmol/L H2SO4、1%无水乙醇。
1.9 病害调查和真菌生物量检测
通过观察植株表型并计算病情指数(DI)评估病害严重程度,用0~4级共5个等级对植株病情进行分级统计[23]。病情指数=∑(各级病株数×各级代表值)/(4×调查总株数)×100。
通過qRT-PCR对拟南芥中Vd991生物量进行量化[24]。将Vd991接种于拟南芥植株,接菌后14 d采集样品研磨,取100 mg细粉末提取基因组DNA,以DNA为模板,用qRT-PCR检测植株中Vd991生物量,引物为qVd-F/-R, 以AtEF-1-α(NM_00666.4)为内参基因,相对阈值法(2-ΔΔCt)计算相对表达量。
2 结果分析
2.1 GhRAR1基因特征
GhRAR1基因开放阅读框663 bp,编码221个氨基酸,包含2个锌结合蛋白功能域CHORD-Ⅰ和CHORD-Ⅱ(图1a)。GhRAR1蛋白的分子量为24.21 kD,等电点为8.46。将AtRAR1(AAF18433.1)、HvRAR1(AAF18432.1)、NtRAR1(AAM62409.1)、TaRAR1(AIX87838.1)与GhRAR1(Gh_D09G189700.1)氨基酸序列进行比对,构建系统发育树(图1b),发现GhRAR1与NtRAR1氨基酸,亲缘关系最近。
2.2 GhRAR1组织表达特征
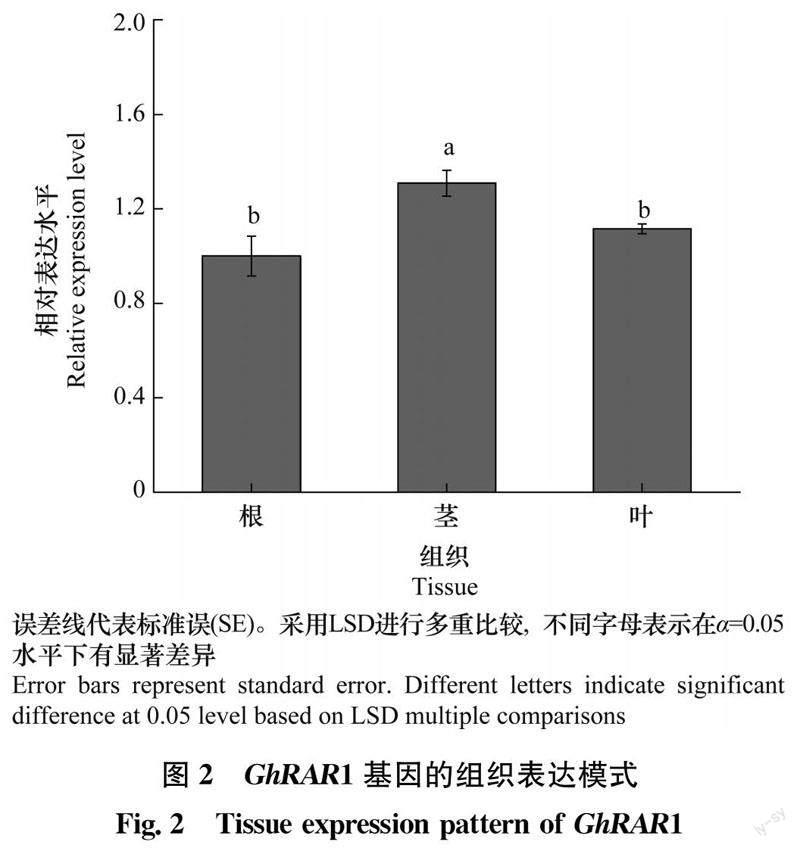
提取‘中植棉2号根、茎和叶的总RNA,反转录成cDNA,进行qRT-PCR定量分析,发现GhRAR1基因在棉花根、茎和叶3种组织中都有表达,在棉花茎部表达量最高(图2)。
2.3 大丽轮枝菌(Vd991)诱导GhRAR1表达
使用大丽轮枝菌强毒力菌株Vd991孢子悬浮液侵染棉株根部,利用qRT-PCR定量分析Verticillium dahliae菌株Vd991侵染后0、6、12、24、48、72 h 的棉花根部GhRAR1基因表达量变化。结果(图3)表明,GhRAR1表达受Vd991诱导,在24 h时GhRAR1表达量上调达峰值,约为0 h表达量的5.5倍。
2.4 GhRAR1基因沉默棉株对大丽轮枝菌的敏感性
利用VIGS技术沉默棉花中的 GhRAR1 基因。
VIGS处理2周时,GhCLA1 沉默植株出现明显白化(图4a)。
利用半定量RT-PCR检测棉株中GhRAR1基因表达水平,发现TRV∷GhRAR1植株的GhRAR1条带亮度明显比对照(TRV∷00)浅,说明 GhRAR1 基因沉默体系构建成功(图4b)。与此同时,用qRT-PCR检测对GhRAR1的沉默效率,结果表明,与TRV∷00植株相比,TRV∷GhRAR1中GhRAR1表达量显著下降(图4c)。接菌18 d后,与TRV∷00相比,TRV∷GhRAR1棉株对Vd991更加敏感,表现为更多的黄叶、枯萎(图4d),茎部出现更严重的维管束褐变(图4e),植株病情指数显著升高(图4f)。
2.5 GhRAR1基因沉默棉株接种大丽轮枝菌后H2O2积累量减少
浓度为1×107个/mL 的Vd991孢子悬浮液侵染TRV∷GhRAR1棉株和TRV∷00棉株后48 h,TRV∷GhRAR1棉株中ROS生成基因GhRbohD的表达量显著低于对照(图5a); H2O2积累量在侵染48 h和72 h时,均显著少于对照(图5b);
对接菌棉株叶片进行DAB染色,发现Vd991侵染72 h时,TRV∷GhRAR1棉株中被染色部分明显少于对照(图5c),与定量测定结果一致,说明GhRAR1沉默棉株接种大丽轮枝菌后H2O2积累量减少。
2.6 GhRAR1过表达拟南芥对棉花黄萎病的抗性增强
为探究GhRAR1基因在植物抵御黄萎病菌过程中的作用,构建植物表达载体pCAMBIA1300-GhRAR1,获得转GhRAR1拟南芥植株,共7个株系。
