水稻烯酰辅酶A水合酶的功能预测分析
2023-05-30单崇蕾山草梅陈兰兰况卫刚林桠春张连虎崔汝强
单崇蕾 山草梅 陈兰兰 况卫刚 林桠春 张连虎 崔汝强


关键词:水稻潜根线虫;烯酰辅酶A 水合酶;抗病基因
中图分类号:S511 文献标识码:A
水稻是世界上最重要的农作物之一,全球超过一半以上的人口以水稻为主食[1]。为满足人口增长需要,据预测,至2030 年,水稻产量需要增加至少25%。随着测序技术的快速发展及基因组的易操作性,水稻已经成为一种重要的模式生物,用以研究植物性状及与病原微生物的互作。水稻生产中受到多种病原物的影响,寄生线虫是一类最具经济破坏性的植物病原体之一,每年导致全球产量损失1730 亿美元,因此,防控线虫病害是水稻生产中的一项重要工作[2-3]。
水稻潜根线虫是一类寄生于水稻根部的迁移性内寄生线虫,该线虫在世界水稻种植区如欧洲、亚洲和非洲等地均有被发现,分布极其广泛[4-6]。在我国水稻种植区,如广东、湖南、湖北、江西、福建等地,均发现有潜根线虫的为害[6]。水稻潜根线虫利用其口针穿刺水稻细胞,进入根表皮细胞,在根表皮细胞中取食迁移,造成细胞机械损伤。线虫的侵染会导致表皮细胞坏死,破坏皮层细胞,导致皮层和坏死区出现空洞,根部腐烂并呈现黄褐色;地上部分的为害表现为水稻伸长迟缓、株高降低、分蘖延迟、产量降低[7]。已有研究报道发现,有11 种潜根线虫可成功侵染水稻,其中水稻细尖潜根线虫是危害江西省水稻种植最严重的寄生线虫[4, 8]。
在本研究前期工作中,已对来自世界各地的560 个水稻品种进行水稻潜根线虫的抗性鉴定,并从中筛选出易感水稻品种‘霸王鞭(R155),同时对接种线虫的‘霸王鞭(RN155)和未接种线虫的‘霸王鞭(R155)进行了转录组测序[7]。结果发现‘霸王鞭品种中编码烯酰辅酶A 水合酶(enoyl-CoA hydratase, OsECH1)的一个基因Os01g0752200(LOC4324637)表达量显著上调,因此,该基因是否参与细尖潜根线虫的侵染,加强或降低水稻的抗病性是一个非常值得研究的问题。鉴于此,本研究以OsECH1 基因作为研究对象,对其结构特点、亚细胞定位及互作蛋白进行初步分析。同时获得OsECH1 基因的过表达及基因沉默的转基因水稻,进一步明确OsECH1 基因的功能,为进一步研究水稻抗潜根线虫的分子机理提供新方向。
1 材料与方法
1.1 材料
‘日本晴水稻种子、过表达载体pCAMBIA-1302、原核载体pET28a、分子伴侣pTf16、农杆菌GV3101、大肠杆菌感受态DH5α、水稻细尖潜根线虫均为本实验室保存。RNAi 载体pBWA(V)HS 及转基因水稻由武汉伯远生物科技有限公司提供。
1.2 方法
1.2.1 OsECH1 生物信息学分析 在NCBI 中進行BlastP 搜索OsECH1 在各种模式植物中的同源蛋白。