以桉树皮为原料的有机基质对植物苗期生长酶活性的影响
2023-05-30黎书会童清唐红燕杨建荣唐永奉李金才张建珠张燕平吴疆翀贺思腾孙启涛
黎书会 童清 唐红燕 杨建荣 唐永奉 李金才 张建珠 张燕平 吴疆翀 贺 思腾 孙启涛
关键词:桉树皮;育苗;腐熟程度;珍珠岩;酶活性
中图分类号:S311 文献标识码:A
目前最常使用、需求旺盛的有机基质泥炭土属于天然不可再生资源,其形成需要特殊的湿地条件和漫长的过程[1]。由于“地球之肾”——湿地环境具有不可替代的生态功能,对其保护也愈发广泛和深入,开采愈发受限,价格日益攀升[2]。反之,农林废弃物价格低廉、材料普遍易得且理化性质稳定,可作为原料用于批量加工有机育苗基质。以桉树皮及木屑为原料的有机育苗基质中含有大量纤维素、木质素以及内源生根抑制物质[3-4],经过堆肥技术以及特殊微生物的有效转化在释放养分的同时可促进抑制物的分解,使其理化性质与泥炭土相似,且养分含量不低于泥炭土,肥力优良,具有代替传统不可再生资源泥炭土用于育苗的潜力[5]。
碳氮代谢作为作物体内最基本的代谢途径[6],直接影响作物的生长发育、产量及品质[7]。碳氮代谢包括无机碳的同化、转运和积累,无机氮的还原、同化及有机含氮化合物的转化、合成等[8],在这一系列过程中碳氮代谢关键酶起着决定性作用[9]。目前,腐熟桉树皮多被用于某些菌菇的生长培育以及仅用作少量添加辅料[10-12],而用于作物的栽培育苗还较少,课题组前期研究表明腐熟桉树皮用于部分作物的育苗在生长量和生物量指标上可取得替代泥炭土的效果[5],而在桉树皮基质对作物碳氮代谢关键酶活性的影响方面尚无研究。本研究旨在揭示不同腐熟程度桉树皮复配不同比例珍珠岩对作物苗期育苗的生理影响,以期为有机育苗基质的后续发展积累经验和提供技术储备,以及变废为宝,减少木材加工剩余物对环境的破坏和污染,提高种苗品质,促进生态农林业的发展。
1 材料与方法
1.1 材料
1.1.1 试验区概况 试验区设置在中国林业科学研究院高原林业研究所(原资源昆虫研究所)大棚内。大棚位于10276E,256N,海拔1919.2 m。
大棚育苗试验开始于2019 年9 月初,止于11 月中旬,棚内日最高温度43℃,最低温度19℃,平均温度31℃,平均相对湿度64%,平均日光照时长11.4 h。