对这7个转基因拟南芥株系中GhRAR1基因进行定量,野生型拟南芥中并未检测到GhRAR1基因。选择GhRAR1稳定表达且表达量较高的拟南芥株系Line 1、Line 4和Line 5及野生型拟南芥,用浓度为1×107个/mL 的Vd991孢子悬浮液进行浸根处理,结果发现,处理后16 d,与野生型相比,转基因拟南芥叶片黄化、坏死等症状明显减轻(图6a),植株中大丽轮枝菌生物量显著降低(图6b),且接菌后18 d病情指数也明显低于野生型拟南芥植株。
2.7 GhRAR1過表达拟南芥接黄萎病菌后H2O2积累量增多
浓度为1×107个/mL的Vd991孢子悬浮液侵染GhRAR1过表达拟南芥植株后,植株中ROS生成基因AtRbohD的表达量明显高于野生型植株(图7a)。
H2O2积累量在侵染植株48 h和72 h时均显著高于野生型植株(图7b)。
2.8 转GhRAR1基因拟南芥植株产生过敏反应限制病原菌的扩散
用台盼蓝溶液对Vd991侵染2 d的拟南芥叶片染色,结果(图8)显示,GhRAR1过表达植株叶片存在明显的HR引起的细胞死亡,且HR的发生明显抑制了大丽轮枝菌菌丝的生长。而野生型拟南芥中菌丝生长更多、扩散更快。
3 结论与讨论
RAR1是抗病基因介导的抗性反应过程中的重要信号元件[25]。本研究从‘中植棉2号中克隆到GhRAR1基因,获得GhRAR1基因沉默的棉株和GhRAR1过表达的拟南芥植株。通过对不同植株进行黄萎病菌接种、病害调查、相关基因表达量检测和组织化学染色探究了GhRAR1基因在抗棉花黄萎病中的作用。
棉花GhRAR1具有植物RAR1蛋白的特征序列,含有2个CHORD结构域。进化树分析表明,GhRAR1与烟草NtRAR1 亲缘关系较近。本研究结果表明GhRAR1对黄萎病菌有响应,于Vd991处理后24 h其表达量达到峰值。据报道,AtRAR1突变以及沉默TaRAR1均导致植物抗病性出现不同程度的减弱[10,26]。为了进一步验证GhRAR1是否参与植物对棉花黄萎病菌的防御响应,本研究构建了GhRAR1沉默棉株和GhRAR1过表达的转基因拟南芥植株。结果表明,沉默棉株对棉花黄萎病敏感性增强;叶片枯黄、坏死程度明显加重,茎部维管束褐变加重。过表达GhRAR1基因的拟南芥植株对棉花黄萎病抗性增强。与野生型相比叶片枯黄、坏死程度明显减轻,大丽轮枝菌定殖量明显减少,表明GhRAR1在植物抗棉花黄萎病反应中起正调控作用。
RAR1被认为是R基因诱导的防御反应的限速因子,在ROS暴发的上游发挥作用[10],且ROS除本身具有杀菌作用外还作为信号参与抗病反应[13,27]。结果证实Vd991处理后GhRAR1基因沉默棉株的ROS生成相关基因RbohD的表达量和H2O2积累量均低于对照,与DAB染色结果一致;GhRAR1基因过表达拟南芥植株中RbohD的表达量和H2O2积累量均高于对照。此外,Vd991侵染后的组织化学染色结果表明,GhRAR1基因过表达拟南芥及时出现HR响应,有效抑制病原菌菌丝的扩散。表明GhRAR1基因可能通过影响H2O2的积累和ROS的暴发,影响HR的发生,在植株抗棉花黄萎病的早期反应中发挥作用。
参考文献
[1] FRADIN E F, THOMMA B P. Physiology and molecular aspects of Verticillium wilt diseases caused by V.dahliae and V. albo-atrum [J]. Molecular Plant Pathology, 2006, 7(2): 71-86.
