水产动物牛磺酸营养研究进展
2022-10-31李正伟刘庆武王连生
■范 泽 李正伟 刘庆武 王连生*
(1.中国水产科学研究院黑龙江水产研究所,黑龙江省水生动物病害与免疫重点实验室,黑龙江哈尔滨 150070;2.黑龙江省水生动物资源养护中心,黑龙江哈尔滨 150070;3.黑龙江省双鸭山市农业技术推广总站,黑龙江双鸭山 155100)
为了应对鱼粉“价格走高、产量较低、依赖进口”的水产饲料行业现状,植物蛋白(豆粕、菜籽粕、玉米蛋白粉等)替代鱼粉和新型蛋白源开发成为国内外研究的热点方向。然而氨基酸平衡问题制约着植物蛋白源和新型蛋白源在水产饲料中的广泛应用。除了赖氨酸、蛋氨酸等十种必需氨基酸的缺乏限制外,某些功能性氨基酸的缺乏同样是限制植物蛋白源和新型蛋白源应用于水产饲料的关键因素,牛磺酸正是其中一种重要的功能性氨基酸。牛磺酸在水产动物体内常以含硫游离氨基酸或小分子二肽或三肽的形式存在。尽管被称为氨基酸,但由于其组成结构中所带的是磺酸基而不是羧基,造成其与传统氨基酸在功能方面存在一定差异,其中最主要的差异是不参与蛋白质合成,同时也不能作为能源物质,而其主要生物学功能在于调节机体正常生理机能,如形成结合胆汁酸、调节渗透压、维持细胞膜稳定性及提升抗氧化能力等。饲料中缺乏牛磺酸会导致鱼类生长缓慢,使机体产生绿色肝胆综合征、肠炎及贫血等症状。文章主要对水产动物饲料牛磺酸的来源、水产动物牛磺酸的合成能力、水产动物牛磺酸营养需求、参与水产动物营养物质的代谢、对水产动物健康的影响、牛磺酸与其他营养物质的相互作用等6 个方面进行综述,以期为牛磺酸在水产动物营养免疫与健康养殖方面的进一步研究提供科学理论基础。
1 水产动物牛磺酸的来源
1.1 外源供给
现有的研究表明水产动物自身合成牛磺酸的能力相对较弱,因此为满足自身需求则需要从饲料中获取,因此,牛磺酸对于水产动物而言是一种条件性必需氨基酸。在常用的水产饲料原料中,鱼粉等动物性蛋白原料为饲料配方中牛磺酸的主要提供者(见表1),而大多数植物性蛋白原料中牛磺酸含量较少或基本为零。但由于养殖环境恶化和饲料成本升高等产业现状,鱼粉等动物性蛋白原料被越来越多的植物性蛋白源所替代,牛磺酸成为主要限制性营养物质之一。因此,在未来饲料配方研发及新型蛋白源开发的过程中,如何满足水产动物牛磺酸营养需要成为主要关注点之一。
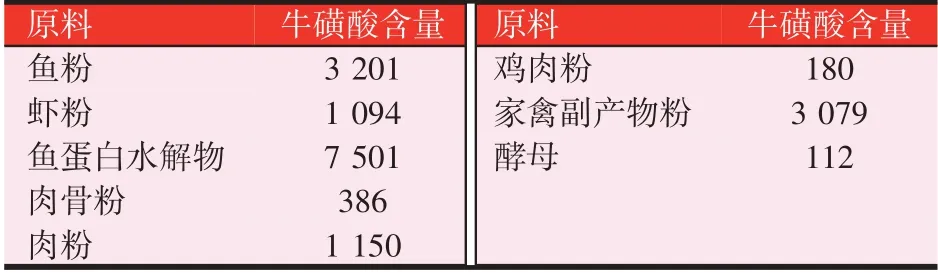
表1 常见水产饲料原料的牛磺酸含量(mg/kg)
1.2 内源合成能力
水产动物牛磺酸合成包括4 种途径(见图1),其中途径1 和途径2 为水产动物牛磺酸合成的主要途径。水产动物牛磺酸的合成能力主要取决于牛磺酸合成关键酶的活力。半胱次磺酸脱羧酶(cysteinesulfinate decarboxylase, CSD)和半胱胺双加氧酶(cysteine dioxygenase, CDO)分别是途径1 和途径2 的关键限速酶。目前关于水产动物牛磺酸合成能力的研究主要集中于鱼类,缺乏对于虾蟹类的研究。肝脏是鱼类牛磺酸合成的主要器官,表2列举了主要养殖鱼类肝脏牛磺酸合成关键酶的活力。