矢车菊素-3-O-葡萄糖苷对H9c2心肌细胞衰老影响
2022-11-16周宏稷田惠张蕊常晓天安毅
周宏稷 田惠 张蕊 常晓天 安毅



[摘要] 目的 探討矢车菊素-3-O-葡萄糖苷(C3G)对H9c2细胞衰老的影响及其机制。
方法 应用D-gal构建H9c2心肌细胞衰老模型,并筛选出C3G最佳干预浓度。将正常H9c2心肌细胞分为正常对照组(应用PBS处理48 h)、D-gal处理组(应用10 g/L浓度D-gal处理48 h)、C3G处理组(应用1 000 μmol/L浓度C3G处理48 h)、C3G+D-gal组(应用1 000 μmol/L浓度C3G与10 g/L浓度D-gal共同处理48 h)。检测各组细胞增殖活性、细胞凋亡水平、细胞内活性氧(ROS)水平;应用SA-β-半乳糖苷酶(SA-β-gal)染色试剂盒测定SA-β-gal表达,应用实时荧光定量PCR方法检测CD38、Sirtuin 6(Sirt6)、己糖激酶(HK2)、乳酸脱氢酶A(LDHA)和端粒酶逆转录酶(TERT)的mRNA表达,WST-8方法检测烟酰胺腺嘌呤二核苷酸(NAD+)的表达。
结果 使用10 g/L D-gal诱导H9c2心肌细胞48 h成功构建衰老心肌细胞模型,并筛选出C3G的最佳干预浓度为1 000 μmol/L。应用1 000 μmol/L浓度C3G进行干预,结果显示C3G可以抑制D-gal所诱导的衰老心肌细胞增殖水平降低(F=189.50,P<0.01)、凋亡水平升高(F=73.12,P<0.01)、细胞内ROS水平升高(F=106.00,P<0.01)以及SA-β-gal阳性细胞升高,降低D-gal诱导的衰老心肌细胞CD38、LDHA、HK2 mRNA 表达(F=23.14~94.00,P<0.01),同时升高TERT、Sirt6 mRNA表达(F=43.24、86.81,P<0.01)及NAD+的水平(F=77.02,P<0.01)。
结论 C3G通过抑制CD38的表达和提高Sirt6的表达,干预心肌细胞凋亡和增殖及糖酵解酶的变化,影响ROS的产生,从而发挥减轻心肌细胞衰老损伤的作用。
[关键词] 细胞衰老;矢车菊素-3-O-葡萄糖苷;CD38;Sirtuin 6
[中图分类号] R329.25;R916.4
[文献标志码] A
[文章编号] 2096-5532(2022)05-0708-06
doi:10.11712/jms.2096-5532.2022.58.161
[开放科学(资源服务)标识码(OSID)]
[网络出版] https://kns.cnki.net/kcms/detail/37.1517.R.20221021.1423.012.html;2022-10-24 09:08:57
EFFECT OF CYANIDIN-3-O-GLUCOSIDE ON SENESCENCE OF H9C2 CARDIOMYOCYTES
ZHOU Hongji, TIAN Hui, ZHANG Rui, CHANG Xiaotian, AN Yi
(Department of Cardiology, The Affiliated Hospital of Qingdao University, Qingdao 266003, China)
[ABSTRACT] Objective To investigate the effect of cyanidin-3-O-glucoside (C3G) on H9c2 cell senescence and its mechanism.
Methods D-gal was used to establish a model of H9c2 cardiomyocyte senescence, and the optimal concentration of C3G was determined. Normal H9c2 cardiomyocytes were divided into normal control group (PBS treatment for 48 h), D-gal treatment group (10 g/L D-gal treatment for 48 h), C3G treatment group (1 000 μmol/L C3G treatment for 48 h), and C3G+D-gal group (co-treatment with 1 000 μmol/L C3G and 10 g/L D-gal for 48 h). Cell proliferation, cell apoptosis level, and intracellular reactive oxygen species (ROS) level were measured for each group; the SA-β-galactosidase (SA-β-gal) staining kit was used to measure the expression of SA-β-gal; quantitative real-time PCR was used to measure the mRNA expression of CD38, Sirtuin 6 (Sirt6), he-xokinase (HK2), lactic acid dehydrogenase A (LDHA), and telomerase reverse transcriptase (TERT); WST-8 assay was used to measure the expression of nicotinamide adenine dinucleotide (NAD+).
