野生种西印度瓜与甜瓜种间杂种的创制
2023-05-29郑莉娜毕研胜孙亚亭王克征钱春桃李季程春燕娄群峰
郑莉娜,毕研胜,孙亚亭,王克征,钱春桃,李季,程春燕,娄群峰
(南京农业大学园艺学院/作物遗传与种质创新利用全国重点实验室,江苏 南京 210095)
甜瓜(CucumismeloL.)属于葫芦科(Cucurbiteace )甜瓜属(Cucumis)作物,为一年生蔓性草本植物。我国的甜瓜种植面积占世界总面积的45%以上,产量占55%以上。甜瓜营养丰富,色、香、味俱佳,深受消费者们的青睐[1-4]。但是由于甜瓜的遗传基础相对狭窄,其抗病虫害和抗逆性较差,严重限制了甜瓜产量和品质的提高。
甜瓜属起源于非洲东部,除甜瓜种外,还包含6系29个甜瓜近缘种[5]。甜瓜近缘野生种与甜瓜之间的遗传差异性,为甜瓜属作物的种间改良提供了宝贵的资源。例如非洲角黄瓜(C.metuliferus)对南方根结线虫和病毒病具有抗性;西印度瓜(C.anguria)对枯萎病、病毒病和南方根结线虫病均具有抗性[6]。在甜瓜属中,野生种之间的杂交比较容易成功,例如C.anguria、C.dipsaceus、C.africanus、C.leptodermis和C.myriocarpus等之间的杂交[7],而野生种与栽培种之间仅有C.hystrix与栽培黄瓜杂交成功[8],栽培甜瓜与野生种之间成功杂交还未见报道。
西印度瓜为甜瓜属的一个野生种,国内外众多学者对其与同属植物之间的种间杂交进行了大量研究。例如,Matsumoto等[9]得到了西印度瓜与甜瓜杂交的果实,但未有成熟的种子;Fassuliotis 等[10]通过胚培养和体细胞杂交获得了C.metuliferus×C.anguria的种间杂交F1;王吉明等[11]开展了系列甜瓜与西印度瓜的种间杂交,在甜瓜V129×西印度瓜组合中获得了杂交F1种子,并在F1植株中检测出西印度瓜的少量SSR条带,但未观察到西印度瓜的性状;王吉明等[12]对西印度瓜与V007进行种间杂交,在授粉3 d后的幼胚DNA中扩增出父本的特征带,表明西印度瓜与甜瓜产生了真实的杂交。但未见F1植株特性的报道,更未见进一步的外源性状(遗传物质)回交转移的报道。
因此,本研究以野生种西印度瓜和栽培种甜瓜为试验材料,进行种间杂交试验,通过对获得的杂交后代F1、BC1及亲本植株进行形态学观察、细胞学观察和分子标记分析,验证获得的F1为种间杂种,并通过其与甜瓜回交将西印度瓜的外源遗传物质导入到栽培甜瓜中,为深入开展甜瓜种间优异基因的转移提供重要参考依据。
1 材料与方法
1.1 试验材料