研究表明鱼的种类是影响鱼类肝脏牛磺酸合成能力的重要因素。一般来说,海水鱼类的牛磺酸合成能力低于淡水鱼类,甚至无合成能力。不同鱼类肝脏牛磺酸合成途径存在差异,多数鱼类主要通过途径1 合成牛磺酸,而对于鲤鱼(Cyprinus carpio)、香鱼(Plecoglossus altivelis)及大太阳鱼(Lepomismm macrochirus)来说,当途径1 合成的牛磺酸无法满足自身需求时,仍可通过途径2进行合成。

表2 鱼类肝脏牛磺酸合成关键酶的活力

图1 水产动物牛磺酸合成途径(Salze等,2015)
2 水产动物牛磺酸需求量
近10年发表的水产动物牛磺酸需求量研究论文见表3。水产动物牛磺酸需求量受到生长阶段、评价指标、饲料中蛋白质组成及养殖环境的影响。在饲料蛋白源统一的条件下,体重为(6.87±0.12)g 和(38.1~38.9)g 草鱼(Ctenopharyngodon idella)的牛磺酸需求量分别为0.12%和0.06%;体重为(6.3±0.01)g 和(165.9±5.01)g 大菱鲆(Scophthalmus maximusL.)的牛磺酸需求量分别为1.15%和0.64%。而在(0.13±0.01)g 虹鳟(Oncorhynchus mykiss)鱼粉饲料中添加0.5%~1.5%牛磺酸未能促进生长,但在(203.26±2.01)g虹鳟鱼粉饲料中添加0.06%牛磺酸能显著提升生长性能。上述研究表明鱼的规格对于水产动物牛磺酸需求量的影响规律仍需进一步探索。在不同生长阶段,水产动物对于牛磺酸的需求量亦存在差异。以增重率为评价指标,尼罗罗非鱼(Oreochromis niloticus)幼鱼在28、56 d 及84 d 时的牛磺酸需求量分别为0.72%、0.68%及0.64%,在对斜带石斑鱼(Epinephelus coioides)幼鱼的研究中得到了类似的变化规律。评价指标的变化也影响着牛磺酸需求量,黄颡鱼(Pelteobagrus fulvidraco)以特定生长率(SGR)为评价指标时所得出的牛磺酸需求量低于以血清溶菌酶为评价指标时。鱼粉是水产动物饲料中牛磺酸的主要来源,而植物蛋白源中牛磺酸相对缺乏,因此植物蛋白源替代鱼粉时应考虑水产动物对牛磺酸的需求变化。田芊芊等(2016)发现在豆粕蛋白替代50%鱼粉的饲料中,青鱼(Mylopharyngodon piceus)牛磺酸需求量为0.1%。在对大口黑鲈(Micropterus salmoides)、红鳍东方鲀(Takifugu rubripes)、凡纳滨对虾(Litopenaeus vannamei)等研究中均得到了一致的结果。不同养殖环境对水产动物牛磺酸需求量有着潜在影响,李航等(2017)在淡水养殖条件下得出的(0.160±0.002)g 凡纳滨对虾牛磺酸的需要量为0.44%~0.58%。而Yue(2013)发现在正常海水盐度下0.48 g 凡纳滨对虾牛磺酸需要量为0.17%。

表3 水产动物牛磺酸需求量(占饲料的百分比)
3 牛磺酸参与水产动物营养物质代谢
