塔里木河上游典型荒漠植物叶片性状及其与土壤因子的关系
2022-07-30李蕊希
李蕊希,吴 雪,*,贡 璐
1 新疆大学生态与环境学院, 乌鲁木齐 830017
2 绿洲生态教育部重点实验室, 乌鲁木齐 830017
3 新疆精河温带荒漠生态系统教育部野外科学观测研究站,精河 833300
叶片作为植物与外界环境接触最为直接的器官,对外界环境变化具有较大的可塑性和敏感性[1]。叶片性状是植物适应环境所体现出的叶片水平上的特征参数,对环境变化能够产生积极的响应,并对其具有重要的指示意义[2]。在干旱或高辐射的影响下,植物可通过改变叶片长度、叶片宽度以及叶面积等结构性状以减小叶片边界层阻力从而实现散热的目的。植物依靠根系从土壤中吸收生长发育所需的养分,并利用叶片进行光合作用来同化和累积有机物,通过不断调节碳、氮、磷等物质元素在体内的代谢与循环维持相对稳定的内环境,从而应对环境的复杂变化。土壤水分和土壤养分是限制和调控植物生存和分布的重要土壤因子,土壤水分的缺失对植物施加了生理约束,从而影响植物个体的生长发育状况[3—6]和植物群落的内部环境变化;土壤碳、氮、磷含量可以影响植物群落结构和生产力,反之植物通过凋落物的输入来影响土壤养分变化[7]。国内外学者通过大量的研究发现,植物在土壤因子影响下最常见的叶片变化包括比叶面积减小[8],叶片厚度增加[9],叶干物质含量增加,叶氮和叶磷含量维持较高水平[10—12]等现象。但目前关于极端干旱地区荒漠植物叶片性状研究相比于其他区域较少,且其与不同土壤因子之间的关系还有待进一步探究。
塔里木河上游地区年均降水量小于50mm,潜在蒸发量高达2000mm,属于典型的降水稀少、蒸发强烈,生态环境脆弱的极端干旱区。植被组成较为简单,主要以耐盐碱、耐干旱、耐风沙的荒漠植物为主,分布于该区域的荒漠植物在形态结构[13—14]、生理代谢[15—16]等方面进化产生一系列独特的响应特征。本研究以塔里木河上游绿洲-荒漠过渡区和荒漠区为研究区,以该区域两个优势种花花柴(Kareliniacaspica)和骆驼刺(Alhagisparsifolia)为研究对象,分析其叶片结构性状和化学性状的差异性,并建立其与土壤因子的相关关系,以揭示荒漠植物适应性生存策略,并确定影响荒漠植物叶片性状的关键土壤因子及其贡献度,以期为极端干旱区荒漠植被的保护和恢复提供参考。
