干旱胁迫下喷施黄腐酸对燕麦光合及其抗氧化酶活性的影响
2023-01-04朱珊珊米俊珍赵宝平武俊英刘景辉
朱珊珊,米俊珍,赵宝平,武俊英,王 莹,刘景辉
(内蒙古农业大学 农学院,呼和浩特 010019)
燕麦(AvenasativaL.)作为中国北方重要的粮饲兼用作物,具有丰富的营养价值和保健功能,多种植于干旱、半干旱地区,干旱是制约燕麦产量最主要的环境因子[1]。随着气候的变暖,极端干旱天气日益频发,植物干旱缺水的情况更加严重[2]。干旱胁迫使植物体内活性氧产生和清除系统之间的平衡遭到破坏,导致活性氧过量积累,造成植物细胞膜结构损伤[3],还导致植物叶片光合色素的积累减少,光合作用受到抑制,细胞正常代谢受阻,最终影响植物生长[4-5]。因此,探究改善燕麦对干旱胁迫适应能力的有效农艺措施及其调节机制具有重要的理论和实践意义。
黄腐酸是腐植酸中具有亲水性、分子量较小的有机化合物,属于水溶性腐植酸,对植株生长起促进作用[6]。黄腐酸可以增加植株干物质积累量,提高光合色素含量,增强植株对逆境的抵抗能力[7]。有研究表明,干旱胁迫下,腐植酸能改善作物叶绿素荧光参数,提高植株气体交换,进而提高作物产量及抗旱能力[8-9]。Zhang等的研究表明,干旱胁迫下喷施黄腐酸能更好地提高苹果植株的抗氧化酶活性,增加脯氨酸含量,降低丙二醛含量使其新陈代谢旺盛,增强植物的抗旱能力[10]。尽管黄腐酸在燕麦抗旱性方面已经有一些研究[8,11-12],但是关于干旱胁迫下燕麦光合特性和抗氧化酶活性相结合对黄腐酸的响应鲜见报道。因此,本研究以燕麦为试验材料,通过分析干旱胁迫下黄腐酸对燕麦光合特性及抗氧化酶活性的影响,为黄腐酸在燕麦抗旱种植中的应用提供理论指导和技术支撑。
1 材料和方法
1.1 试验材料及设计
试验于2021年4月至8月在内蒙古农业大学燕麦产业研究中心温室进行。试验材料为‘燕科2号’,由内蒙古农牧业科学院提供。黄腐酸采购于范德生物科技公司,纯度大于85%。试验选用直径为25 cm,高18 cm的塑料盆,内装混合土5 kg(河沙∶泥炭土体积比1∶1),泥炭土中N+P+K大于10 g·kg-1,有机质量大于50 g·kg-1,pH值为5.5~8.5,土壤含水量为9%。播种前以2 g/盆磷酸二铵做底肥施入(N 18%,P2O546%)。每盆播种30株,三叶期定植为20株。采用随机区组排列,随机分成7组做如下处理:对照组(CK),正常供水(田间持水量的75%)+叶面喷施蒸馏水;单独干旱胁迫处理(D0),干旱胁迫(田间持水量的45%)+叶面喷施蒸馏水;干旱胁迫下叶面喷施黄腐酸处理,干旱胁迫(田间持水量的45%)+叶喷不同浓度的黄腐酸处理(D1~D5),各处理黄腐酸浓度依次为200、400、600、800、1 000 mg/L。每个处理重复3次,每次重复种植3盆,共63盆。从2021年5月10日拔节期开始采用称重差值法进行控水[13],在控水开始后的上午9:00-11:00进行喷施黄腐酸和清水,连续喷施3 d,每盆喷施量为45 mL,喷施处理结束后第7天进行各项指标测定。