3.1 脂肪代谢
在饲料中添加牛磺酸可以降低草鱼、斜带石斑鱼、龙胆石斑鱼(Epinephelus lanceolatus)、杂交条纹鲈(Morone chrysops×M.saxatilis)体脂肪含量。牛磺酸可与胆酸结合形成牛磺胆酸等结合胆汁酸,并通过乳化脂肪,增大脂肪与脂肪酶接触面积,促进脂肪消化吸收。饲料中添加0.33%牛磺酸能够避免塞内加尔鳎(Solea senegalensis)肝脏胆汁酸盐合成的中断并维持脂质消化吸收能力。此外,投喂添加牛磺酸0.05%、0.1%、0.15%或0.2%的饲料8 周的鲤鱼肠道脂肪酶活力均高于对照组,显著改善鲤鱼对饲料脂肪的消化和吸收能力,表明牛磺酸能够增强机体脂肪消化、分解及吸收能力。王清滨(2014)在高脂饲料中添加0.07%~0.17%牛磺酸可有效提高草鱼肝胰脏中脂肪分解酶类的活性,增加肝脏中三酰甘油的分解,并提高胆固醇7α-羟化酶(CYP7A1)的活性及其基因表达量,使胆固醇降解为胆汁酸,降低机体内脂肪含量。而在尼罗罗非鱼和凡纳滨对虾饲料中添加牛磺酸却会增加鱼体脂肪含量。因此,牛磺酸在水产动物脂肪代谢中可能发挥着关键作用,但在不同水产动物中的具体调控机制仍需深入研究。

表3(续) 水产动物牛磺酸需求量(占饲料的百分比)
3.2 糖代谢
牛磺酸能够缓解浅水胁迫应激引起的黄斑蓝子鱼(Siganus canali-culatus)和团头鲂(Mylopharyngodon piceus)血清血糖水平升高,也可以显著降低青鱼急性拥挤胁迫后的血清葡萄糖水平,表明牛磺酸能够参与水产动物葡萄糖稳态的调节。Zhang 等(2019)以未添加牛磺酸的15%糖水平组作为对照,采用2×3 双因素(糖水平为15%、30%;牛磺酸水平为0.4%、1.2%和2.0%)试验明确了在高糖饲料条件下牛磺酸能够通过显著提升大菱鲆肝脏糖酵解,葡萄糖转运,糖原合成能力,抑制糖异生水平,维持机体正常血糖水平,且在对肝脏糖酵解及葡萄糖转运载体基因表达影响上牛磺酸和饲料糖水平存在显著的交互作用。因此,牛磺酸对水产动物的糖代谢机能具有一定调节作用。
3.3 蛋白质代谢
尽管牛磺酸不参与水产动物体内蛋白质的合成,但在草鱼、斜带石斑鱼、鲤鱼、凡纳滨对虾及中华绒螯蟹(Eriocheir sinensis)的饲料中添加牛磺酸均能够提高水产动物机体蛋白质含量。对于饲料中添加适量牛磺酸增强机体蛋白质沉积的解释,目前可以归纳为3 种观点:首先,牛磺酸可提升水产动物体内蛋白质的消化吸收效率。伍琴等(2015)发现牛磺酸通过调节氨肽酶N(APN)、小肽转运蛋白(PepT1)、基因系尾型同源盒基因(CDX2)、L-氨基酸转运载体蛋白(LAT2)mRNA 在鲫鱼肠道中的相对表达量来调控蛋白质的水解,小肽的转运以及氨基酸的消化吸收。此外,牛磺酸能够调节蛋白质代谢相关激素的分泌。邱小琮等(2006)发现,0.2%、0.4%、0.6%及0.8%牛磺酸能够显著提高鲤鱼血清甲状腺激素含量,血清甲状腺激素决定着鱼类生长激素的基因表达和合成,而生长激素能够增强鱼类蛋白质合成能力。最后,外源性牛磺酸的补充能够减少含硫氨基酸用于合成牛磺酸的比例,提升蛋白质质量,提高蛋白质利用率。