Results H9c2 cardiomyocytes were induced by 10 g/L D-gal for 48 h to successfully establish a model of cardiomyocyte senescence, and the optimal concentration of C3G was determined as 1 000 μmol/L. After intervention with 1 000 μmol/L C3G, the results showed that C3G inhibited the reduction in cell proliferation (F=189.50,P<0.01), the increase in cell apoptosis level (F=73.12,P<0.01), the increase in intracellular ROS level (F=106.00,P<0.01), and the increase in the proportion of SA-β-gal-positive cells induced by D-gal in senescent cardiomyocytes, and it also reduced the mRNA expression of CD38, LDHA, and HK2 (F=23.14-94.00,P<0.01) and increased the mRNA expression of TERT and Sirt6 (F=43.24,86.81;P<0.01) and the level of NAD+ (F=77.02,P<0.01) in D-gal-induced senescent cardiomyocytes.
Conclusion C3G can alleviate the senescence of cardiomyocytes by inhibiting the expression of CD38, upregulating the expression of Sirt6, intervening against the apoptosis and proli-feration of cardiomyocytes and the change in glycolytic enzyme, and affecting the production of ROS.
[KEY WORDS] cell senescence; cyanidin 3-O-glucoside; CD38; Sirtuin 6
细胞衰老和机体衰老存在因果关系,组织器官内衰老细胞聚集将影响机体正常的生物功能。因D-半乳糖(D-gal)致衰老模型构建简便,有较高可行性与稳定性,被广泛应用于基础实验中。矢车菊素-3-O-葡萄糖(C3G)为普遍存在于自然界中的一种稳定的花青素,具有抗氧化、抗炎等作用,对于延缓衰老具有重要意义。但其是否能干预心肌衰老进程及机制犹未明确。研究发现C3G具有CD38抑制剂的作用,而CD38的表达与衰老存在相关性。CD38作为细胞中烟酰胺腺嘌呤二核苷酸(NAD+)主要的代谢酶,在与年龄相关的衰老伴随的NAD+衰退中起着核心作用。前期研究发现,NK细胞中CD38高表达抑制了Sirtuin 6(Sirt6)的表达。有研究显示,C3G参与影响Sirt6功能,Sirt6通过沃伯格效应干预糖酵解酶的变化与衰老进程相关。本研究拟采用C3G干预D-gal诱导的大鼠H9c2心肌细胞衰老模型,探讨C3G对心肌细胞衰老的影响及其机制。
1 材料与方法
1.1 实验材料
大鼠心肌细胞H9c2购于中国科学院典型培养物保藏委员会细胞库。C3G、D-gal(索莱宝,中国);CCK-8细胞增殖及毒性检测试剂盒(MCE,美国); DMEM高糖培养液、胎牛血清(普诺赛公司,中国);Annexin V-FITC / PI荧光双染细胞凋亡检测试剂盒(Elabscience,中国);细胞衰老相关β-半乳糖苷酶(SA-β-gal)染色试剂盒、活性氧(ROS)检测试剂盒(碧云天,中国);大鼠Sirt6、CD38、乳酸脱氢酶A(LDHA)、端粒酶逆转录酶(TERT)、己糖激酶Ⅱ(HK2)引物(生工,中国); 氧化型辅酶Ⅰ/还原型辅酶Ⅰ(NAD+/NADH)检测试剂盒(碧云天,中国);RNA iso Plus(Takara,日本);HiScript Ⅲ RT SuperMix、SYBR qPCR Master Mix(诺唯赞,中国)。
1.2 D-gal与C3G最佳干预浓度筛选及实验分组
H9c2细胞在含有体积分数0.10胎牛血清、100 U/L青霉素、100 U/L链霉素的DMEM培养液中,37 ℃、体积分数0.05 CO培养箱中培养12 h,待细胞贴壁后,分别用0.1、0.5、1.0、5.0、10.0 g/L浓度的D-gal处理48 h,应用CCK-8方法测定细胞增殖水平,筛选构建衰老模型D-gal的最佳浓度。应用CCK-8方法,检测10、50、100、500、1 000 μmol/L濃度C3G对D-gal处理后H9c2细胞与正常H9c2细胞增殖水平的影响,筛选出C3G的最佳干预浓度。将正常H9c2细胞分为正常对照组(A组)、D-gal处理组(B组)、C3G处理组(C组)、D-gal+C3G组(D组)。正常对照组:H9c2细胞加入PBS处理48 h;D-gal处理组:应用10 g/L D-gal处理H9c2细胞48 h;C3G处理组:应用1 000 μmol/L浓度C3G处理H9c2细胞48 h;D-gal+C3G组:应用1 000 μmol/L浓度C3G与10 g/L浓度D-gal共同处理H9c2细胞48 h。
1.3 检测指标及方法