1.1.2 供试品种 3 种供试蔬菜植物种子购买自中国种子集团有限公司,分别为青花(碧玉西兰花Brassica oleracea L. var. italic Planch.)、生菜(奶油生菜Lactuca sativa L. var. ramosa Hort.)和番茄(‘粉冠2 号Lycopersicon esculentum Mill.)。多用途经济作物辣木(辣木良种Moringa oleifera Lam. cv‘. 中林3 号)种子收集自中国林业科学研究院高原林业研究所经济林栽培研究组种质圃。
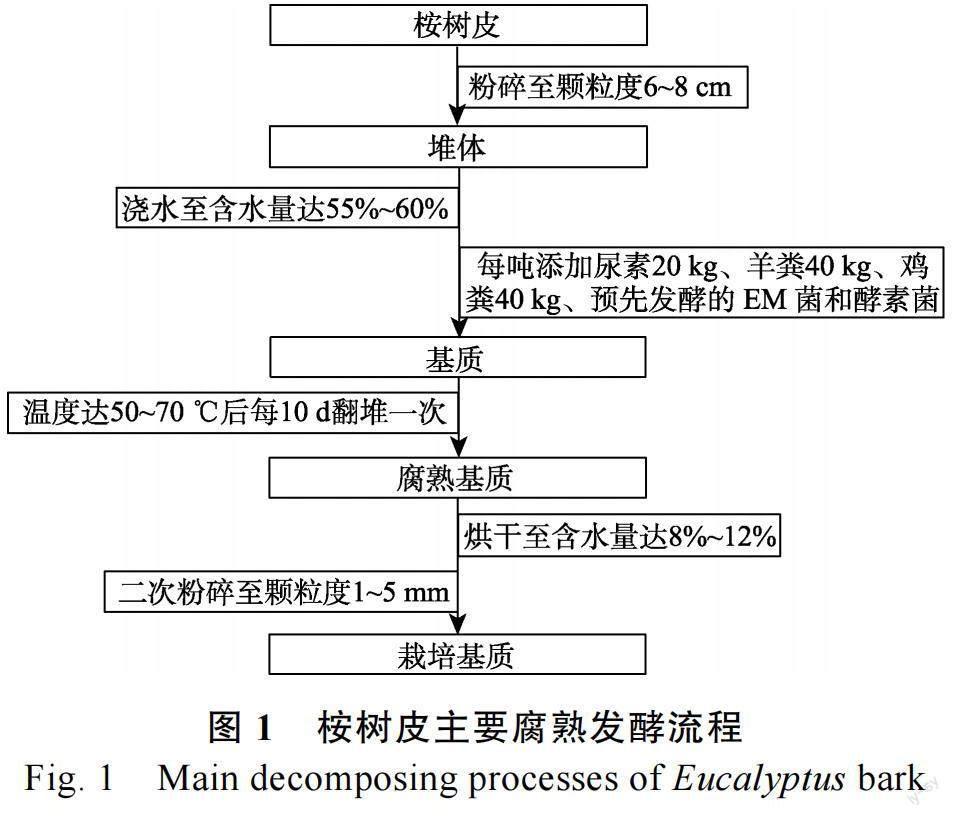
1.1.3 供试材料 (1)栽培基质。作为本试验栽培基质的原材料为尾巨桉(Eucalyptus urophylla ×E. grandis)树皮粉碎后纤维长2~8 mm 的生料,分别腐熟40 d 和60 d 的腐熟尾巨桉树皮基质,3种不同腐熟程度桉树皮均由云南省景谷县林涛木业有限责任公司提供。腐熟发酵方法参见本研究组前期国家发明专利“以思茅松树皮为原料的有机育苗轻基质加工方法”(ZL 201210498290.8),如图1。
(2)其他。粒径为3~6 mm 的珍珠岩,市售市场占有率高且育苗效果口碑好的泥炭土(丹麦品氏Pindstrup,购自昆明斗南花卉农资市场)。
1.2 方法
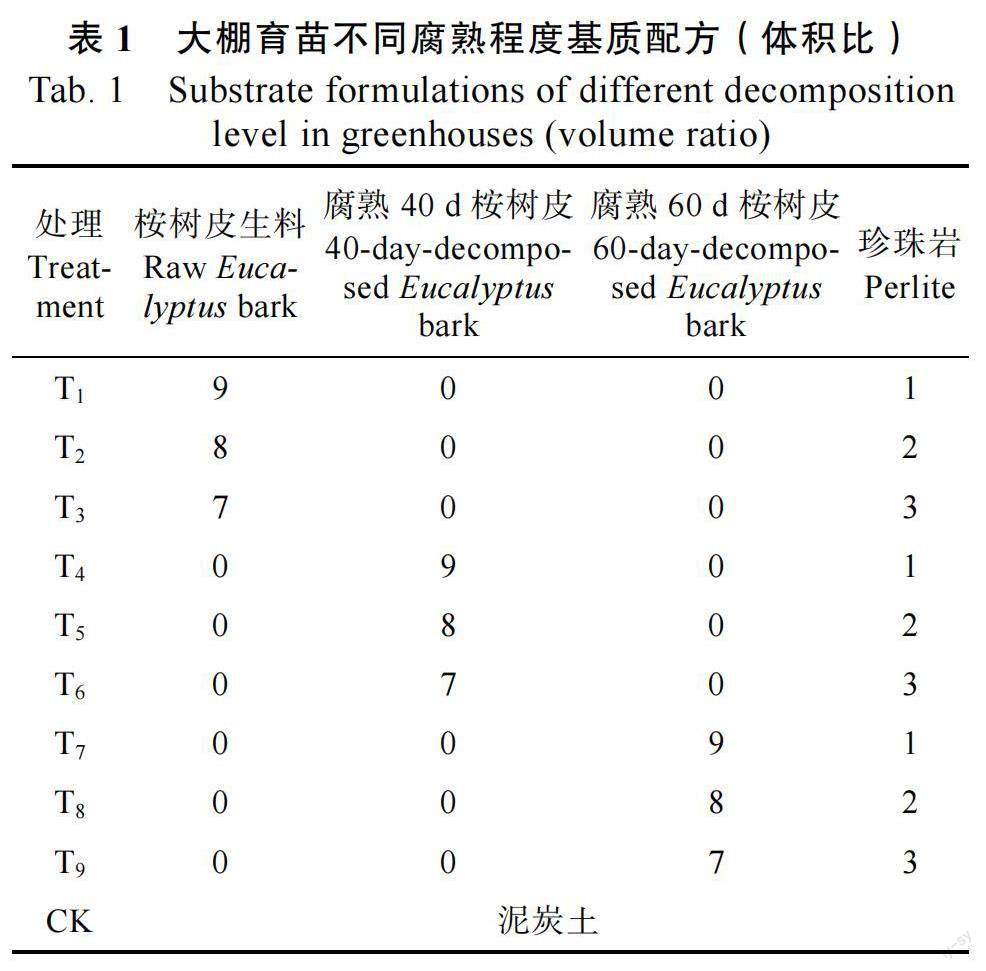
1.2.1 试验设计 采用两因素三水平完全随机设计,共设10 个处理,5 次重复。如表1 所示,其中T1~T3 为桉树皮生料,T4~T6 为腐熟40 d 桉树皮,T7~T9 为腐熟60 d 桉树皮。试验设1 个对照,为CK(泥炭土)。育苗方式采用12 孔漂浮育苗盒育苗,将以上充分拌匀的各个处理基质装入育苗盒内。将4 种供试植物:生菜(2 粒),番茄(2粒),青花(2 粒),辣木(1 粒)以不同粒数播入育苗盒的每个孔中,其中辣木播种前需要浸种15 h。每种植物各处理重复播种5 盒。每天进行浇水等日常管理。
1.2.2 采样 在生菜、番茄和青花播种后6 周,辣木播种后8 周开始取样,采样时间在上午9:00—10:00 進行,采样时用消毒剪随机摘取每个处理的30 株植株从顶叶往下的第3 至第5~8 片功能叶,立即用湿润的试纸擦拭干净后装入编号的自封袋,每个处理重复取样3 次。取完后放入?80 ℃超低温冰箱。
1.2.3 磨样 将冷藏的叶片剔除叶脉,置于已冷却的研钵中,加入液氮,将叶片研磨(加入少量石英砂)成粉末,后用5 mL 的离心管进行分装,分装时用万分之一的电子天平精确称取叶片粉末到相应质量,标记后保存于液氮中。
蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS):每份样品磨样0.2 g;核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco):每份样品磨样0.5 g;谷氨酰胺合成酶(GS)和谷氨酸合成酶(GOGAT):每份样品磨样0.5 g;硝酸合成酶(NR):每份样品磨样0.2 g。
1.2.4 指标测定 (1)SS 和SPS 活性:参考ZHU等[13]与MIRAJKAR 等[14]的方法测定。(2)Rubisco活性:参考CHENG 等[15]与CRISTINA 等[16]的方法测定。(3)GS 和GOGAT 活性:参考CHUAN等[17]的方法测定。(4)NR 活性:参考YU 等[18]的方法测定。
1.3 数据处理
采用Microsoft Excel 2010 软件对实验数据进行统计和处理;采用SPSS Statistics 23 软件进行方差分析和差异显著性检验(最小显著性差异法LSD);用SigmaPlot 12.5 和Adobe Photoshop CC2018 软件制图。
2 结果与分析
2.1 腐熟程度与珍珠岩对植物苗期6种酶活性的影响
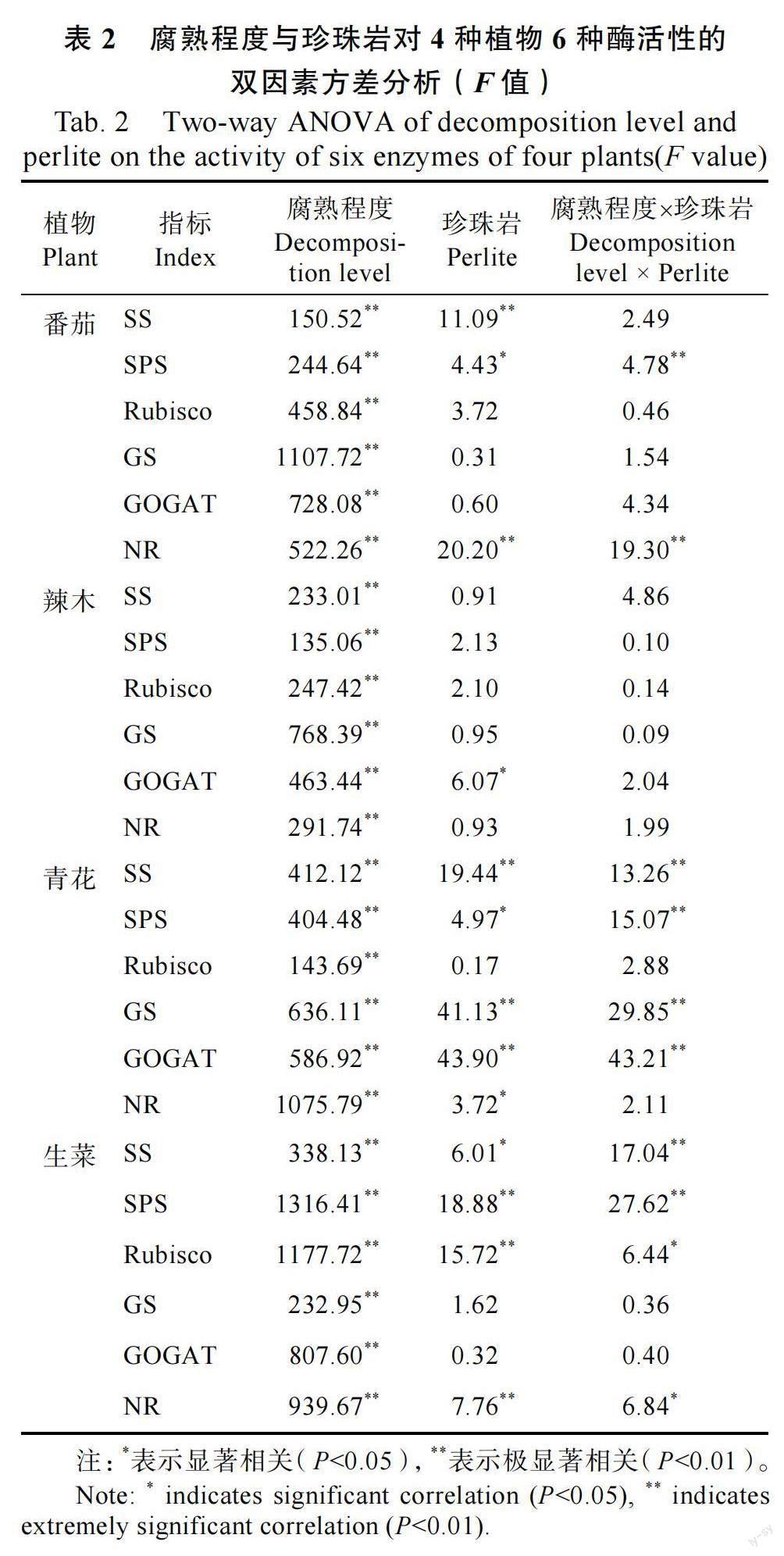
如表2 所示,以3 种不同腐熟程度桉树皮为原料的有机基质用于番茄、辣木、青花与生菜育苗,对这4 种植物苗期叶片中的6 种与碳氮代谢相关酶活性的影响均具有极显著差异(P<0.01)。而添加不同比例珍珠岩于基质中,仅对番茄叶片中的SS、NR 活性,青花叶片中的SS、GS、GOGAT活性以及生菜叶片中的SPS、Rubisco、NR 活性产生极显著影响(P<0.01)。桉树皮的不同腐熟程度与不同配比珍珠岩对番茄、青花与生菜的部分酶活性具交互影响,其中对这3 种蔬菜的SPS 活性均具极显著的交互影响(P<0.01)。
2.2 不同处理对碳代谢相关酶活性的影响
2.2.1 不同处理对SS 和SPS 活性的影响 番茄、辣木、青花与生菜叶片中SS 和SPS 活性对10 种处理基质的响应如图2。
番茄在10 种处理基质中SS 活性为0.057~0.107 U/mg,SPS 活性为0.003~0.015 U/mg。其中,番茄在T8中的SS 和SPS 活性均显著高于除T7之外的其他处理(P<0.05),T8与T7差异不显著。其次,SS 在T9与T5中的活性差异不显著,而SPS在T9中的活性显著高于T5(P<0.05)。这2 种酶在腐熟60 d 桉树皮中的活性均显著高于泥炭土(P<0.05)。