试验共选用13份种质材料,其中普通甜瓜栽培种12份和野生种材料西印度瓜1份(表1)。杂交试验于2020至2021年春季和秋季,在南京农业大学白马科研教学基地试验场地进行。
1.2 试验方法
1.2.1 种间杂交以甜瓜栽培种与野生种作为双亲,共配制正反种间杂交组合24个,以母本自交作为对照。有性杂交处理:在开花前一天,选取生长健壮,状态良好的母本植株雌花去雄,用镊子轻轻剖开花萼及花冠,然后用镊子夹住花丝,将雄蕊全部去掉,并进行套袋隔离和挂牌标记。
在开花当天上午09:00至10:00,花粉活力最好时使用2~3朵雄花对母本雌花进行第1次授粉,次日和第3天早上分别进行第2次和第3次授粉,共计授粉3次[13]。授粉后,对雌花子房喷施浓度为0.01~0.02 g·L-1的坐果灵2~3次,再使用0.01~0.02 mg·L-1TDZ激素喷施2~3次。
1.2.2 胚拯救借鉴Chen等[8]的胚拯救经验,进行以下操作:取杂交授粉后14~21 d的幼嫩果实,清洗果实表面后,再用75%(体积分数)乙醇对表面消毒,然后在超净工作台中用解剖刀将幼胚剥出;然后用滤纸吸干多余的水分,将幼胚放在添加1 mg·L-16-BA的MS培养基中,然后置于26~28 ℃的培养室内,在光周期16 h/8 h的条件下培养,直接诱导幼胚分化成苗。
当幼苗生长至4~5片真叶,植株顶端即将到达培养瓶的瓶口时,将培养瓶的瓶口打开12 h进行炼苗,使其适应外界的环境,然后将植株从培养基中取出,清洗干净根部的培养基,移栽到装有基质的盆钵中进行育苗。移栽的第1周置于温度为25~28 ℃,光照度为1 400~1 600 lx,湿度为61%~81% 的光照培养箱中进行移栽驯化培养。驯化1周左右,将植株移栽到田间进行栽培管理。
1.2.3 植物学形态特征鉴定2021年3月初,将父本P1、母本P2和杂交所得的F1播种于南京农业大学白马科研教学基地,3月中旬进行定植。随机选取双亲及其杂种F1进行形态学观察,主要从植株的株型、花形、叶形以及果实性状等方面进行植株的形态鉴定。随机选取9个植株,测量株高作为整株形态指标;从每个植株上随机选取9个花序,测量统计花径、花瓣数量、花瓣颜色和形状;从顶芽以下同一节位区间的叶片作为对象,每一棵植株随机选取9张叶片测量叶片的长度和宽度,作为叶形态指标;从每个植株中随机选取9个果实测量果实的纵横径等作为果实性状指标。所得性状数据使用SPSS Statistics 25软件进行单因素方差分析。

表1 供试种质材料Table 1 Tested germplasm materials