1.3.1 细胞增殖活性检测 将未处理H9c2细胞悬液以每孔100 μL接种入96孔板内,按照1.2方法分组处理后,弃上清。重新加入含血清的DMEM高糖培养液,每孔内加入CCK-8试剂10 μL,37 ℃培养2 h。应用Infinite 200 Pro荧光酶标仪检测450 nm处吸光度值,以其表示细胞增殖水平。
1.3.2 细胞凋亡水平检测 将处理后的各组细胞用PBS洗涤,使用胰酶消化并离心,重悬于Anne-xin V Binding Buffer(1×)中。分别加入Annexin V-FITC Reagent与Propidium Iodide(PI) Staining Solution,避光孵育20 min。使用Apogee A50流式细胞仪(英国)分析细胞染色情况,以AnnexinV-FITC(+)/PI(+)、AnnexinV-FITC(+)/PI(-)细胞占比之和计为凋亡细胞总数。
1.3.3 SA-β-gal染色 使用SA-β-gal染色试剂盒测定SA-β-gal表达,按照试剂盒说明书进行操作。应用相差显微镜(100倍)观察各组细胞染色情况。衰老细胞表现为高表达SA-β-gal,SA-β-gal染色阳性细胞呈青蓝色,根据其比例判定细胞衰老状态。
1.3.4 细胞内ROS检测 应用 ROS检测试剂盒检测各组细胞ROS水平,按试剂盒说明书进行操作。使用荧光酶標仪检测ROS水平,激发波长为488 nm,发射波长为525 nm。
1.3.5 实时荧光定量PCR(qPCR)检测CD38、Sirt6、LDHA、HK2和TERT mRNA的表达 使用RNA iso Plus从H9c2细胞中提取总RNA,利用HiScript Ⅲ RT将提取的RNA逆转录为cDNA。将cDNA添加到ChamQ Universal SYBR qPCR Master Mix中,使用BIOER9660 FQD-96A荧光定量PCR仪(中国)检测CD38、Sirt6、LDHA、HK2和TERT mRNA表达,采用2方法计算结果。以β-actin mRNA作为内参。qPCR引物及其序列见表1。
1.3.6 WST-8法检测细胞NAD+的表达 应用NAD+/NADH检测试剂盒检测各组NADtotal及NADH的表达,根据说明书要求操作。荧光酶标仪检测450 nm处吸光度值,以NADtotal水平-NADH水平表示NAD+水平。
1.4 统计学分析
应用SPSS 24.0 软件进行统计学处理,计量资料结果以±s表示,数据间比较采用单因素方差分析及两因素析因设计的方差分析。以P<0.05为差异有统计学意义。
2 结 果
2.1 构建H9c2心肌细胞衰老模型的D-gal最佳作用浓度
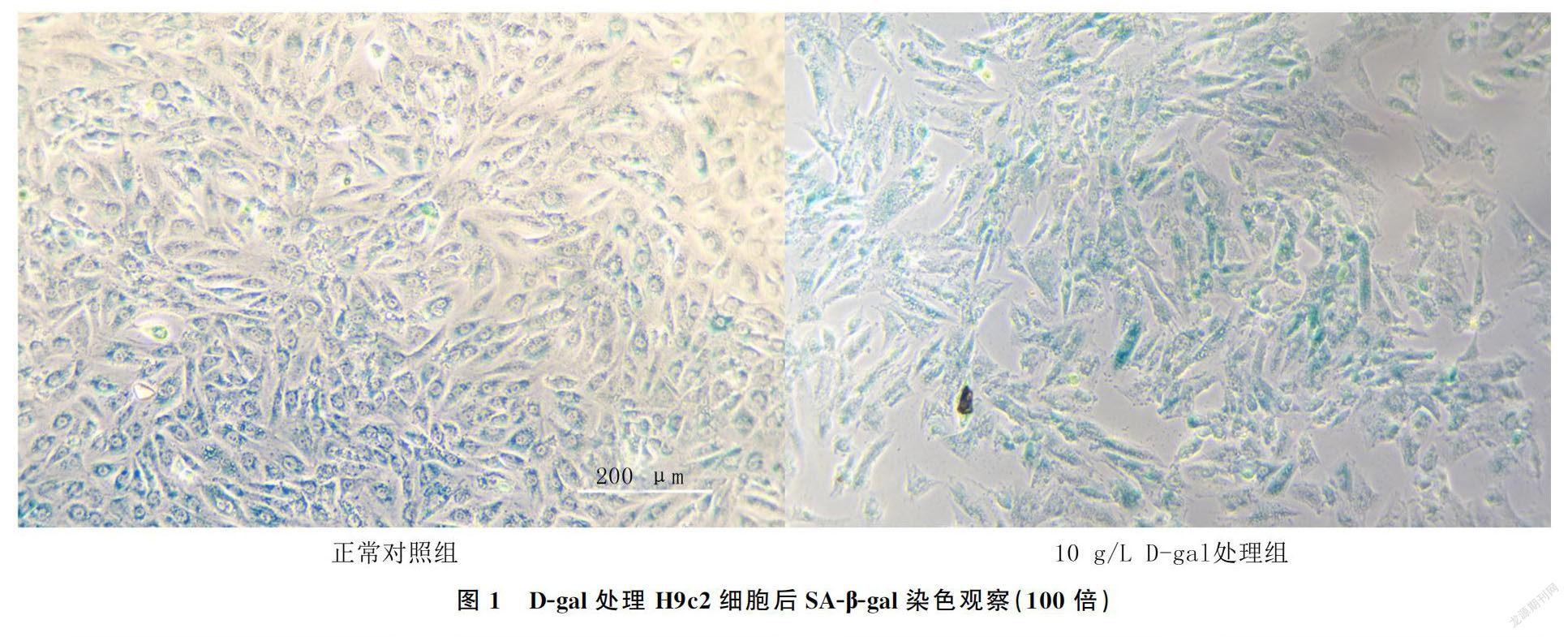
正常对照组及0.1、0.5、1.0、5.0、10.0 g/L浓度D-gal处理组H9c2细胞增殖水平分别为2.44±0.03、2.17±0.13、2.10±0.16、2.02±0.07、1.89±0.02、1.75±0.02。不同浓度D-gal处理组细胞增殖活性均较正常对照组降低,差异有统计学意义(F=21.64,P<0.01);D-gal浓度为10 g/L时细胞增殖活性最低,为D-gal构建衰老模型的最佳浓度。SA-β-gal染色结果显示,10 g/L浓度D-gal处理组的阳性细胞比例较正常对照组明显增多(图1)。