1.2 测定项目与方法
1.2.1 地上部鲜、干重每个重复取5株燕麦幼苗,放入纸袋称鲜重,每个处理重复3次。测完鲜重的样品放入烘箱105 ℃杀青30 min,80 ℃烘干到恒重,称取干重。
1.2.2 光合特性的测定(1)光合气体交换参数:在晴朗无风的上午9:00-11:30,用便携式光合仪(CIRAS-3)测定旗叶气体交换参数,包括净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)。每个处理各测3株。测定时饱和光强为1 200 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1,叶室温度为30 ℃。
(2)光合色素含量:采用乙醇浸提法[14]。取燕麦旗叶,用蒸馏水洗净、擦干后剪碎混匀,取0.20 g放入25 mL容量瓶中,用95%乙醇定容后置于黑暗处浸提,直至叶片完全变白,最后以95%乙醇为对照,测定其在470 nm、649 nm和665 nm处的吸光值,计算叶绿素a、叶绿素b和类胡萝卜素含量,每个处理重复3次。
(3)叶绿素荧光参数:每个处理选择3片生长完好的旗叶叶片,用FMS-2便携式脉冲调制式荧光仪测定叶绿素荧光参数。先测定光下最大荧光(Fm′)、稳态荧光(Fs)和光下最小荧光(Fo′),然后将同部位燕麦幼苗叶片暗适应20 min,测定初始荧光(Fo)、最大荧光(Fm),通过以上测定的叶绿素荧光参数计算出光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)=(Fm-Fo)/Fm,光系统Ⅱ实际光化学效率(ΦPSⅡ)= (Fm′-Fs)/Fm′,光化学猝灭系数(qP)=(Fm′-Fs)/(Fm′-Fo′)和非光化学猝灭系数(NPQ)=(Fm-Fm′)/Fm′,电子传递效率(ETR)=ΦPSⅡ*PAR*0.84*0.5,PAR为光合有效辐射。
1.2.3 抗氧化酶活性测定取各处理的燕麦旗叶,剪成小段混匀,用来测定以下生理指标。分别称取0.5 g,采用氮蓝四唑(NBT)光还原法测定超氧化物歧化酶(SOD)活性[15],采用愈创木酚法测定过氧化物酶(POD)活性[16];称取0.1 g,采用过氧化氢法测定过氧化氢酶(CAT)活性[17]。以上生理指标每个处理3次重复。
1.3 数据统计分析
采用Microsoft Excel2016进行数据处理及作图,使用SPSS22.0软件进行单因素方差分析,并运用Duncan’s检验法对显著性差异进行多重比较(P<0.05)。