1 材料与方法
1.1 研究区概况
研究区位于塔里木河上游地区的绿洲—荒漠过渡区和荒漠区,隶属新疆生产建设兵团农一师11团和12团境内,地处塔克拉玛干沙漠北缘,天山山脉南麓,海拔约为1000m。研究区属暖温带大陆性干旱气候,干旱少雨,蒸发强烈,年均气温约为12.4℃,年均降水量小于50mm,但年均潜在蒸发量高达2000mm,表现冬季干冷、夏季干热且昼夜温差大等特点。研究区主要的土壤类型为灌漠土、棕漠土、盐碱土和风沙土,植被类型主要为灌木和草本,植被分布较为稀疏。其中,灌木植物主要有柽柳(Tamarixramosissima)、盐穗木(Halostachyscaspica)、盐节木(Halocnemumstrobilace)和白刺(NitrariatangutorumBobr)等,草本植物主要有骆驼刺(Alhagisparsifolia)、花花柴(Kareliniacaspica)、芦苇(Phragmitesaustralis)、猪毛菜(Salsolaarbuscula)等。其中,花花柴是多年生草本植物,叶片厚,叶面积小,叶肉质化明显,具有耐高温、耐干旱、耐盐碱等广谱抗性;骆驼刺根系发达,枝上带大量刺,种皮坚硬、致密,具有较好的耐盐碱、抗旱性,可用来抵制草原退化和防风固沙。两种植物均是喜生于高温荒漠地带的典型荒漠植物代表,也是分布于塔克拉玛干沙漠北缘的优势物种。
1.2 样地选择
2020年7月,对塔里木河上游地区全面踏查的基础上(表1),在绿洲—荒漠过渡区和荒漠区分别选择盐碱地和沙地两种不同生境分别设置3个研究样地,在每一个样地内设置4个2m×2m的样方,以典型荒漠植物花花柴和骆驼刺为研究对象,开展植被调查以及样品采集工作。
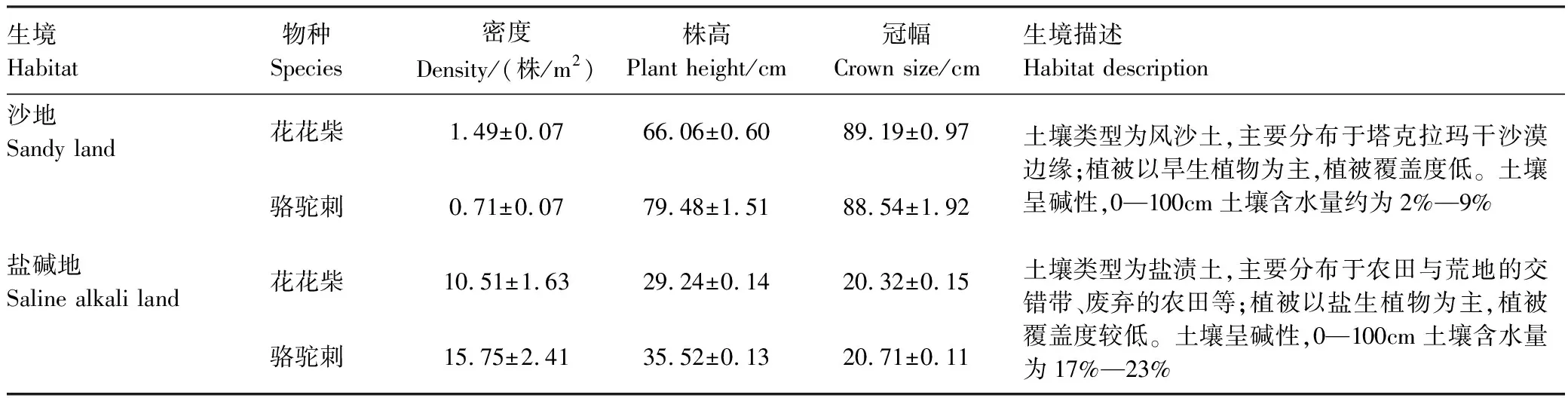
表1 样方基本概况
1.3 样品采集
在每一样方内,对分布在其中的花花柴和骆驼刺进行植被调查,详细记录其数量及株高、冠幅等结构性状。调查完成后,选择形态结构大小相近、生长状况良好的花花柴和骆驼刺植株作为目标对象,分别从东、西、南、北四个方位采集其成熟、健康的叶片若干,及时装入具有编号的牛皮纸信封中,放入低温冰盒保存,带回实验室。