2.2 C3G对H9c2心肌细胞的最佳干预浓度
CCK-8检测显示,正常对照组及10、50、100、500、1 000 μmol/L C3G处理H9c2细胞的增殖活性分别为1.76±0.04、1.59±0.13、1.90±0.16、1.98±0.07、1.99±0.02、2.00±0.07,100、500、1 000 μmol/L浓度C3G处理组细胞增殖水平较正常对照组升高,差异有统计学意义(F=9.43,P<0.01),其中1 000 μmol/L浓度C3G处理组细胞增殖水平最高。正常对照组、D-gal处理组及10、50、100、500、1 000 μmol/L C3G处理心肌细胞衰老模型组H9c2细胞的增殖活性分别为2.44±0.03、1.75±0.02、1.82±0.31、1.82±0.08、2.28±0.22、2.34±0.04、2.49±0.04。100、500、1 000 μmol/L C3G处理心肌细胞衰老模型组细胞增殖水平较D-gal处理组升高,差异有统计学意义(F=14.05,P<0.01),其中1 000 μmol/L浓度C3G处理组细胞增殖水平最高。
2.3 C3G对D-gal处理的H9c2细胞增殖、凋亡以及SA-β-gal染色的影响
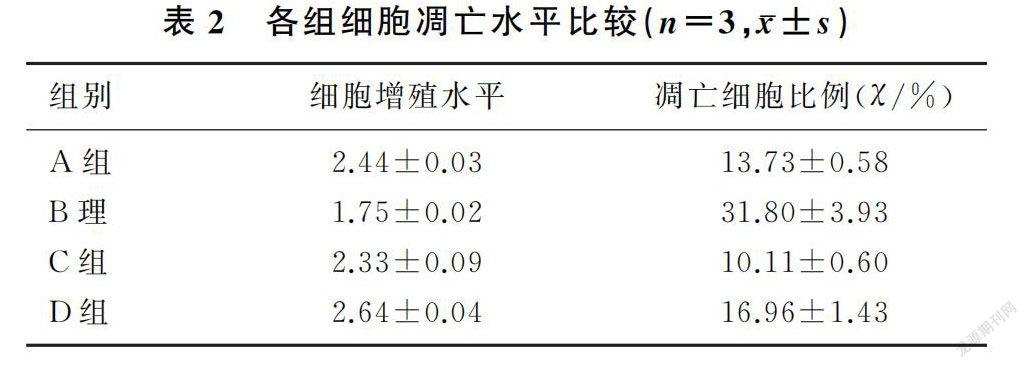
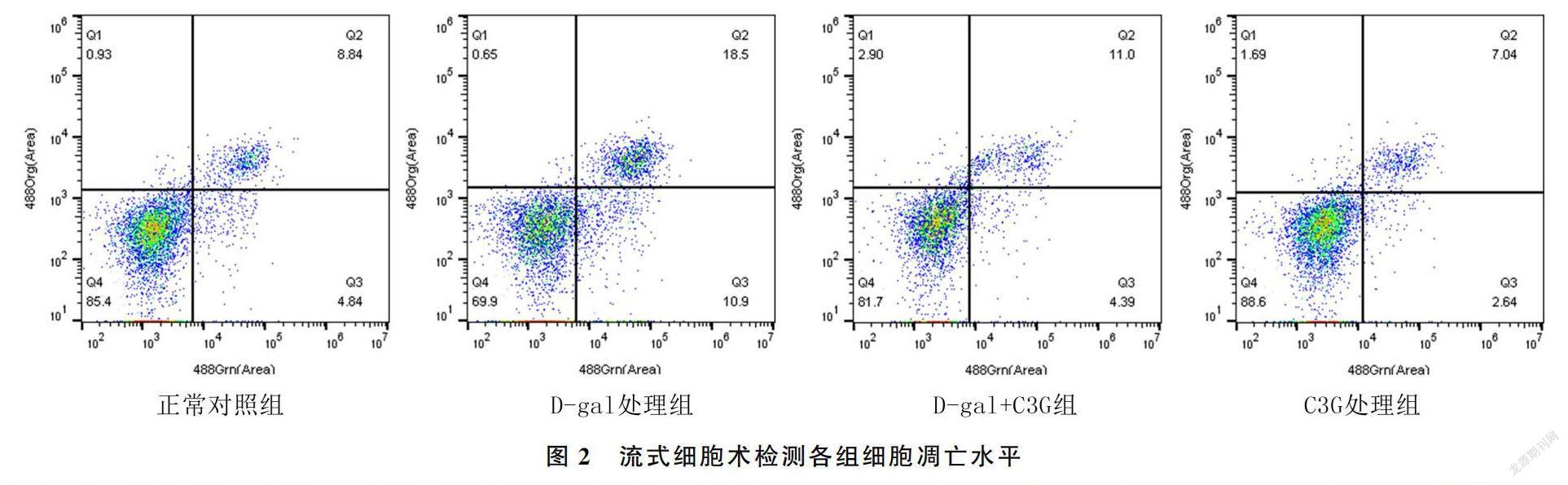
在C3G未干预的情况下,D-gal处理组相较于D-gal未处理组细胞的增殖水平明显降低(F=265.85,P<0.01),凋亡水平明显增高(F=108.465,P<0.01);在C3G干预情况下,D-gal处理组相较于D-gal未处理组细胞增殖水平明显降低(F=53.11,P<0.01),而凋亡水平明显增高(F=20.43,P<0.05)。在D-gal未干预情况下,C3G处理组相较于C3G未处理组细胞增殖活性升高(F=22.55,P<0.01),凋亡水平降低(F=7.06,P<0.05);而在D-gal干预的情况下,C3G处理组相较于C3G未处理组细胞增殖活性增高(F=189.50,P<0.01),凋亡水平降低(F=73.12,P<0.01)。见表2、图2。SA-β-gal染色结果显示,与正常对照组比较,D-gal处理组阳性细胞比例显著升高;与D-gal处理组比较,C3G处理组阳性细胞比例显著降低(图3)。
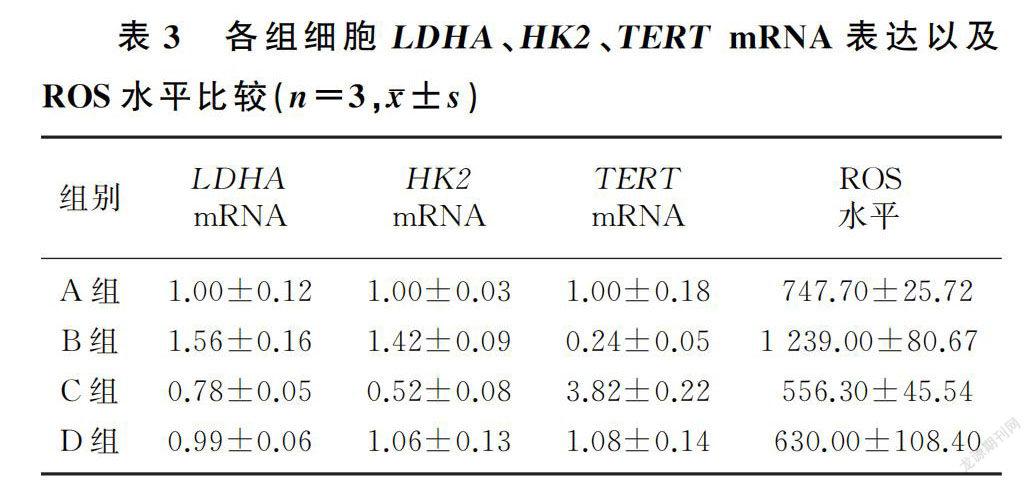