2 结果与分析
2.1 喷施黄腐酸对干旱胁迫下燕麦地上部鲜、干重的影响
图1显示,燕麦的地上部鲜重和干重在单独干旱胁迫处理(D0)下较CK分别显著降低了25.39%和37.84%(P<0.05);与D0处理相比,干旱胁迫下喷施黄腐酸处理(D1~D5)中,D3处理燕麦幼苗的地上鲜重和干重显著提高(P<0.05),其余处理无显著变化,其中D3处理的地上鲜重和干重比单独干旱胁迫处理(D0)显著升高了28.59%和39.13%(P<0.05)。说明叶面喷施适宜浓度的黄腐酸可显著促进干旱胁迫下燕麦幼苗地上生长,并以600 mg/L黄腐酸处理效果最好。
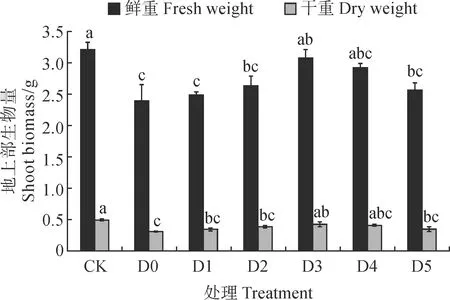
CK. 对照,正常供水(田间持水量的75%);D0. 干旱胁迫(田间持水量的45%);D1~D5. D0 +叶面分别喷施200、400、600、800、1 000 mg/L的黄腐酸处理;不同小写字母表示同一指标不同处理间差异显著(P<0.05),下同图1 干旱胁迫下黄腐酸对燕麦地上部鲜重和干重的影响CK. Control, normal water supply (75% of field capacity); D0. Drought stress (45% of field capacity); D1-D5. D0 and foliar sprayed with 200, 400, 600, 800 and 1 000 mg/L fulvic acid, respectively; Different lowercase letters represent significant differences among different treatments of the same index (P<0.05); the same as belowFig.1 Effects of fulvic acid on dry and fresh weight of A. sativa shoot under drought stress
2.2 喷施黄腐酸对干旱胁迫下燕麦叶片光合气体交换参数的影响
由表1可知,干旱胁迫处理下(D0),燕麦幼苗的Pn、Tr和Gs较CK均显著下降,分别降低了54.45%、26.88%和57.69%,而Ci则提高了35.59%,差异均达显著水平(P<0.05)。与D0相比,叶面喷施不同浓度黄腐酸对燕麦的Pn、Tr和Gs均具有一定促进作用,但随着黄腐酸处理浓度的增加呈现先升高后降低的趋势,并均在D3处理达到最大值,增幅分别为74.38%、26.47%和43.34%,而Ci则表现出完全相反的规律,并在D3处理达到最小值,降幅为19.52%,差异均达显著水平(P<0.05)。可见,适宜浓度的黄腐酸处理能有效提高干旱胁迫下燕麦幼苗的光合速率,增强光合能力。