在每一样方的中心位置,去除表层凋落物后,利用土钻采集0—20cm的土壤样品1份,去除植物根系、石头等杂物后装入自封袋中。此外,采集部分土壤装入铝盒以备土壤含水量测定,并通过环刀法采集原状土样以备土壤容重测定。
1.4 指标测定与方法
1.4.1叶片性状指标测定
本研究测定的指标有植物的叶厚度(leaf thickness,LT,cm)、叶面积(leaf area,LA,cm2)、比叶面积(specific leaf area,SLA,cm2/g)、叶组织密度(leaf tissue density,LTD,g/cm3)等结构性状和植物叶有机碳(leaf organic carbon,LOC)、氮(leaf nitrogen,LN)、磷(leaf phosphorus,LP)元素的含量(g/kg)及其元素化学计量比等化学性状。选取完全伸展、无病虫害、成熟且完整的叶片分散平铺在扫描仪(CanoScan LiDE300)上扫描并用ImageJ软件分析可得叶面积,使用游标卡尺对选取的叶片进行叶厚度测量。将采集到的叶片放于105℃烘箱中杀青2小时,随后在80℃条件下烘干至恒重,烘干后的样品先进行称重得到叶干质量,再用粉碎机粉碎过0.15mm筛,装入密封袋保存待测。SLA和LTD通过以下的公式计算得出:
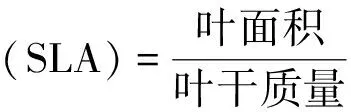
(1)

(2)
LOC含量采用重铬酸钾外加热法测定;LN含量采用凯氏定氮法测定;LP含量采用钼锑抗比色法测定,随后计算得出叶片C/N、C/P、N/P的值。
1.4.2土壤因子测定
本研究测定的土壤因子指标包括容重(bulk density,BD,g/cm3)、土壤含水量(soil water content,SWC,%)、pH值、电导率(electrical conductivity,EC,μs/cm)以及土壤有机碳(soil organic carbon,SOC)、氮(soil nitrogen,SN)、磷(soil phosphorus,SP)元素含量(g/kg)以及土壤有效氮含量(available nitrogen,AN,mg/kg)、土壤有效磷含量(available phosphorus,AP,mg/kg)等。