选取的序列有黍( Panicum hallii, XP025814293.1)、柳枝稷(Panicum virgatum, XP039849084.1 )、小米( Setaria italica, XP004969966.1)、玉米(Zea mays, NP 001150290.1)、高梁(Sorghum bicolor, XP 002458497.1)、短柄草(Brachypodium distachyon, XP 003567122.1)、籼稻(Oryza sativa Indica Group, EEC 71489.1)、大麦( Hordeum vulgare subsp. Vulgare, XP044977061.1 ) 和小麦( Triticum aestivum, XP044341339.1, XP 044357827.1, XP 044350069.1)。根据得到的OsECH1 的同源蛋白,利用MEGA 7.0软件采用最大似然法(maximum likelihood)构建系统发育树,并进行1000 次的bootstrap 的分析验证。利用在线软件MEME 进行motif 分析( https://meme-suite.org/meme/tools/streme ); 在NCBI 中进行蛋白结构域的预测分析(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi);在NCBI 中对‘日本晴基因组文件下载,并完成对‘日本晴中ECH 家族的基因定位(https://www.ncbi.nlm.nih.gov/genome)。利用在线软件EXPASY对OsECH1 蛋白的理化性质进行分析(https://web.expasy.org/protparam/);利用在线软件EXPASYProtscale对OsECH1 蛋白的亲疏水性进行分析(https://web.expasy.org/protscale/);通过NetPhos3.1 Server 预测OsECH1 蛋白氨基酸序列的磷酸化位点情况(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)。
1.2.2 水稻材料的生长条件 水稻种子在28 ℃的无菌水中发芽3 d 后,移栽至营养土中(营养土壤∶珍珠岩∶蛭石=3∶1∶1),并在人工气候培养箱中培养,设置气温范围为28~30 ℃,相对湿度范围为85%~90%。
1.2.3 水稻RNA 制备和RT-qPCR 分析 利用RNeasy Plant Mini Kit(Qiagen)试剂盒提取水稻根总RNA, 利用EasyScript? One-Step gDNARemoval and cDNA Synthesis SuperMix(TRAN)试剂盒合成cDNA。在CFXTM 96 Real-Time PCRdetection system(Bio Rad)中进行实时定量PCR,使用PerfectStart Green qPCR SuperMix?(TRAN)完成RT-qPCR,其引物序列见表1,并使用2–ΔΔCT方法分析数据[9]。
1.2.4 OsECH1 亚细胞定位 构建pCAMBIA1302-OsECH1-GFP 载体用于亚细胞定位。使用电转法将质粒转入根癌农杆菌菌株GV3101 感受态中;转化完成后,将转化的细菌在含有抗生素LB 培养基中于28 ℃培养过夜;次日,取1 mL 过夜菌液以1∶100 的比例扩大培养,直至菌液在OD600=1~1.5 , 使用MES 缓冲液( 10 mmol/LMgCl2,10 mmol/L MES,pH 5.6)收集菌体重悬3 次并将悬浮液调整至OD600=0.25~0.5;将悬浮液注射至水稻原生质体中,并将材料在黑暗培养中保持24~48 h,最后使用激光共聚焦显微镜观察亚细胞定位的荧光信号。