[2] LIU Nana, ZHANG Xueyan, SUN Yun, et al. Molecular evidence for the involvement of a polygalacturonase-inhibiting protein, GhPGIP1, in enhanced resistance to Verticillium and Fusarium wilts in cotton [J/OL]. Scientific Reports, 2017(7):39840. DOI: 10.1038/srep39840.
[3] ZHANG Yan, WU Lizhu, WANG Xingfen, et al. The cotton laccase gene GhLAC15 enhances Verticillium wilt resistance via an increase in defence-induced lignification and lignin components in the cell walls of plants [J]. Molecular Plant Pathology, 2019, 20(3):309-322.
[4] ZHANG Baolong, YANG Yuwen, CHEN Tianzi, et al. Island cotton Gbve1 gene encoding a receptor-like protein confers resistance to both defoliating and non-defoliating isolates of Verticillium dahliae [J/OL]. PLoS ONE, 2012,7(12):e51091. DOI:10.1371/journal.pone.0051091.
[5] CHENG Huanqing, HAN Libo, YANG Chunlin, et al. The cotton MYB108 forms a positive feedback regulation loop with CML11 and participates in the defense response against Verticillium dahliae infection [J]. Journal of Experimental Botany, 2016,67(6):1935-1950.
[6] MO Huijuan, SUN Yanxiang, ZHU Xiaoli, et al. Cotton S-adenosylmethionine decarboxylase-mediated spermine biosynthesis is required for salicylic acid-and leucine-correlated signaling in the defense response to Verticillium dahliae [J]. Planta, 2016, 243(4): 1023-1039.
[7] LI Xiancai, PEI Yakun, SUN Yun, et al. A cotton cyclin-dependent kinase E confers resistance to Verticillium dahliae mediated by jasmonate-responsive pathway [J/OL]. Frontiers in Plant Science, 2018(9):642. DOI:10.3389/fpls.2018.00642.
[8] LIU Yujia, LIU Xin, LONG Lu, et al. GbABR1 is associated with Verticillium wilt resistance in cotton [J]. Biologia, 2018(73):449-457.