2.4 D-gal及C3G处理对细胞糖酵解酶、ROS以及衰老相关蛋白酶TERT变化的影响在C3G未干预的情况下,D-gal处理组相较于D-gal未处理组LDHA、HK2 mRNA表达及ROS水平明显增高(F=31.71~69.00,P<0.01),TERT mRNA表达降低(F=38.37,P<0.01);在C3G干预的情况下,D-gal处理组相较于D-gal未处理组LDHA、HK2 mRNA表达明显增高(F=11.67、83.53,P<0.05),TERT mRNA表达水平明显降低(F=507.91,P<0.01),而ROS水平无明显差异(F=1.55,P>0.05)。在D-gal未干预情况下,C3G处理组相较于C3G未处理组细胞LDHA mRNA表达(F=12.37,P<0.01)、HK2 mRNA表达(F=69.20,P<0.01)及ROS水平(F=10.46,P<0.05)明显降低,TERT mRNA表达水平明显升高(F=525.23,P<0.01)。而在D-gal干预情况下,C3G处理组相较于C3G未处理组细胞LDHA mRNA表达(F=50.11,P<0.01)、HK2 mRNA表达(F=23.14,P<0.01)以及ROS水平(F=106.00,P<0.01)明显降低,TERT mRNA水平明显升高(F=43.24,P<0.01)。见表3。
2.5 D-gal和C3G处理对CD38、Sirt6 mRNA表达以及NAD+水平的影響
在C3G未干预的情况下,D-gal处理组相较于D-gal未处理组CD38 mRNA表达明显增高(F=94.00,P<0.01),Sirt6 mRNA表达(F=185.30,P<0.01)及细胞内NAD+的水平(F=112.20,P<0.01)明显减低;在C3G干预的情况下,D-gal处理组相较于D-gal未处理组CD38 mRNA表达明显增高(F=15.58,P<0.01),Sirt6 mRNA表达(F=289.16,P<0.01)及细胞内NAD+的水平(F=52.40,P<0.01)明显减低。在D-gal未干预情况下,C3G处理组细胞相较于C3G未处理组细胞CD38 mRNA表达水平降低(F=19.50,P<0.01),Sirt6 mRNA表达(F=161.52,P<0.01)及细胞内NAD+水平(F=29.41,P<0.01)升高;而在D-gal干预情况下,C3G处理组相较于C3G未处理组细胞CD38 mRNA的表达水平降低(F=103.33,P<0.01),Sirt6 mRNA表达及细胞内NAD+的水平升高(F=86.81、77.02,P<0.01)。见表4。
3 讨 论
衰老是机体和细胞水平上由多因素调节所导致的自然进程。细胞增殖与凋亡作为相互对应两方面,与衰老有着不可分割的联系。NAD+作为生命体的必要成分,随着年龄增加其水平逐渐降低,其涉及衰老的多种分子机制,如氧化应激与DNA损伤等;其也作为糖酵解的重要中间代谢产物维持糖酵解通量。CD38作为负责催化NAD+代谢酶,可能是NAD+水平随着年龄的增长而下降的主要原因。通过抑制CD38可逆转动物衰老及年龄升高所导致的组织内NAD+水平的下降,CD38并被认为是延长寿命的药理靶点。