其中BD测定采用环刀法,SWC采用烘干法测定。将采集的表层土壤样品风干、研磨、过筛后,pH值采用电位计法测定;电导率采用电导法测定;SOC含量采用重铬酸钾外加热法测定;SN含量采用凯氏定氮法测定;SP含量采用钼锑抗比色法测定;AN含量采用碱解扩散法测定;AP含量采用碳酸氢钠浸提法测定。
1.5 数据统计分析
用Microsoft Excel 2013、Origin 2018、R、SPSS 23和CANOCO 4.5软件对所得数据进行数据处理、统计分析、绘图。用双因素方差分析(Two-Way ANOVA)和单因素方差分析(One-Way ANOVA)检验不同生境下荒漠植物的叶片性状差异及土壤因子差异。用Pearson相关分析荒漠植物叶片性状各指标间的相关性。借助主成分分析法(Principal Component Analysis,PCA)通过降维对荒漠植物叶片性状进行排序获取主要指标。其中,公因子方差均有较大分值表明在因子信息归类过程中信息损失较小;随后以特征值>1为原则,提取主成分得出特征值和主成分贡献率;依据初始因子的旋转成分矩阵得到各性状指标综合得分,其得分大小表明各个性状指标对主成分的贡献情况;指标因子排名越靠前表明其占据更重要的比重。用冗余排序(Redundancy Analysis,RDA)分析荒漠植物的叶片性状与土壤因子之间的关系,蓝色箭头代表物种,红色箭头代表环境变量;箭头连线长度代表变量所占比重,箭头连线越长表示影响越大;箭头与排序轴的夹角表示物种与环境变量的相关性的大小,夹角越小表明相关性越大。
2 结果分析
2.1 不同生境叶片性状特征
双因素方差分析结果表明,植物叶片LT、LA、SLA、LTD等结构性状在花花柴和骆驼刺两个物种之间具有显著差异(P<0.05)(表2,图1);花花柴和骆驼刺植物叶片LOC、LN、LP、C/N、N/P、C/P等化学性状在沙地和盐碱地等不同生境中均具有显著差异(P<0.05)(表3)。