4 牛磺酸对水产动物健康的影响
4.1 改善水产动物抗氧化能力及免疫力
补充适宜牛磺酸能够提升水产动物的抗氧化能力及免疫机能。孔圣超等(2020)在研究克氏原螯虾(Procambarus clarkii)时发现,在饲料中添加300 mg/kg牛磺酸可在4 周后显著提高肝胰脏超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活力,抑制丙二醛(MDA)的产生,提升血清中溶菌酶(LZM)的活性,增强鱼体抗氧化及免疫能力。Li 等(2016)研究表明全植物蛋白饲料中牛磺酸含量达到1.09%时可以显著提高黄颡鱼抗氧化酶活力,进而提升抗氧化能力。王清滨(2014)研究表明,高脂饲料中添加700~1 200 mg/kg牛磺酸显著提高了抗氧化酶活力,减轻脂质过氧化程度,缓解高脂对机体的氧化损伤。Kumar等(2009)对淡水鲶鱼(Clarias batrachus)研究发现,饲料中5 mg/kg牛磺酸通过降低肝脏、肾脏及红细胞的脂质过氧化物浓度,增强SOD 和CAT 活力,改善镉诱导的氧化应激。此外,黄岩(2017)在牛磺酸改善CCl4诱导斜带石斑鱼原代肝细胞氧化损伤的研究中发现,10 mmol/L 牛磺酸不仅能够显著增强肝细胞活力及抗氧化酶活力,同时上调了CAT和GSH-Px 的基因表达量,从细胞层面初步揭示了牛磺酸抗氧化的分子机制。Yan等(2019)研究发现,全植物蛋白饲料中添加0.99%牛磺酸可提升草鱼幼鱼肠道中抗菌物质的活力和含量,并通过抑制核因子-κB 抑制蛋白激酶(IKKs)/核因子-κB 抑制蛋白α(IκBα)/核转录因子κB(NF-κB)信号通路,缓解嗜水气单胞杆菌感染后引发的炎症反应,强化了肠道免疫屏障功能,降低肠炎发病率。
4.2 提升水产动物抗胁迫应激能力
水产动物应激时的生理变化与体内营养状况紧密关联。牛磺酸除了能够提升水产动物抗氧化能力及免疫力,还能够提升水产动物抗胁迫应激能力。罗志成等(2020)研究了基础饲料中添加牛磺酸对慢性热应激条件下虹鳟血清生化指标的影响,发现预添加1 200 mg/kg牛磺酸组在24 ℃及25 ℃慢性热应激24 h后可显著增强机体抗氧化能力、非特异性免疫能力及抗损伤能力。Zhou 等(2017)研究发现冷应激条件下牛磺酸的添加可有效抑制活性氧自由基的产生及非特异性脂酶(NSE)的活性,并通过增加透明细胞比例、降低半颗粒细胞及颗粒细胞的比例以改善细胞黏附性,进而抑制细胞凋亡。Tan 等(2018)以杂交乌鳢(Channamaculatus♀×Channa argus♂)为试验模型,发现6.0~15.0 g/kg 牛磺酸能提高抗氧化酶活性及热应激蛋白-70(HSP-70)基因表达量,减少活性氧簇(ROS)含量、细胞凋亡比例,有效缓解氨氮胁迫产生的应激损伤,降低死亡率。此外,在对稻田养殖黄鳝(Monopterus albus)的研究中发现,低鱼粉饲料中添加1.5 g/kg 牛磺酸可降低空气暴露产生的应激反应,并提升血清中SOD 活性及总抗氧化能力(T-AOC),并抑制MDA 的产生,有效降低空气暴露引发的氧化损伤。因此,牛磺酸对不同应激源造成的鱼体应激反应及损伤均有良好的改善作用,但需进一步研究确定牛磺酸针对不同种类水产动物及不同种类应激源的适宜剂量和具体调控机制。