表1 干旱胁迫下黄腐酸对燕麦叶片光合气体交换参数的影响
2.3 喷施黄腐酸对干旱胁迫下燕麦叶片光合色素含量的影响
由表2可知,干旱胁迫处理下(D0), 燕麦叶片的叶绿素a、叶绿素b和类胡萝卜素及叶绿素a+b含量较CK均显著下降(P<0.05),分别降低了28.14%、14.95%和42.42%及23.86%。与D0相比,叶面喷施不同浓度黄腐酸对燕麦叶片的叶绿素a、叶绿素b和类胡萝卜素及叶绿素a+b含量均具有促进作用,但随着黄腐酸处理浓度的增加呈现先升高后降低的趋势,并均在D3处理达到最大值。与D0相比,D3处理的叶绿素a、类胡萝卜素和叶绿素a+b含量,分别增加了25.17%、47.37%和21.03%,差异均达显著水平(P<0.05),叶绿素b含量增加了13.19%,差异不显著。

表2 干旱胁迫下喷施黄腐酸对燕麦叶片光合色素含量的影响
2.4 喷施黄腐酸对干旱胁迫下燕麦叶片叶绿素荧光参数的影响
表3显示,燕麦幼苗叶片Fv/Fm、ΦPSⅡ、qP和ETR在单独干旱胁迫处理(D0)下均较CK明显降低,而Fo、NPQ升高,且均达到显著差异水平(P<0.05)。在干旱胁迫条件下,叶面喷黄腐酸处理(D1~D5)幼苗叶片的Fv/Fm、ΦPSⅡ、qP和ETR均随着黄腐酸浓度的增加而先升高后降低,并均在D3处理时达到最大值,而其叶片Fo、NPQ则随着黄腐酸的增加而先降低后升高,并在D3处理时达到最小值;与D0相比,幼苗叶片的Fv/Fm、ΦPSⅡ、qP和ETR在D1~D5处理均不同程度升高,其中Fv/Fm、ΦPSⅡ和ETR在D3达到显著水平(P<0.05),增幅分别为6.49%、69.57%和70.71%,qP未达到显著水平;其叶片Fo、NPQ在D1~D5处理下均不同程度降低,其中D2~D5处理的降幅均达到显著水平(P<0.05),最低的D3处理降幅分别为13.32%、43.75%。
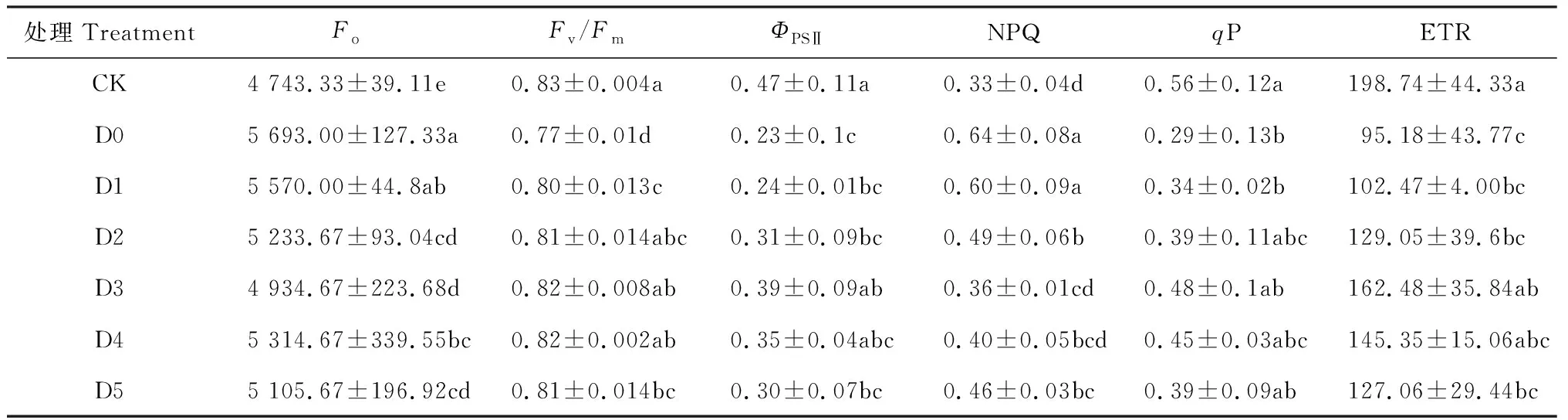
表3 干旱胁迫下喷施黄腐酸对燕麦叶片叶绿素荧光参数的影响
2.5 喷施黄腐酸对干旱胁迫下燕麦叶片抗氧化酶的影响
图2显示,燕麦叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性在单独干旱胁迫处理(D0)下均比CK明显增加,增幅分别为25.68%、19.98%和7.29%,且SOD和POD达到显著差异水平(P<0.05)。在单独干旱胁迫下,叶喷黄腐酸处理(D1~D5)叶片的SOD、POD、CAT活性均随着黄腐酸浓度的增加而先升高后降低,并均在D3处理时达到最大值。