其中,分布在盐碱地的花花柴和骆驼刺的LOC(438.60g/kg,457.50g/kg)、LN(18.58g/kg,17.59g/kg)、N/P(21.08,19.07)和C/P(502.06,499.69)显著高于沙地(P<0.05),而分布在沙地的花花柴和骆驼刺的LP(1.34g/kg,1.33g/kg)和C/N(29.55,35.39)显著高于盐碱地(0.89g/kg,0.92g/kg和23.76,26.50)(P<0.05)(图2)。
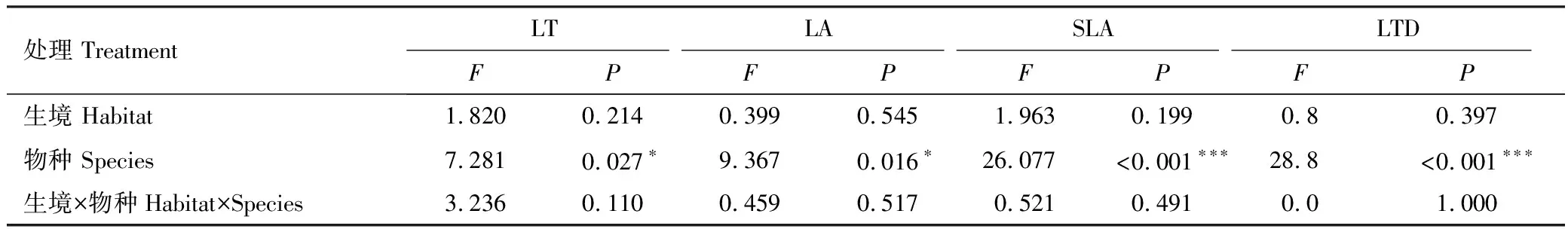
表2 不同生境下植物叶片结构性状之间的差异性
2.2 不同生境叶片性状相关关系
Pearson相关分析结果表明(图3),在两种生境下植物的LA与SLA呈极显著正相关性(P<0.01),而LA、SLA均与LTD呈极显著负相关(P<0.01);LN与C/N、LP与C/P呈极显著负相关(P<0.01)。在沙地中,LOC与LN呈极显著正相关(P<0.01),LA、SLA均与LOC、LN、N/P呈正相关(P<0.05),LTD与C/N呈极显著正相关(P<0.01)。在盐碱地中,LN与LP呈显著正相关(P<0.05),LT、LA、SLA均与LOC呈负相关(P<0.05),LT与C/N、C/P呈极显著负相关(P<0.01)。
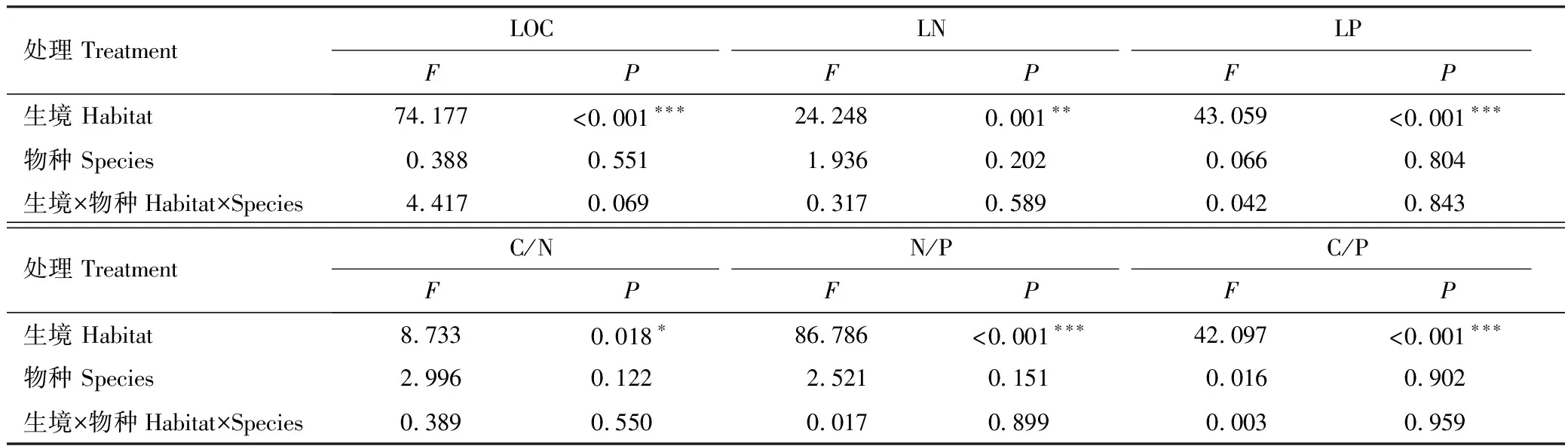
表3 不同生境下植物叶片化学性状之间的差异性
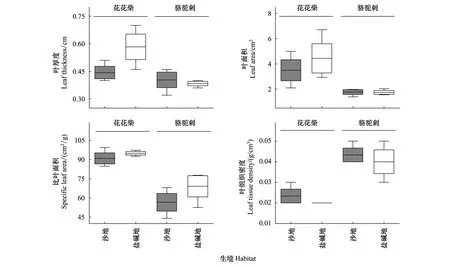
图1 塔里木河上游不同生境中典型荒漠植物叶片结构性状Fig.1 Leaf structure traits of typical desert plants in different habitats in the upper reaches of Tarim River
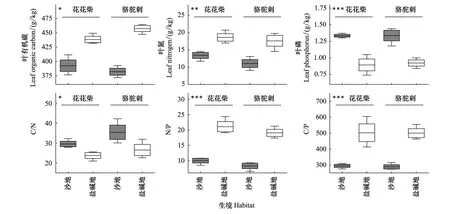