辣木在10 种处理基质中SS 活性为0.050~0.146 U/mg,SPS 活性为0.006~0.017 U/mg。辣木在泥炭土与生料中的这2 种酶活性均显著高于T6和3 种腐熟60 d 桉树皮处理基质(P<0.05),3个不同生料配比处理间无显著差异,但均显著低于泥炭土(P<0.05)。辣木在腐熟40 d 桉树皮中生长不良,在T4与T5中未生长出足够检测这2种酶活性的叶片。
青花在10 种处理基质中SS 活性为0.038~0.107 U/mg,SPS 活性为0.004~0.015 U/mg。其中在T8中的SS 活性显著高于其他处理(P<0.05),而在其他处理中的SPS 活性显著低于T9(P<0.05)。其次,SPS 在3 种腐熟60 d 桉树皮处理基质和T5中的活性均显著高于泥炭土(P<0.05),而SS 在T5与泥炭土中的活性无显著差异。
生菜在10 种处理基质中SS 活性为0.028~0.110 U/mg,SPS 活性为0.002~0.013 U/mg。其中,生菜在T9中的SS 活性显著高于除T7外的其他处理(P<0.05),而在T9 中的SPS 活性显著高于其他处理(P<0.05)。其次,SS 在3 种腐熟60 d桉树皮处理基质和T5中的活性均显著高于泥炭土(P<0.05),而SPS 在T5与泥炭土中的活性无显著差异。
2.2.2 对Rubisco 活性的影响 10 种处理对4 种植物叶片中Rubisco 活性的影响如图3。
番茄在10种处理基质中Rubisco 活性为4.44~10.12 U/g,在3 个腐熟60 d 桉树皮处理基质和T5中的Rubisco 活性均显著高于泥炭土(P<0.05),且3 个腐熟60 d 桉树皮处理间无显著差异。其次,T6和3 个不同生料配比处理基质与泥炭土间无显著差异。T4对番茄生长影响较大,未生长出足够检测Rubisco 活性的叶片。
辣木在10 种处理基质中Rubisco 活性为3.80~12.04 U/g。在泥炭土与生料中的活性均显著高于腐熟40 d 和腐熟60 d 桉樹皮(P<0.05)。3个不同生料配比处理间无显著差异,但均显著低于泥炭土(P<0.05),其次腐熟60 d 桉树皮显著优于T6(P<0.05)。T4与T5中缺少足够检测该酶活性的辣木叶片。