与D0相比,幼苗叶片的POD、CAT活性在D1~D5处理均不同程度升高,且均达到显著水平(P<0.05),其中在D3处理下增幅最大,分别为76.57%、55.26%。其叶片SOD活性在D1~D5处理下均不同程度升高,其中的D3处理增幅达到显著水平(P<0.05),增幅为12.19%。说明黄腐酸能显著提高干旱胁迫下燕麦幼苗的SOD、POD、CAT活性,增强燕麦幼苗叶片细胞对活性氧的清除能力,缓解因干旱胁迫而引起的膜脂过氧化程度。
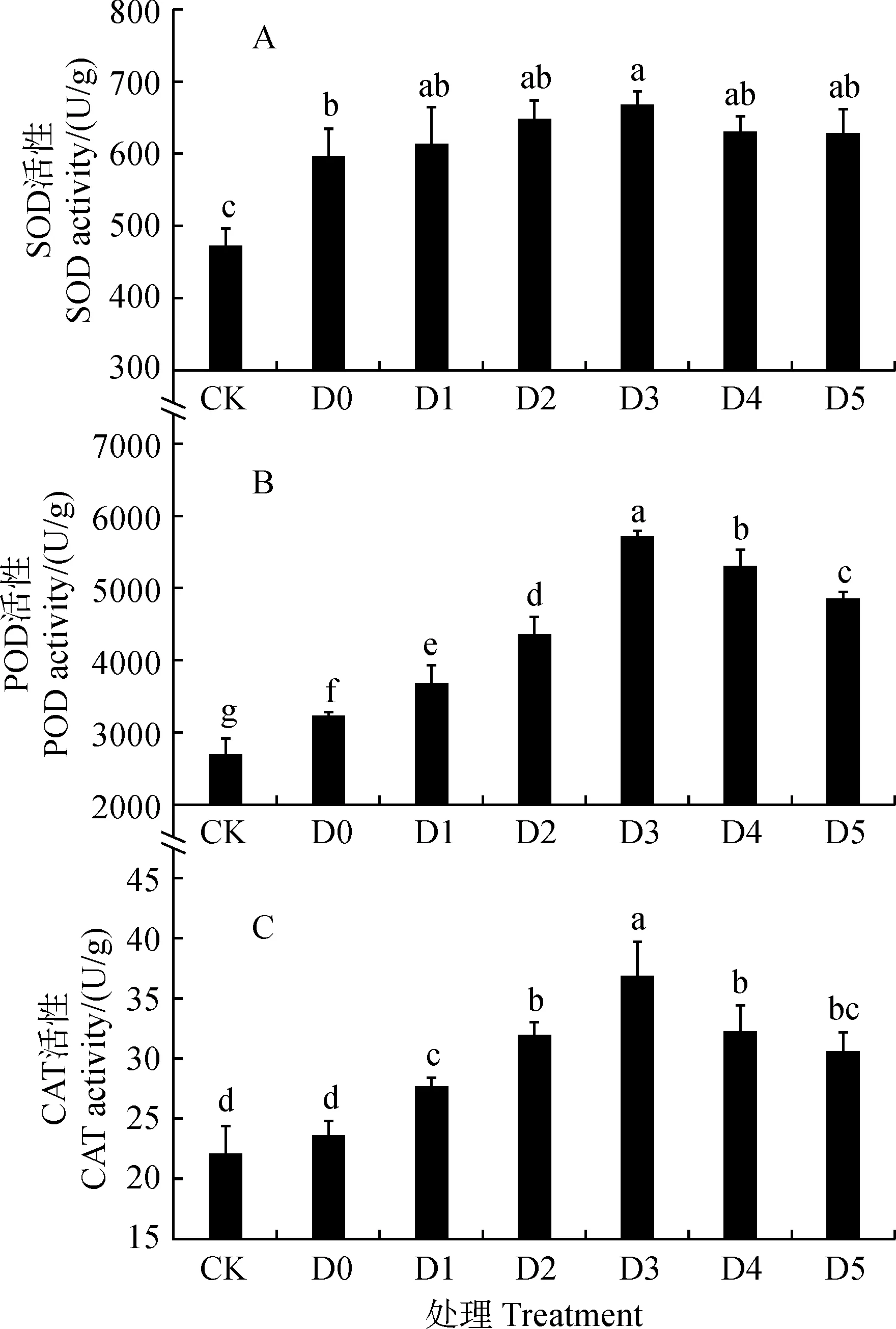
图2 干旱胁迫下喷施黄腐酸对燕麦叶片抗氧化酶活性的影响Fig.2 Effects of fulvic acid on antioxidant enzyme activities in A. sativa leaves under drought stress
3 讨 论
有研究表明,黄腐酸可以缓解干旱对甘蔗[18]、马铃薯[19]、黄瓜[20]的伤害,提高其耐旱性,且存在浓度效应。本试验发现,喷施200~1 000 mg/L FA可以降低干旱胁迫下燕麦叶片的膜脂过氧化伤害,提高燕麦的耐旱性,这与前人研究结果一致,而本试验条件下以600 mg/L FA的综合效果最好,与前人研究结果在浓度上有所差异,可能是因为所施用的作物不同,作物所能吸收的量也不同。
气孔是植物与周围空气进行气体交换的重要通道,为植物的光合作用提供原料CO2。干旱胁迫下光合速率受抑制有气孔因素和非气孔因素两种原因,气孔因素是叶片气孔关闭,从环境中吸收CO2的途径受到阻碍,从而降低光合速率;非气孔因素是外界环境的变化导致叶肉细胞光合活性降低,光合速率降低[21-23]。本研究表明单独干旱胁迫处理下,燕麦叶片的Pn、Gs和Tr均显著降低,而Ci显著增加,说明燕麦幼苗光合速率的下降是由非气孔因素导致的,究其原因可能是干旱破坏了叶绿体结构的稳定性,使叶片光合机构损伤,光合活性下降,导致CO2的固定和转化受到抑制,Ci升高[24-25]。Anjume等[26]研究表明黄腐酸能促进光合作用,降低气孔开放状态和蒸腾速率,从而促进生长,减少水分流失。本试验结果显示600 mg/L黄腐酸能显著提高干旱胁迫下燕麦叶片Tr、Pn和Gs,降低Ci,可能是黄腐酸通过提高干旱胁迫下植物体内光合作用过程中相关碳同化酶的活性来增强植物光合能力[11]。
光合色素是植物进行光合作用吸收、传递、转化光能的主要色素[27]。干旱胁迫导致植物叶片失水,破坏类囊体膜结构,影响叶绿素的合成,甚至促进已合成的叶绿素分解,使得叶绿素和类胡萝卜素含量降低[28-29]。本试验结果表明,干旱胁迫下燕麦叶片的光合色素含量明显降低,与上述研究结果相似。但也有研究发现,干旱胁迫使叶片含水量下降,叶片扩展生长受阻,导致叶片单位面积的叶绿素含量升高[30],与本研究结果相反,说明不同类型作物对干旱胁迫的反应不同。刘彩娟等[20]研究表明,喷施黄腐酸能提高干旱胁迫下黄瓜的光合色素含量。本试验发现,干旱胁迫下喷施600 mg/L黄腐酸能显著增加燕麦光合色素含量,说明黄腐酸可以缓解干旱胁迫对燕麦幼苗叶绿素的降解,增强燕麦叶片对光能的吸收和利用,进而提高干旱条件下燕麦的Pn,这与姜磊[31]在黄果柑抗旱性研究中的结果一致。