图2 塔里木河上游不同生境中典型荒漠植物叶片化学性状Fig.2 Leaf chemical traits of typical desert plants in different habitats in the upper reaches of Tarim River***P<0.001,**P<0.01,*P<0.05

图3 塔里木河上游不同生境下典型荒漠植物叶片性状相关性系数Fig.3 Correlation coefficient of leaf traits of typical desert plants in different habitats in the upper reaches of Tarim RiverP<0.01**,P<0.05*;LT:叶厚度 leaf thickness;LA:叶面积 leaf area;SLA:比叶面积 specific leaf area;LTD:叶组织密度 leaf tissue density;LOC:叶有机碳含量 leaf organic carbon;LN:叶氮含量 leaf nitrogen;LP:叶磷含量 leaf phosphorus
通过主成分分析可以得出(表4),沙地和盐碱地的具有最小公因子方差的叶片性状指标分别为LOC和LA。沙地的特征值分别为5.019、2.435和1.729,3个主成分贡献率分别为50.190%、24.352%和17.291%,累积贡献率为74.542%和91.833%;盐碱地的特征值分别为4.500,2.982和1.783,3个主成分贡献率分别为44.999%,29.820%和17.835%,累积贡献率为74.819%和92.654%,说明沙地和盐碱地各自的3个主成分因素可以代表荒漠植物叶片性状变化的主要因素。通过对两种生境的综合得分位次进行排名可知,沙地的前三个指标因子为LA、N/P和LN,而盐碱地的前三个指标因子为N/P、SLA、LA。
2.3 不同生境土壤因子特征
通过单因素方差分析结果表明(图4),除BD、SN、AN、SP在盐碱地与沙地间无显著差异外,SWC、pH、EC、SOC和AP等土壤因子在不同生境间均存在显著差异(P<0.05或P<0.01),其中盐碱地的SWC(22.30%)、pH(8.62)、EC(2396.67μs/cm)和SOC(4.03g/kg)是高于沙地(0.55%、8.24、1640.67μs/cm和2.18g/kg),沙地的AP(1.91mg/kg)高于盐碱地的AP(0.87mg/kg)。