5 牛磺酸增强其他营养物质的利用能力
水产动物体内的牛磺酸主要由蛋氨酸等含硫氨基酸进行合成,因此为了使体内更多的蛋氨酸等含硫氨基酸用于蛋白质合成,会在饲料中补充适量蛋氨酸等含硫氨基酸用于牛磺酸合成。Gibson 等(2007)采用3×3 双因素(牛磺酸和蛋氨酸含量均为0、5 g/kg 和10 g/kg)试验明确了当饲料中没有合成蛋氨酸添加时,5 g/kg 牛磺酸组虹鳟的增重率及含肉率显著提升,而当饲料中补充蛋氨酸时各组间虹鳟的增重率及含肉率并未出现显著差异,同时发现牛磺酸和蛋氨酸会对亚牛磺酸、蛋氨酸、胱氨酸及胱硫醚含量、含肉率产生显著的交互作用,表明全植物蛋白饲料中补充牛磺酸是必要的,且蛋氨酸的补充并不会影响水产动物对饲料中牛磺酸的需求,同样的结果在波斯鲟(Acipenser persicus)及欧洲鲈鱼(Dicentrarchuslabrax)的研究中也得到了证实。
补充适宜的牛磺酸有助于机体胆固醇的代谢。而在植物蛋白替代鱼粉条件下,胆固醇作为水产动物必需的营养素,往往需要饲料外源胆固醇进行补充。因此在水产动物上也开展了关于牛磺酸和胆固醇间相互作用的研究。Yun 等(2012)以大菱鲆(Scophthalmus maximusL.)为研究对象探讨牛磺酸和胆固醇对其生长及胆固醇代谢的影响时发现,高植物蛋白饲料中添加1.0%胆固醇、1.0%牛磺酸或者二者的组合可显著提高大菱鲆的生长性能,同时添加牛磺酸和胆固醇有叠加效应;此外,在高胆固醇饲料基础上添加牛磺酸可通过降低大菱鲆肝脏中3-羟基-3 甲基戊二酸单酰CoA 还原酶(HMG-CoAr)mRNA 相对表达量,抑制肝脏中胆固醇合成,进而降低血浆中胆固醇含量。
6 小结与展望
在植物蛋白源替代鱼粉及新型蛋白源开发的行业大背景下,饲料中氨基酸平衡问题及功能性营养物质缺乏问题凸显,不仅会造成水产动物机体产生生理病理变化,引发营养性疾病,还会增加饲料及药物使用量,提高养殖成本。而牛磺酸正是植物蛋白源及某些新型蛋白源易缺乏的功能性氨基酸之一。目前,牛磺酸在水产动物上的营养生理与免疫作用及其应用已有很多研究,但仍需从以下几个方面开展相关研究:①在“缺乏牛磺酸的植物性蛋白源及某些新型蛋白源广泛应用于水产饲料”的大背景下,进一步评估不同种类水产动物牛磺酸的营养需求情况;②从不同生长阶段及食性等角度,比较研究水产动物牛磺酸合成代谢调控机制;③从与激素的协同作用、改善胰岛素等角度更深入探讨牛磺酸调控水产动物糖代谢的潜在机制;④从绿肝综合征和贫血症等疾病入手深入探究牛磺酸防治水产动物营养性疾病的潜在机理。通过对牛磺酸生理功能及作用机制研究的逐渐深入,牛磺酸在水产动物饲料中的应用潜力将被进一步释放,将为水产养殖业的绿色健康发展奠定坚实基础。