叶绿素荧光技术是观测PSⅡ活性、功能以及电子传递方面的探针[32],逆境胁迫造成PSⅡ反应中心损伤,使光合电子传递和PSⅡ的光合作用活力受到抑制,光合速率下降[33]。本试验中,干旱胁迫导致燕麦叶片Fv/Fm、qP、ETR和ΦPSⅡ的显著下降,Fo、NPQ显著上升。说明干旱胁迫使燕麦叶片的PSⅡ反应中心活性和开放程度受损,能量耗散途径受到抑制,PSⅡ反应中心的过剩光能大量积累,光合机构受到伤害,从而导致其光能利用能力降低[34]。黄腐酸喷施缓解了干旱胁迫下Fv/Fm、qP、ETR和ΦPSⅡ的下降,Fo、NPQ的上升。说明黄腐酸能提高PSⅡ对光能的捕获能力和转化效率,修复干旱胁迫对PSⅡ反应中心的损伤、降低热耗散程度,增加光合电子传递速率,维持光合机构的稳定,从而提高光合效率,有效缓解了干旱胁迫对燕麦幼苗生长的抑制作用,这与梁太波等[35]在腐植酸处理旱作小麦上的研究结果相一致。
正常情况下植株体内活性氧的产生与消除处于动态平衡,当遇到逆境胁迫时,这种动态平衡会被破坏,此时植株通过提高抗氧化酶活性来清除自由基,从而缓解胁迫对植株所带来的伤害[36]。本试验发现,干旱胁迫下,燕麦叶片中的抗氧化酶 SOD、POD、CAT活性显著高于CK,说明燕麦在遭受到干旱胁迫后,通过自身调节机制使SOD、POD、CAT活性增强,使植株适应干旱环境的能力提高。Ramin等[37]研究发现,喷施黄腐酸可明显提高小麦等作物体内SOD、CAT活性。王娟等[38]结果表明,腐植酸处理下的鸡冠花幼苗SOD等保护酶活性提高,减缓了水分胁迫造成的伤害。本研究中,600 mg/L黄腐酸处理显著增强了干旱胁迫下燕麦幼苗的SOD、POD和CAT的活性,且随着黄腐酸浓度的升高呈先增加后降低的趋势,这与周海涛等在皮燕麦上报道的黄腐酸提高干旱胁迫下SOD、POD和CAT活性的研究结果一致[12],说明600 mg/L黄腐酸对细胞膜结构具有良好的修复和保护作用,维持了细胞内环境的稳定,有效缓解了干旱胁迫造成的氧化损伤,增强了燕麦幼苗的抗旱性。
4 结 论
黄腐酸通过缓解干旱胁迫下燕麦叶片的光合作用的减弱,提高光合色素含量及抗氧化酶活性,降低PSⅡ的损伤程度,来改善植株光合能力、维持抗氧化酶系统相对稳定,进而缓解活性氧、自由基对植物的氧化损伤,抑制干旱胁迫对其造成的伤害,并具有浓度效应,以600 mg/L的黄腐酸缓解作用效果最佳。