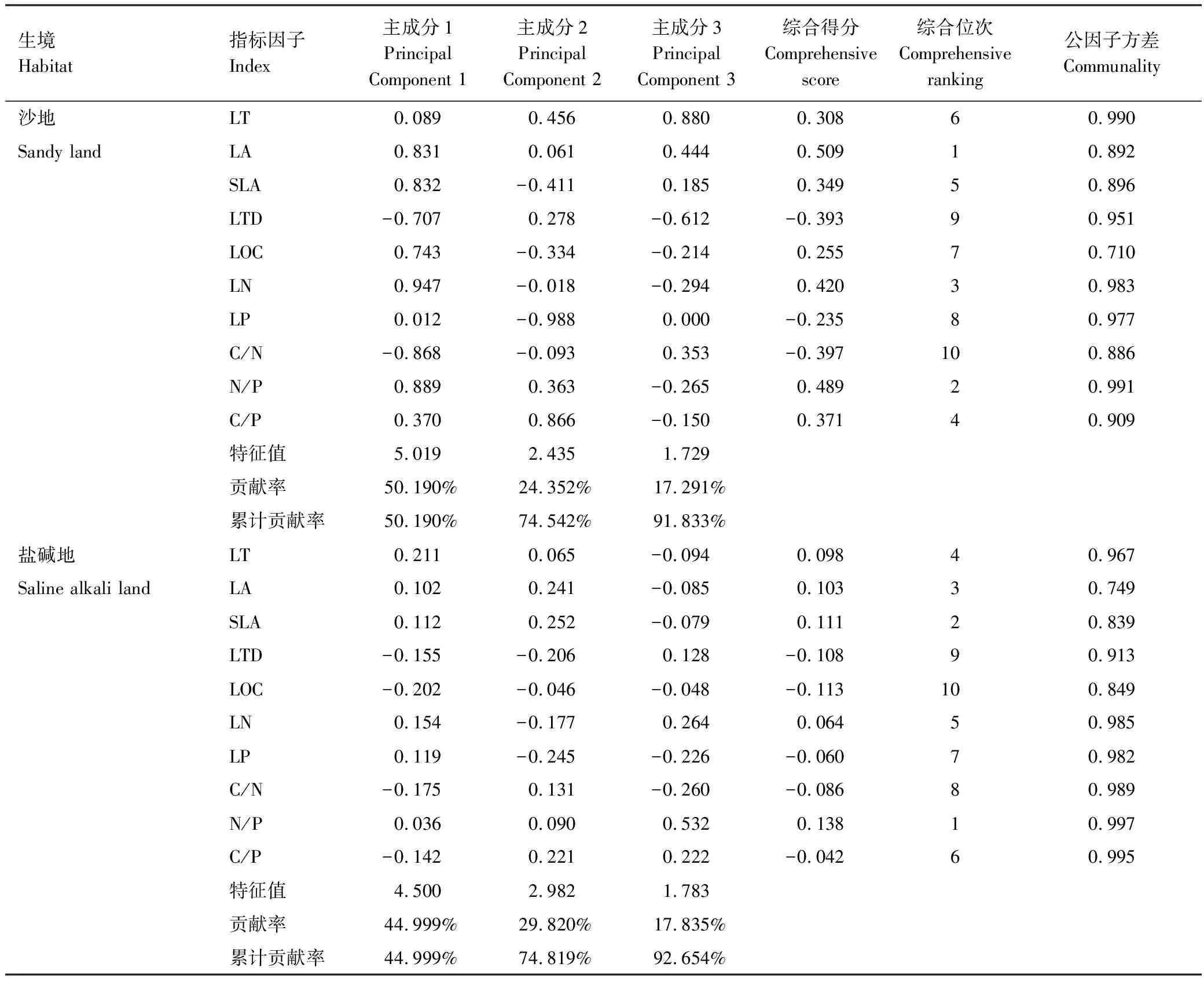
表4 初始因子旋转成份矩阵与主成分贡献率
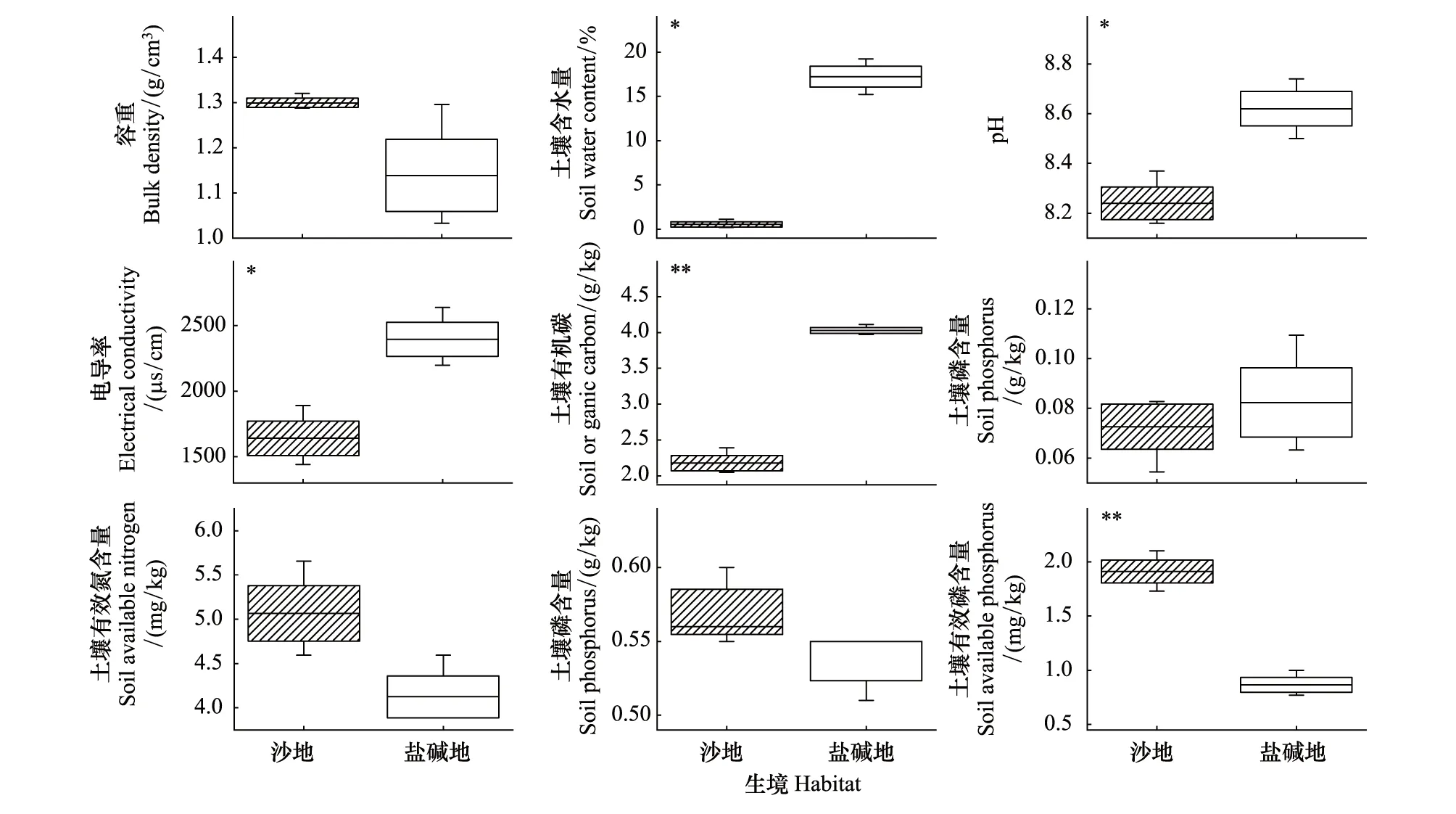
图4 塔里木河上游不同生境土壤因子的差异Fig.4 Difference of soil factors in different habitats in the upper reaches of Tarim River
2.4 叶片性状与土壤因子的关系
通过对2种荒漠植物的叶片性状与不同土壤因子进行RDA排序分析后(图5),结果表明荒漠植物叶片性状特征在第I轴、第II轴的解释量分别为56.5%和6.4%,第III轴、第IV轴的解释量之和仅为3.2%,且前两轴累计解释植物叶片性状特征量为62.9%,对植物叶片性状和土壤因子关系的累计解释量为95.1%。由此可知,前两轴能够很好地反映植物叶片性状和土壤因子关系,并且主要由第Ⅰ轴决定。结果表明,土壤因子中的SWC、SOC和SN箭头连线最长,说明SWC、SOC和SN对荒漠植物叶片性状变异起到了较好的解释。SWC和SOC与LTD、LP和C/N成负相关关系,与LA、SLA、LOC、LN、N/P和C/P成正相关关系,其中SOC与N/P的正相关性最大,SWC与LN的正相关性最大。
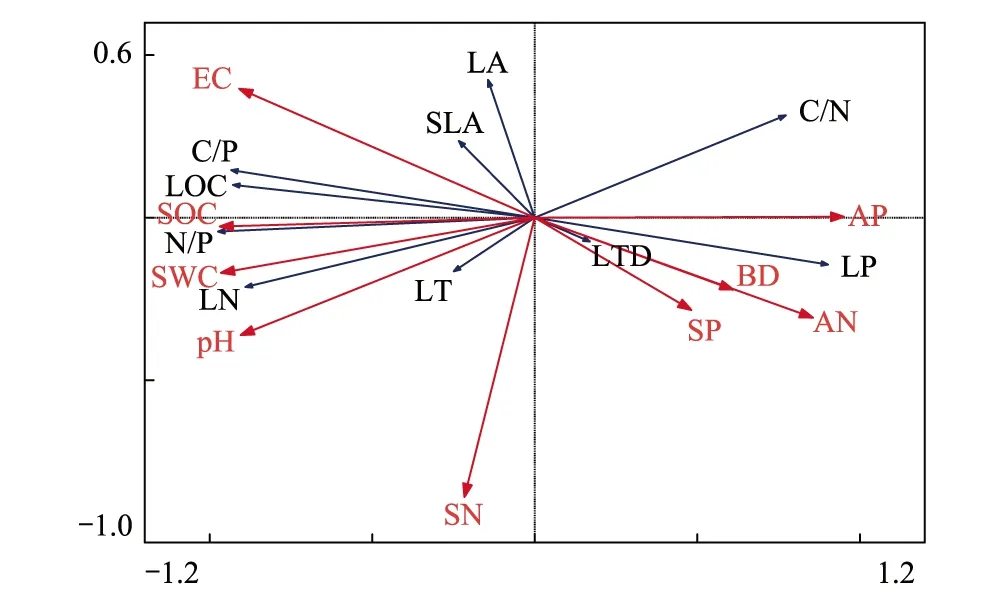
图5 塔里木河上游典型荒漠植物叶片性状与土壤因子的RDA排序Fig.5 RDA sorting of leaf traits and soil factors of typical desert plants in the upper reaches of Tarim RiverBD:容重 bulk density;SWC:土壤含水量 soil water content;EC:电导率 electrical conductivity;SOC:土壤有机碳含量 soil organic carbon;SN:土壤氮含量 soil nitrogen;AN:土壤有效氮含量 available nitrogen;SP:土壤磷含量 soil phosphorus;AP:土壤有效磷含量 available phosphorus