根据Peterson等[14]的方法,在盛花时期,晴朗天气随机选取亲本及F1植株即将开放的2朵花;将花粉涂抹在载玻片上,并用醋酸洋红染色,盖上盖玻片,滤纸吸取多余染液;在显微镜10×镜下观察每朵花的3个视野,统计可染花粉粒占视野花粉粒总数的比例即花粉可染率。
1.2.4 杂交F1染色体数目鉴定在胡静轩等[15]方法的基础上,对杂交植株进行继代扩繁时,按单株取培养瓶中F1的健康根尖1.0~2.0 cm,并迅速置于8-羟基喹啉溶液中预处理1 h,然后转入M∶G卡诺固定液(甲醇与冰醋酸的体积比为3∶1)中固定24 h以上,用于根尖染色体数目鉴定。染色体制片采用酶解去壁低渗火焰法,经DAPI(4′,6-二脒基-2-苯基吲哚)荧光染料染色后,使用荧光显微镜(Image Z2)在100×镜下观察并拍照。
1.2.5 分子标记分析DNA的提取:采用改良CTAB法[16]提取父母本以及F1新鲜幼嫩叶片基因组DNA,用核酸浓度检测仪检测DNA浓度,并将DNA的质量浓度稀释至50~100 ng·μL-1,置于-20 ℃保存备用。
SSR标记分析:选用多态性好、条带清晰稳定的54对SSR引物,对亲本P1、P2和F1进行分子标记分析。PCR总反应体系为10 μL,其中正、反引物各1 μL,DNA1 μL,Mix 5 μL,ddH2O 2 μL。PCR反应扩增程序为:95 ℃ 5 min;95 ℃ 30 s,50 ℃ 30 s,72 ℃ 40 s,28个循环;72 ℃ 10 min;4 ℃保存。PCR产物采用聚丙烯酰胺凝胶电泳以及银染法进行检测。
2 结果与分析
2.1 种间杂交F1的获得
栽培甜瓜以及野生种均能正常开花结果,进行人工杂交授粉的24个杂交组合中,只有西印度瓜ב新红瑞’杂交组合获得果实,且果实中含有半饱满状态的种胚;还有13个杂交组合也得到了果实,但没有类似的种胚;其余的杂交组合均没有得到果实(表2)。
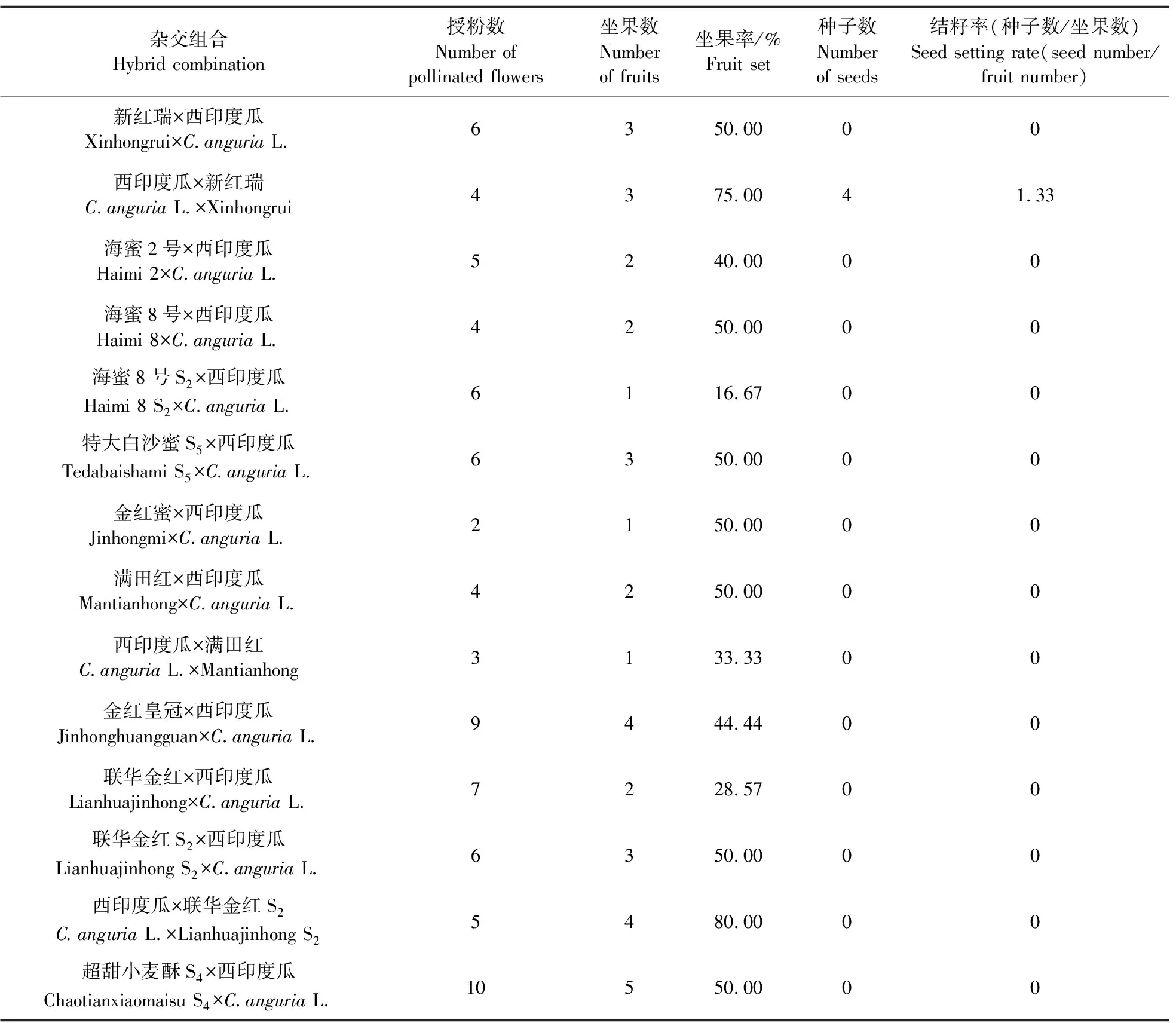
表2 不同类型杂交组合坐果收种情况Table 2 Information on fruit set and seeds collection of different hybrid combinations
采用胚拯救技术对授粉后14~21 d的4粒杂交幼嫩胚胎进行组织培养,结果(图1-A)显示,其中1粒幼胚在 6 d后开始膨大萌动;15 d后幼苗子叶出现颜色淡黄现象,26 d后发育成完整植株。此外,还有1粒幼胚虽发育时间推迟(约1个月),但亦发育成植株。利用幼胚拯救技术,共有3粒幼胚萌发,出苗率为75.00%。将第1粒培养出的再生植株进行扩繁,获得了2株扩繁苗,其他的每粒各获得1株再生植株。经过驯化移栽,最后成功保存下4株杂交F1代植株。