[9] SHIRASU K, LAHAYE T, TAN M W, et al. A novel class of eukaryotic zinc-binding proteins is required for disease resistance signaling in barley and development in C.elegans [J]. Cell, 1999, 99(4): 355-366.
[10]MUSKETT P R, KAHN K, AUSTIN M J, et al. Arabidopsis RAR1 exerts rate-limiting control of R gene-mediated defenses against multiple pathogens [J].The Plant Cell, 2002, 14(5): 979-992.
[11]DANGL J L, JONES J. Plant pathogens and integrated defence responses to infection [J]. Nature, 2001, 411(6839): 826-833.
[12]高武軍, 孙毅, 郭玉永, 等. 活性氧在植物抗病反应中的作用[J]. 中国农学通报, 1999, 15(3): 40-42.
[13]SOARES C, CARVALHO M E A, AZEVEDO R A, et al. Plants facing oxidative challenges — A little help from the antioxidant networks [J]. Environmental and Experimental Botany, 2019(161): 4-25.
[14]STASKAWICZ B J, AUSUBEL F M, BAKER B J, et al. Molecular genetics of plant disease resistance [J].Science, 1995, 268(5211): 661-667.
[15]白辉, 宋振君, 宋丹丹, 等. 谷子抗病相关基因SiRAR1的克隆及表达分析[J]. 农业生物技术学报, 2019, 27(12): 2091-2100.
[16]GAO Xiquan, WHEELER T, LI Zhaohu, et al. Silencing GhNDR1 and GhMKK2 compromises cotton resistance to Verticillium wilt [J]. Plant Journal, 2011, 66(2): 293-305.
[17]GAO Xiquan, LI Fangjun, LI Maoying, et al. Cotton GhBAK1 mediates Verticillium wilt resistance and cell death [J]. Journal of Integrative Plant Biology, 2013, 55(7): 586-596.
[18]DONG Yiyu, BURCH-SMITH T M, LIU Yule, et al. A ligation-independent cloning tobacco rattle virus vector for high-throughput virus-induced gene silencing identifies roles for NbMADS4-1 and -2 in floral development [J]. Plant Physiology, 2007, 145(4): 1161-1170.
[19]许红梅, 张立军, 刘淳. 农杆菌蘸花法侵染拟南芥的研究[J]. 北方园艺, 2010(14): 143-146.
[20]TORRES M A, DANGL J L, JONES J D G. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response [J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(1): 517-522.
[21]ZHU Yingfang, SCHLUTTENHOFFER C M, WANG Pengcheng, et al. Cyclin-dependent kinase 8 differentially regulates plant immunity to fungal pathogens through kinase-dependent and -independent functions in Arabidopsis [J]. The Plant Cell, 2014, 26(10): 4149-4170.
[22]HUANG Lihua, HU Chao, CAI Wei, et al. Fumarylacetoacetate hydrolase is involved in salt stress response in Arabidopsis [J]. Planta, 2018, 248(2): 499-511.
[23]WANG Ping, ZHOU Lin, JAMIESON P, et al. The cotton wall-associated kinase GhWAK7A mediates responses to fungal wilt pathogens by complexing with the chitin sensory receptors [J]. The Plant Cell, 2020, 32(12):3978-4001.
[24]ZHU Yutao, HU Xiaoqian, WANG Ping, et al.GhPLP2 positively regulates cotton resistance to Verticillium wilt by modulating fatty acid accumulation and jasmonic acid signaling pathway [J/OL]. Frontiers in Plant Science, 2021(12): 749630. DOI: 10.3389/fpls.2021.749630.
[25]AZEVEDO C, SADANANDOM A, KITAGAWA K, et al. The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance [J]. Science, 2002, 295(5562): 2073-2076.
[26]WANG Xiaojing, WANG Yaru, LIU Peng, et al. TaRar1 is involved in wheat defense against stripe rust pathogen mediated by YrSu [J/OL]. Frontiers in Plant Science, 2017(8): 156. DOI: 10.3389/fpls.2017.00156.
[27]CHEN Zhixiang, SILVA H, KLESSIG D. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid [J]. Science, 1993, 262(5141): 1883-1886.
(責任编辑:杨明丽)