花青素是植物中的天然色素,对人体有着重要的营养及保护价值;C3G是桑椹花青素内的最主要与常见的成分之一,其对个体寿命影响的研究较多。包括C3G在内的花青素作为CD38的抑制剂,通过消耗NAD+,阻止CD38催化环腺苷二磷酸核糖的合成发挥作用。Sirt6自发现以来被冠以“长寿基因”的称号,也是长寿物种中有效的DNA双链断裂修复的关键基因。糖酵解为糖类物质的主要代谢方式,参与多种细胞衰老机制的调控。糖酵解酶HK2与LDHA的表达,不仅受Sirt6的抑制还与衰老相关。自由基衰老学说的核心内容是细胞氧化应激导致ROS过度生成及消除能力降低所引起的平衡性破坏,ROS表达与糖酵解酶LDHA和HK2都有着密切联系。同时ROS的过度聚集会导致端粒的损伤,而TERT可起到维持端粒稳定性的作用。TERT作为衰老相关蛋白酶,可反映细胞内端粒的状态,TERT表达水平与DNA的消耗和细胞衰老及DNA损伤有着重要联系。TERT不仅可以作为判定衰老水平的工具,也可以作为衰老机制探讨的靶点。
本研究通过检测细胞增殖活性水平,筛选出10 g/L浓度D-gal与1 000 μmol/L浓度C3G作为构建H9c2心肌细胞衰老模型以及C3G干预的最适浓度;应用D-gal处理后,衰老细胞增殖水平减低,凋亡水平增加;而C3G可以有效逆转衰老细胞增殖水平的降低、凋亡水平的增高;C3G能抑制正常细胞凋亡,促进其增殖;而且C3G能降低衰老心肌细胞SA-β-gal高表达细胞比例和ROS水平,升高TERT mRNA表达。提示C3G能减轻D-gal所诱导的心肌细胞衰老损伤。
本文对C3G干预衰老进程机制研究结果显示,衰老的H9c2细胞CD38 mRNA表达升高,Sirt6 mRNA表达及NAD+含量降低,C3G干预后可减轻这种衰老的变化;C3G干预可以降低正常细胞CD38 mRNA的表达,增加Sirt6 mRNA的表达及NAD+的水平。提示C3G可以通过干预CD38与Sirt6的表达进而影响D-gal构建的细胞模型衰老的发生与发展,且C3G作为CD38的抑制剂可以参与影响Sirt6的表达。C3G会降低正常细胞与衰老心肌细胞内LADH与HK2 mRNA的表达,提示C3G可通过影响Sirt6干预糖酵解进程最终参与衰老进程。
综上所述,D-gal通过影响CD38和Sirt6的表达以及NAD+水平干预衰老进程。Sirt6可通过沃伯格效应干预HK2及LDHA的表达进而影响细胞衰老的糖代谢水平。C3G可降低CD38的表达,升高NAD+水平,进而升高Sirt6表达而干预衰老进程中糖酵解酶的变化,减轻ROS所诱发的氧化应激损伤,发挥减轻D-gal所致心肌细胞衰老损伤的作用。
[参考文献]
[1]赵焕东,胥洪鹃,李坚,等. 高剂量D-半乳糖复制C57BL/6J衰老小鼠模型的研究[J]. 中国现代医学杂志, 2019,29(16):10-16.
[2]刘迪,张浩,张寒雪,等. 花青素和矢车菊素的生物活性及抗氧化功能研究进展[J]. 吉林医药学院学报, 2021,42(1):58-60.