2.2 F1植物学形态特征观察
为证实杂交F1代植株是否为真正的种间杂种,在植物生长期间对种间杂种F1代植株及其双亲进行田间性状观察统计。由表3可见:双亲与F1在叶片宽度这一性状上均差异显著,F1的节间长度与母本P2西印度瓜差异显著,而与父本P1无显著性差异,其余性状F1均与母本无显著性差异,与父本存在显著性差异。种间杂种F1植株在幼苗期,其顶部的叶片有明显的黄化现象,但父母本均无此现象(图1-B);对比双亲与杂种F1茎基部的幼叶发现,F1与甜瓜的叶片尖端形状均为钝状,明显不同于母本的叶端尖锐(图1-C),但整个叶片形状与母本相似。杂交后代F1的雄花花瓣大小和形状都介于双亲之间,属于中间型,且F1的雄花与父本雄花一样具有花晕(图2)。但是,在植株成熟期,F1的叶片颜色、形状和雄花等性状都趋于与母本西印度瓜的表现型一致,F1果实的果梗,均短于母本西印度瓜的果梗。另外,西印度瓜茎部的毛为绒毛,栽培甜瓜茎部的毛为毛刺,F1的茎部既有软绒毛又有硬的毛刺,这一性状居于双亲中间(图3)。

图1 F1组培幼苗观察及其与栽培甜瓜P1(父本)和野生种P2(母本)的形态学比较Fig.1 Observation of F1 plantlets in tissue culture and its morphological comparison with cultivatedmelon P1(male parent)and wild species P2(female parent)A. 杂种F1的组织培养(红色箭头表示萌发的种胚);B. 幼苗;C. 幼苗期叶片。A. Tissue culture of hybrid F1(The germinated embryo is indicated by red arrow);B. Plant;C. Leaves at seedling stage.
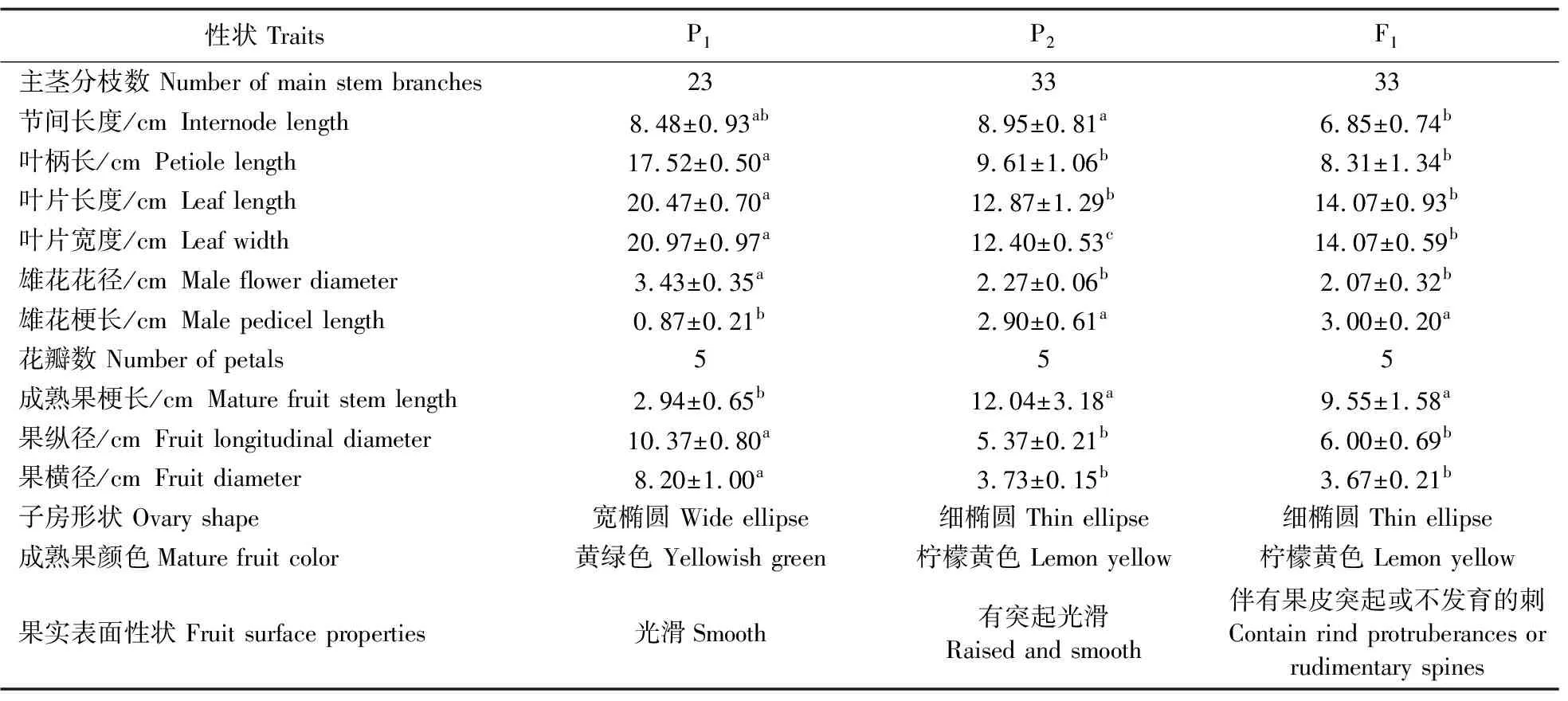
表3 双亲及杂种F1的农艺性状Table 3 Agronomic characters of parents and hybrid F1

图2 幼苗植株前期的雄花比较Fig.2 Comparison of male flowers in the early stageA. 栽培甜瓜‘新红瑞’P1;B. 杂种 F1;C. 西印度瓜P2。A. Cultivated melon‘Xinhongrui’P1;B. Hybrid F1;C. C.anguria P2.