青花在10 种处理基质中Rubisco 活性为4.74~9.65 U/g。在T7与T9中的活性显著高于除T8外的其他处理(P<0.05),3 个腐熟60 d 桉树皮处理间无显著差异。T8与T5无显著差异,且均显著优于生料与泥炭土(P<0.05)。
生菜在10 种处理基质中Rubisco 活性为2.30~10.98 U/g。在3 个腐熟60 d 桉树皮处理基质中的活性存在显著差异(P<0.05),T9显著优于T8(P<0.05),T8显著优于T7(P<0.05)。其次,泥炭土中的活性显著低于腐熟60 d 桉树皮和T5(P<0.05)。生菜在生料中生长不良,其叶片中的Rubisco 活性均较低。
2.3 对氮代谢相关酶活性的影响
2.3.1 对GS 和GOGAT 活性的影响 4 种植物叶片中GS 和GOGAT 活性对10 种处理基质的响应如图4。
番茄在10 种处理基质中GS 活性为1.275~3.778 U/mg,GOGAT 活性为0.264~0.921 U/g。其中在3 个腐熟60 d 桉树皮处理基质及T5中的这2种酶活性均显著高于泥炭土与其他处理基质(P<0.05)。GS 在3 个腐熟60 d 桉树皮处理间的活性无显著差异,在3 个不同生料配比处理与T6中的活性显著低于泥炭土(P<0.05);而GOGAT在T8中的活性显著高于T9(P<0.05),且在T6中的活性与泥炭土无显著差异。
辣木在10 种处理基质中GS 活性为1.161~4.483 U/mg,GOGAT 活性为0.130~1.030 U/g。在泥炭土中的这2 种酶活性均显著高于所有桉树皮基质处理(P<0.05),其中生料显著优于腐熟40 d和腐熟60 桉树皮(P<0.05)。GS 在3 个不同生料配比处理中的活性无显著差异;而GOGAT 在T3中的活性显著低于另外2 个生料处理(P<0.05),且在T6中的活性显著偏低(P<0.05)。
青花在10 种处理基质中GS 活性为1.448~4.039 U/mg,GOGAT 活性为0.333~0.930 U/g。这2 种酶在3 个腐熟60 d 桉树皮处理基质和T5中的活性均显著高于泥炭土(P<0.05)。其中,GS 活性在3 个腐熟60 d 桉树皮处理间无显著差异,且均显著高于T5(P<0.05)。而GOGAT 在T9中的活性显著高于T8与T7(P<0.05),且T7 与T5无显著差异。