[3]CHEN W, MLLER D, RICHLING E, et al. Anthocyanin-rich purple wheat prolongs the life span of Caenorhabditis elegans probably by activating the DAF-16/FOXO transcription factor[J]. Journal of Agricultural and Food Chemistry, 2013,61(12):3047-3053.
[4]KELLENBERGER E, KUHN I, SCHUBER F, et al. Flavonoids as inhibitors of human CD38[J]. Bioorganic & Medicinal Chemistry Letters, 2011,21(13):3939-3942.
[5]GERASIMENKO M, LOPATINA O, SHABALOVA A A, et al. Distinct physical condition and social behavior phenotypes of CD157 and CD38 knockout mice during aging[J]. PLoS One, 2020,15(12):e0244022.
[6]CHINI C, HOGAN K A, WARNER G M, et al. The NADase CD38 is induced by factors secreted from senescent cells providing a potential link between senescence and age-related cellular NAD+ decline[J]. Biochemical and Biophysical Research Communications, 2019,513(2):486-493.
[7]WANG H X, LI S T, ZHANG G Q, et al. Potential therapeutic effects of cyanidin-3-O-glucoside on rheumatoid arthritis by relieving inhibition of CD38+ NK cells on Treg cell differen-tiation[J]. Arthritis Research & Therapy, 2019, 21(1):220.
[8]RAHNASTO-RILLA M, TYNI J, HUOVINEN M, et al. Natural polyphenols as sirtuin 6 modulators[J]. Scientific Reports, 2018,8(1):4163.
[9]PILLAI V B, SAMANT S, HUND S, et al. The nuclear sirtuin SIRT6 protects the heart from developing aging-associated myocyte senescence and cardiac hypertrophy[J]. Aging, 2021,13(9):12334-12358.
[10]董振. SIRT6通過CDK4-CCND1复合体调控沃伯格效应[D]. 重庆:西南大学, 2017.
[11]张竣,张莹,王树坤,等. 非受体酪氨酸激酶对糖氧剥夺诱导的少突胶质细胞凋亡的影响及其机制[J]. 精准医学杂志, 2021,36(3):207-211.
[12]李靳,杨莉,万静枝,等. 三七总皂苷对D-半乳糖致H9c2细胞衰老的保护作用研究[J]. 中药材, 2014,37(8):1421-1424.
[13]WATT K I, HARVEY K F, GREGOREVIC P. Regulation of tissue growth by the mammalian hippo signaling pathway[J]. Frontiers in Physiology, 2017,8:942.
[14]GOMES A P, PRICE N L, LING A J, et al. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging[J]. Cell, 2013,155(7):1624-1638.
[15]SCHEIBYE-KNUDSEN M, MITCHELL S J, FANG E F, et al. A high-fat diet and NAD(+) activate Sirt1 to rescue premature aging in cockayne syndrome[J]. Cell Metabolism, 2014,20(5):840-855.
[16]CANTÓC, MENZIES K J, AUWERX J. NAD(+) metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus[J]. Cell Metabolism, 2015,22(1):31-53.
[17]TARRAGÓ M G, CHINI C C S, KANAMORI K S, et al. A potent and specific CD38 inhibitor ameliorates age-related me-
tabolic dysfunction by reversing tissue NAD decline[J]. Cell Metabolism, 2018,27(5):1081-1095.e10.
[18]CHINI E N. CD38 as a regulator of cellular NAD: a novel potential pharmacological target for metabolic conditions[J]. Current Pharmaceutical Design, 2009,15(1):57-63.
[19]ROICHMAN A, ELHANATI S, AON M A, et al. Restoration of energy homeostasis by SIRT6 extends healthy lifespan[J]. Nature Communications, 2021,12:3208.
[20]TIAN X, FIRSANOV D, ZHANG Z H, et al. SIRT6 is responsible for more efficient DNA double-strand break repair in long-lived species[J]. Cell, 2019,177(3):622-638.e22.
[21]孫慧. miR-141-3p靶向PFKM影响细胞糖代谢参与调控间充质干细胞衰老的作用研究[D]. 长春:吉林大学, 2021.
[22]PAN C Y, LI B, SIMON M C. Moonlighting functions of me-
tabolic enzymes and metabolites in cancer[J]. Molecular Cell, 2021,81(18):3760-3774.
(本文编辑 黄建乡)