1.2.5 OsECH1 互作蛋白的获得 将没有信号肽序列的OsECH1 蛋白的编码区插入至pET28a 载体中,并将其转入至含有pTf16 的感受态细胞中(本实验室保存)。构建成功的表达菌株于37 ℃涂布培养, 并挑取单菌落扩大培养至OD600=0.6~0.8。加入IPTG 诱导,超声后分别吸取上清、菌体,加入6×protein loading buffer 煮沸,进行SDS-PAGE 和Western blot 分析。对重组蛋白的表达条件优化后进行大量表达,用于后续的蛋白纯化。利用ProteinIso? Ni-NTA Resin 试剂盒获得纯化蛋白,所有步骤均按照说明书进行。诱导表达纯化和浓缩的蛋白与水稻根总蛋白孵育互作完成后,使用290 mmol/L 咪唑洗脱液洗脱互作蛋白,然后进行SDS-PAGE 凝胶电泳检测。蛋白质质谱检测由武汉金开瑞生物工程有限公司完成。
2 结果与分析
2.1 OsECH1 蛋白的生物信息学分析
根据水稻‘霸王鞭接种细尖潜根线虫的转录组数据分析,结果发现一个编码烯酰辅酶A 水合酶( enoyl-CoA hydratase, OsECH1) 的基因Os01g0752200(LOC4324637)在线虫侵染水稻时,表达量明显上调。从图1 可看出,水稻‘霸王鞭在被细尖潜根线虫侵染时,OsECH1 基因的表达量明显提高,说明该基因响应细尖潜根线虫的侵染。
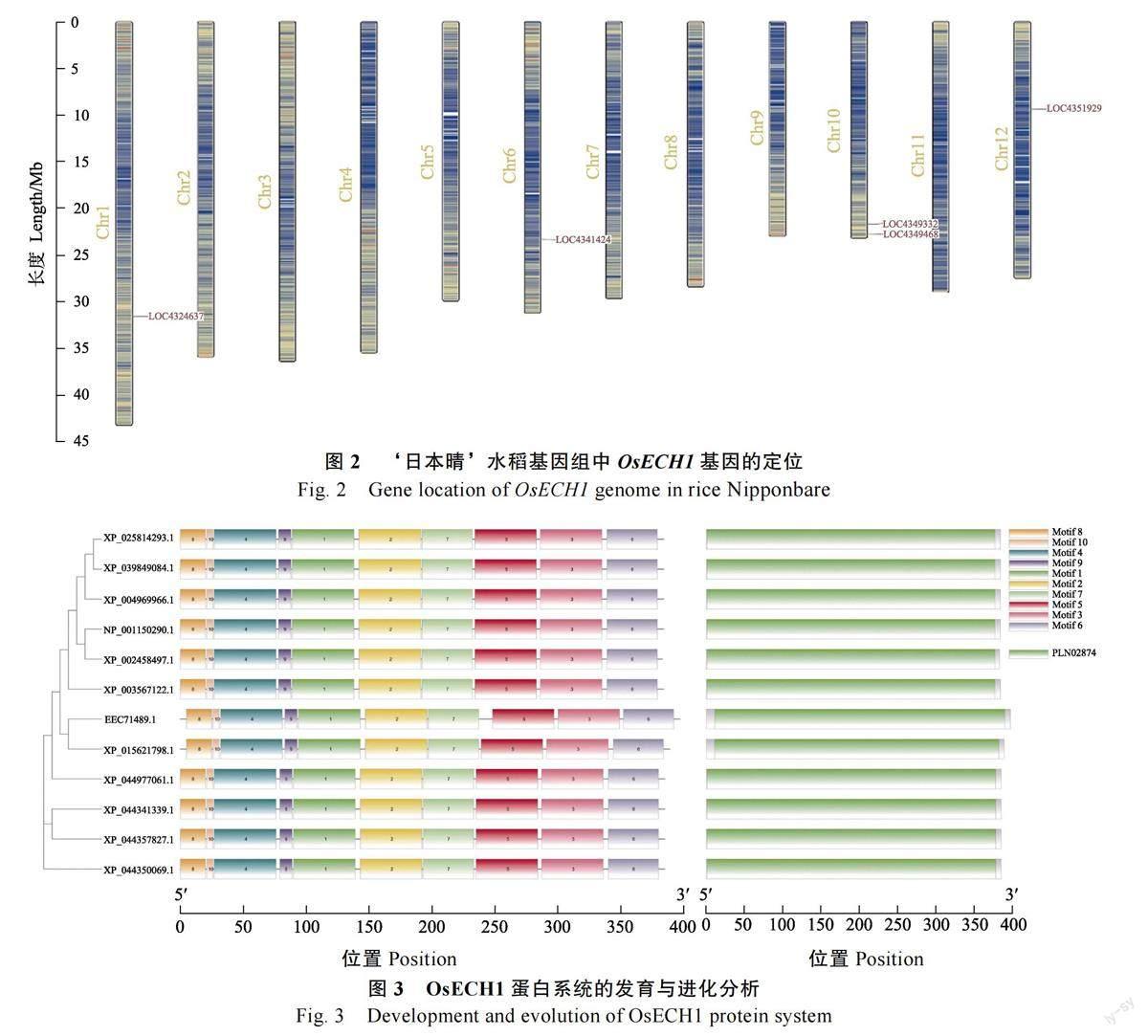
利用OsECH1 蛋白序列在水稻基因组中的序列比对分析,结果发现水稻基因组中有5 个同源基因,OsECH1 基因位于Chr1(染色体1)上(图2)。对来自其他8 个物种中的OsECH1 同源蛋白进行序列比对和系统发育分析, 结果表明,OsECH1 与籼稻(Oryza sativa Indica Group, EEC71489.1)更相似。此外,这些同源蛋白均具有相同的基序和保守结構域,这表明OsECH1 蛋白具有烯醇辅酶A 水合酶活性(图3)。
2.2 OsECH1 蛋白的理化性质分析利用DNAMAN 8 软件对OsECH1 基因序列及其编码氨基酸序列进行分析,结果显示,该基因ORF 为1170 bp,编码389 个氨基酸。利用在线软件ExPASy-ProtParam tool 分析OsECH1 蛋白的氨基酸组成及理化性质,结果表明,该蛋白分子式为C1915H3029N517O595S15,相对分子质量为43.30 kDa,理论pI 为5.15,脂肪系数为84.50,该蛋白有159 个疏水性氨基酸,230 个亲水性氨基酸,84 个酸性氨基酸,51 个碱性氨基酸(图4)。
2.3 OsECH1 蛋白的亲疏水性分析
借助软件ProtScale 分析OsECH1 蛋白的亲/疏水性,结果显示,最大值为–2.75,最小值为2.25,总平均亲水性(GRAVY)为–0.316,表明OsECH1 蛋白为亲水性肽链(图5)。
2.4 OsECH1 蛋白的磷酸化位点分析
OsECH1 蛋白含有丝氨酸(Ser)、苏氨酸(Thr)和酪氨酸(Tyr)磷酸化位点数分别为26 个、14个和3 个(图6)。