图3 栽培甜瓜P1、杂种F1以及野生种P2植株茎部绒毛比较Fig.3 Comparison of stem villi of cultivated melon P1,hybrid F1and wild species P2

图4 F1根尖有丝分裂中期制片Fig.4 Apical mitotic metaphasesection of F1
2.3 杂交F1的染色体数目鉴定
对西印度瓜ב新红瑞’杂交获得的4株F1子代,进行染色体数目观察,经鉴定杂种后代F1的染色体数目为24条(图4)。说明染色体数相同的甜瓜属植物(2n=2x=24),在杂交后代F1的形成过程中染色体数量不变。因此还需要对杂交后代进行进一步的分子标记鉴定。
2.4 种间杂交F1的SSR分析
参考钟洋敏等[17]的方法,从54对SSR引物中筛选出52对具有多态性的引物,对父母本及杂种F1进行扩增,其中8对引物即3-4、4-1、5-1、5-2、8-1、8-2、8-5、11-1(表4),在F1杂种中均扩增有父母本的共显性标记,分别表现为父母本的特征带,扩增条带表现为双亲互补型,即在分子水平上证明了野生种西印度瓜与栽培甜瓜杂交所获得的种间杂种的真实性(表2,图5)。

图5 SSR分子标记杂种鉴定结果Fig.5 The result of SSR molecular analysis for hybrid identificationⅠ—Ⅵ分别代表F1与双亲扩增产物的6种不同类型;箭头所指代表SSR特异性识别位点。Ⅰ-Ⅵ represent six different types of F1 and parental amplification products respectively;Black arrows indicate the SSR specific recognition site.

表4 杂种鉴定SSR引物序列Table 4 Sequences of SSR primers for hybrid identification
54对SSR扩增产物的分析表明,P1、P2及其F1代扩增产物可分为以下6种类型(图5):(Ⅰ)P1、P2和F1存在有杂合带,它反映了F1与双亲DNA的同源关系(表4中的8对引物);(Ⅱ)F1与双亲之一存在共有带,且皆为F1与P1的共有带(引物2-1、3-1、6-5);(Ⅲ)仅一个亲本有带、另一亲本和F1无带,且皆为P1的独有带(引物2-5、6-1、7-1、10-1、10-3);(Ⅳ)双亲皆没有带,仅F1有带(引物1-3、2-3、2-5);(Ⅴ)双亲有带而F1无带(引物2-4);(Ⅵ)其余的引物均为双亲和F1都有带,但是F1的带型与母本一样。

图6 以甜瓜为母本的BC1植株SSR鉴定Fig.6 SSR identification of BC1 plantswith melon as female parent
2.5 回交植株BC1的特征及SSR分析
以栽培甜瓜‘新红瑞’为轮回亲本与种间杂种F1进行回交,获得2个回交后代BC1株系。通过对母本‘新红瑞’、F1和回交后代BC1进行田间表型观察,发现回交植株后代的卷须有苦味,但双亲的卷须均无苦味,除该性状外,其他性状与栽培甜瓜基本一致。初步推断苦味性状的出现,可能与种间杂交回交过程中双亲遗传物质发生了重组交换,导致新的基因突变有关。使用54对SSR引物对回交后代植株进行分子生物学鉴定,引物5-5(F:TCCGAAGTAAACATCAAAGACA;R:GGTCAGTCAAGATAGTTACGGTTG)的扩增结果显示具有杂合互补条带,表明BC1中同时具有母本甜瓜‘新红瑞’和F1的特征条带(图6),也从侧面表明BC1中含有少量西印度瓜的遗传片段。