生菜在10 种处理基质中GS 活性为1.359~2.696 U/mg,GOGAT 活性为0.275~0.655 U/g。其中,GS 在3 个腐熟60 d 桉树皮处理基质和T5中的活性显著高于泥炭土和其他桉树皮处理(P<0.05),且3 个腐熟60 d 桉树皮处理间无显著差异;而GOGAT 在T5中的活性与泥炭土无显著差异。
2.3.2 对NR 活性的影响 10 种处理基质对4 种植物叶片中NR 活性的影响如图5。
番茄在10 种处理基质中NR 活性为0.003~0.016 U/mg。在3 个腐熟60 d 桉树皮处理和T5中的活性均显著高于泥炭土(P<0.05)。其中在T9中的活性显著高于其他9 个处理(P<0.05),其次T8显著高于T7(P<0.05),T7显著高于T5(P<0.05)。T4、T6与CK 间差异不显著,但均显著高于3 个生料处理(P<0.05)。
辣木在10 种处理基质中NR 活性为0.003~0.018 U/mg。在泥炭土中的活性显著高于所有桉树皮基质(P<0.05),其中在3 个不同生料配比处理中的活性均显著高于腐熟40 d 和腐熟60 d 桉树皮处理(P<0.05)。3 个不同腐熟60 d 桉树皮配比处理之间无显著差异,但显著优于T6(P<0.05)。
青花在10 种处理基质中NR 活性为0.004~0.013 U/mg。在腐熟60 d 桉树皮与腐熟40 d 桉树皮中的活性差异与在这2 种腐熟程度桉树皮中的GS 活性差异大致相同,不同的是,在CK 中的NR 活性显著高于T5(P<0.05)。
生菜在10 种处理基质中NR 活性为0.003~0.011 U/mg。在T9 中的NR 活性显著高于其他所有处理(P<0.05),而T7与T8的差异不显著,但均显著优于T5、CK 及生料(P<0.05),其次T5顯著优于CK 和生料(P<0.05),而CK 优于生料(P<0.05),3 个不同生料配比处理之间无显著差异。
3 讨论
蔗糖合成酶(SS)与蔗糖磷酸合成酶(SPS)是反映植物碳代谢强度的重要指标。SS 是蔗糖合成途径中的一个重要控制点,其活性反映蔗糖生物合成途径的能力[19-20],也是唯一能使蔗糖参与到组织构建、物质贮藏(淀粉合成)和植物细胞新陈代谢(糖酵解)等多种路径的酶[21]。而SPS主要作用于诱导蔗糖的合成,并在一定程度上反映“源”端供给能力[22-23]。核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)是光合作用中决定碳同化速率的关键酶,同时也是植物光呼吸的关键酶,其活性的高低反映了净光合速率的大小[24]。谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)和硝酸还原酶(NR)等被称为植株体氮素代谢的关键酶[25]。GS/GOGAT 循环是高等植物氮同化的代谢中心,二者的活性均能反映植物氮素同化能力的强弱[26]。NR 是植物氮代谢的首个限速酶,其活性可反映植物的氮利用率[27]。
本研究中,桉树皮生料中的3 种蔬菜叶片中的6 种与碳、氮代谢相关酶活性均显著低于腐熟处理基质,其形态表现在:3 种蔬菜生长至2~3周后叶片颜色发生变化,生菜叶片发黄近白色;部分青花和番茄幼苗不能长出真叶,且子叶由绿色转变为紫红色。这主要因生料中的主要养分(NPK)含量仅为腐熟40 d 桉树皮中的62%,腐熟60 d 桉树皮中的23%[5]。植株缺乏养分的供给,叶片中与碳、氮代谢相关酶活性偏低,从而表征到叶片上,这与前人的研究结果一致[28-29]。此外,生料中并未添加尿素、鸡羊粪以及菌种进行腐熟处理,其添加物对植株苗期相关酶活性是否存在直接影响有待进一步研究。而辣木由于具有发达胚乳给植株提供养分[30],且不耐受桉树皮经腐熟后释放的内源抑制物,如辣木的部分种子腐烂[5],因此生料中的6 种酶活性显著高于腐熟40 d 和60 d 桉树皮。3 种耐受桉树皮内源抑制物的蔬菜在腐熟60 d 桉树皮中与碳、氮代谢相关的6 种酶活性均显著高于泥炭土,这主要与二者的肥力与肥效有关,泥炭土中的主要养分(NPK)含量仅为腐熟60 d 桉树皮中的50%[5],番茄生长5 周后缺肥致叶片发黄,且泥炭土的肥效快而短暂,生菜与青花在泥炭土中播种后2~3 周出现茎的徒长现象,而腐熟60 d 桉树皮肥力高且肥效长,3 种蔬菜在其中生长良好,其中生菜又以复配30%珍珠岩的T9 处理效果更佳,说明较大的基质孔隙度更有利于生菜的营养生长。
有研究表明部分酚酸类化合物有促进种子萌发与幼苗生长的作用[31-34],本研究亦发现,3 种蔬菜在T4和T6中的6 种酶活性大部分相比泥炭土不显著,而在T5中的酶活性显著高于泥炭土,因复配不同比例珍珠岩对部分酶活性亦产生显著影响,因此桉树皮内源酚酸类物质与养分含量以及基质的物理特性都可能是致使T5 中酶活性高于泥炭土的原因。在泥炭土和不同腐熟程度处理组间,3 种氮代谢相关酶活性差异比3 种碳代谢相关酶活性差异大,说明氮代谢相关酶的活性受基质中养分含量以及桉树皮内源化合物的影响比碳代谢相关酶的影响更大,而桉树皮内源化合物对碳、氮代谢相关酶活性的影响机理有待进一步的研究。