2.5 OsECH1 蛋白的亚细胞定位
蛋白质的亚细胞定位是阐明其相互作用的蛋白质、功能及其在细胞中的潜在作用的重要步骤[10]。本研究将构建好的1302-OsECH1 质粒转入水稻原生质体细胞中, 荧光信号结果显示OsECH1 蛋白定位于细胞核中(图7)。
2.6 OsECH1蛋白与5个防御相关蛋白互作分析
将重组载体pET28a-OsECH1 成功转入BL21(pTf16)中进行扩大培养后,用IPTG 大量诱导表达。利用亲和镍层析柱进行蛋白纯化,分别用浓度从低至高20、60、80、100、200、500 mmol/L咪唑进行洗脱。以纯化的OsECH1 蛋白作为诱饵,以水稻根总蛋白作为猎物,进行His-tag pull down分析。结果发现5 个蛋白与OsECH1 蛋白相互作用(表2),包括vATP-synt_E 家族蛋白(OsVE,Os01g0659200)、Ricin-B 家族蛋白(OsR40C1,Os03g0327600;OsR40G2,Os07g0683900)、过氧化氢酶(OsCATB,Os06g0727200)、B_lectin(OsBAT1, Os09g0111950)。
2.7 OsECH1 基因调控5 个互作蛋白的表达
为了更好地了解OsECH1 基因对其互作蛋白的调控,本研究构建了OsECH1 基因的过表达载体及RNAi 载体,并获得过表达(OE-OsECH1)水稻和基因沉默(RNAi-OsECH1)水稻。为了确保已获得转基因水稻的阳性植株,在转基因水稻中观察OsECH1 的表达水平变化。与野生型‘日本晴(WT)相比,该基因的表达量在RNAi-ECH水稻中显著减少,在OE-ECH 水稻中显著增强。表明已成功获得了OsECH1 基因的过表达和基因沉默转基因水稻(图8A)。
为了研究OsECH1 基因与5 个蛋白之间的关系,分别分析其在3 种水稻中的表达量变化情况。RT-qPCR 结果显示,OsVE、OsCATB、OsR40G2基因的表达量在RNAi-OsECH1 水稻中显著上调,OsBAT1 基因的表达量在OE-OsECH1 水稻中显著上调, OsR40C1 基因在RNAi-OsECH1 、OEOsECH1水稻中均显著上调(图8B)。以上结果表明,OsECH1 基因对这5 个蛋白具有调控作用,为后续探究OsECH1 基因的功能提供新思路。
2.8 OsECH1 蛋白影响细尖潜根线虫的侵染
为了验证转录组分析结果,比较OsECH1 基因在健康的野生型‘日本晴(WT)与受侵染的野生型‘日本晴中的表达水平(图9A)。OsECH1基因在受侵染的WT 水稻根部中表达较高(12%~15%)。通过比较3 种水稻中细尖潜根线虫基因的表达量变化,发现与WT 水稻相比,在RNAi-OsECH1 水稻中细尖潜根线虫的侵染量增加,但在OE-OsECH1 水稻中细尖潜根线虫的侵染量减少(图9B)。结果再次表明OsECH1 蛋白可能在植物与细尖潜根线虫的互作中起作用。
3 讨论