3 讨论
远缘杂交是拓宽作物遗传基础,扩大基因库的有效途径之一。Li等[18]通过远缘杂交将野生小麦具有的强适应型和抗条锈病特性基因成功导入栽培小麦中;Chen等[19]利用远缘杂交成功开发新的耐盐水稻Spartina种质;陈发棣等[20]将野生种质资源特有的优良性状通过远缘杂交进行遗传重组,培育成系列高抗(耐)性菊花新品种,在抗蚜虫性、花期及株型等育种方面取得了突破性进展。本研究将抗病抗逆性强的野生种西印度瓜与栽培种甜瓜‘新红瑞’进行种间杂交,获得种间杂种后代F1,为进一步实现西印度瓜有益性状的应用,尤其是为抗南方根结线虫这一性状向栽培种甜瓜中转移提供了可能,目前为止,在栽培甜瓜中还未发现抗根结线虫的种质资源[21]。
一般在进行杂交时,正向和反向杂交所获得的F1有明显差别,例如正向杂交的结实率和出胚率都较高,但反向杂交的结实率却较低,这种单向杂交不亲和性在多种作物远缘杂交中常有报道[22-25]。本课题组前期进行黄瓜(C.sativus)和酸黄瓜(C.hystrix)正反杂交时,首先在野生种为母本的反向C.hystrix×C.sativus杂交中取得成功[26]。本研究在以西印度瓜为母本的杂交组合中,收到含有成熟种子的果实,反向以甜瓜为母本仅收到果实,这与柳李旺等[27]的报道类似。
种间杂交过程中内源激素变化异常,会导致杂种胚胎发育异常,使得胚胎在心形胚阶段就停止发育或直接败育[28]。此时及时外施生长激素能够明显促进杂种胚珠生长,促进胚乳发育,为胚胎的分化与发育提供必需的生理条件[29-30]。本研究进行甜瓜种间杂交时,对子房喷施了适量的外源激素(TDZ),促进坐果,并获得少量较饱满的种子(胚),可能喷施外源激素可以克服内源激素异常带来的不利影响,从而促进胚胎的发育。但是绝大多数的组合仅收到不含种子的果实,因此对于外施激素的精准使用(如激素种类、浓度、处理时间)以及外源激素是如何促进杂种胚胎发育的机制还有待深入研究。
研究表明,当杂交亲本之一为野生种时,因野生性状多为显性,F1植物表现野生种性状较多[27,31],不再表现为中间型。本研究的F1在幼苗生长前期,叶片和雄花的形状大小等含有父本特征,表现偏向于中间型;但在F1生长中后期,除果梗长短和植株茎部绒毛外,其余性状均与野生种母本西印度瓜保持一致,不再表现为中间型。这些性状表现出一种动态消失的变化过程,可能是栽培种甜瓜(父本)的性状表达逐渐受到野生种西印度瓜遗传背景的作用而导致的。也有人认为在植物远缘杂交过程中,存在着染色体消除的现象,即亲本一方的染色体在杂种合子或幼胚发育初期被有选择消除[32-34],从而导致该亲本性状在杂种后代中未能出现或出现之后又消失的现象。当远缘杂交后代性状受父母本影响的程度不对等时,可以在种间杂交的基础上进行多次迭代杂交,也许能够促进更多的父本遗传物质渗入到杂交后代中[35]。陈劲枫等[36]的研究表明,当杂种作为供体亲本与栽培黄瓜进行回交时,抗性又被进一步转移到回交一代中。因此,我们下一步将开展多次迭代杂交,以促进西印度瓜的遗传物质向栽培甜瓜中转移。
本试验SSR的鉴定结果表明,F1同时具有母本西印度瓜和父本甜瓜的特征带和F1子代中仅有父本甜瓜的特征带,这2种情况都表明远缘杂交的幼胚中含有父本甜瓜的遗传物质。双亲均无条带,但F1仍可扩增出条带,以及F1中扩增出不同于双亲的条带,这两种情况表明父母本之间可能发生了遗传重组,从而产生了新条带。另外,在回交子代BC1中扩增出双亲杂合带的是一对新的引物,不同于在F1扩增出杂合带的8对引物,再次表明F1遗传物质已经发生重组,产生了新的遗传组合,但关于其具体机制还需要深入的研究和求证。