目前,水稻病害日益严重,尤其是植物寄生线虫病。随着水稻中植物寄生线虫抗性基因的分离和克隆技术的快速发展,极大地促进了植物与线虫之间互作的研究,更有利于防治植物线虫病。虽然关于线虫效应子的研究越来越多,但关于水稻潜根线虫的研究较少。通过研究线虫效应子寻找水稻抗病基因一直是一个研究热点。廖金铃和卓侃团队发现了根结线虫效应子MgMO289 靶向水稻铜金属伴侣OsHPPO4 抑制水稻的方位反应,以影响植物免疫并促进侵染,这是首次发现植物病原物利用植物超氧阴离子降解系统清除活性氧,从而抑制植物免疫反应[11]。虽然在寄主植物中已经成功克隆了一些抗线虫基因,但其抗性机制尚不清楚,并且寄主植物与线虫的相互作用模型尚未建立。因此,线虫效应子与抗性基因蛋白相互作用的研究将有助于进一步研究寄主植物-寄生线虫相互作用的机制。
前期研究表明,烯酰辅酶A 水合酶(ECH)属于水合酶/异构酶家族,并且ECH 在不同物种中序列同源性非常高[12-13]。ECH 是脂肪酸β 氧化过程中不可缺少的酶,ECH 的异常代谢会导致脂肪酸代谢紊乱[14]。因此,酰辅酶A 水合酶在生命活动中有着非常重要的作用。目前,ECH 的功能在哺乳动物中被广泛研究,而在植物中的研究较少。已有研究证明水稻中的ECH 与MAP1 相互作用,MAP 蛋白与稻瘟病的防治有关,从而证明ECH 可能为稻瘟病的防治提供新的方向[15]。编码烯脂酰辅酶A 合成酶的ACTT5、编码烯脂酰辅酶A 水合酶ACTT6 是葡萄孢白僵菌柑橘致病型中活性毒素生物合成的必需基因[16]。
本研究在日本晴根部cDNA 中克隆出OsECH1基因并获得其纯化蛋白。通过His-tag pull down实验发现5 种蛋白与OsECH1 互作,包括vATPsynt_E 家族蛋白、Ricin-B 家族蛋白、过氧化氢酶(CAT)和B_lectin 家族蛋白,而这些家族均已经被证明与防御相关。
OsR40C 家族于1997 年由MOONS 等[17]首次报道,OsR40C1 和OsR40G3 的表达水平在盐处理的水稻根中显著上调。CAT 在植物生长发育、胁迫防御反应、氧化衰老等生理过程中发挥着重要作用。研究表明,脱落酸(ABA)通过诱导水分胁迫下水稻叶片中的OsCATB 控制H2O2 积累[18]。低温、干旱、氧化应激、水杨酸(SA)和ABA可诱导拟南芥中CAT1、CAT2 和CAT3 基因的表达和CAT 酶活性[19]。V-ATPase 酶对植物生长和发育至关重要。研究发现细胞中的V-ATPase 酶参与植物对寒冷、盐分和重金属胁迫的反应。高盐和低温胁迫可以诱导V-ATPase 酶的结构和数量发生变化,从而提高个体植物对环境变化的适应性,减少逆境对个体植物的伤害[20]。拟南芥的V-ATPase 酶的a、e 和g 亞基也可被高盐胁迫诱导,这表明 V-ATPase 酶在应激反应中存在潜在功能[21]。植物凝集素广泛分布于动物、植物和微生物中,现在已发现1000 多种植物凝集素,它已被公认是高表达蛋白,在水稻中已经发现了1100个基因[22]。植物凝集素可以特异性结合糖类,在植物防御反应中发挥重要作用[23]。Pi-d2 是水稻中的一种G 型凝集素受体激酶,主要功能是参与抗稻瘟病[24]。OsLecRK 不仅参与调控水稻种子萌发,而且在抗病性方面也发挥着重要作用。敲除该基因可抑制抗病基因的表达,从而降低水稻对真菌和细菌病原菌及害虫的抗性[25]。这些结果为进一步研究OsECH1 的功能提供了新思路。通过分析OsECH1 基因的功能,可增加对ECH 家族功能的理解,进一步分析OsECH1 蛋白及互作蛋白的功能,有助于探究抗潜根线虫侵染的作用机理。