3 讨论
3.1 不同生境植物叶片性状的变化规律
在全球气候变化背景下,植物通过改变其性状做出响应,尤其以植物叶片对环境变化的响应最为敏感[17]。植物LT和LTD可表征植物自我保护机制和养分分配与保持能力[18—21],SLA则与植物对贫瘠和干旱环境的适应性有关[22]。本研究发现,花花柴和骆驼刺叶片LT、LA、SLA和LTD在盐碱地和沙地两种不同生境中差异不显著,但在不同物种间具有显著差异。花花柴的LT、LA和SLA高于骆驼刺,但LTD明显低于骆驼刺。这说明骆驼刺为降低自身叶片失水、抵御高温伤害,可能采取通过减少自身LA、提高LTD来减缓植物体生长速率的策略,而花花柴则通过增加LT、LA和SLA来提高自身的生长速率,获取生存空间。
叶片C、N、P含量及其比值在一定程度上能反映植物的营养状况、植物对营养物质的吸收能力以及植物所处生境C积累动态和N、P养分限制格局[17,23]。在长期自然选择过程中,生长在不同生境中的植物可通过优化自身资源配置以应对环境压力,植物叶片所含化学元素存在差异是表现之一[24]。研究区两种荒漠植物叶片C、N、P含量平均值为417.67g/kg、15.14g/kg、1.12g/kg,这与孙力等[25]对塔里木盆地北缘荒漠典型植物、Yang等[26]对温带沙漠典型植被、温晨等[27]对半干旱黄土典型植被的研究结果较为相近,但低于全球平均水平(464g/kg、18.3g/kg、1.80g/kg)[28]。这可能是由于研究区生态环境较恶劣,植物对养分的吸收和利用速率减慢,因此叶片所能积累的有效养分含量降低,造成植物叶片C、N、P含量较低[29]。此外,本研究中盐碱地植物叶片C、N含量高于沙地,但叶片P含量低于沙地。这可能是由于盐碱地土壤SOC含量较高,因此分布在盐碱地生境的植物吸收积累的C较多;但盐碱地和沙地土壤N含量差异不大,分布在盐碱地生境植物叶片N含量却显著高于沙地,说明生长在盐碱地生境的荒漠植物从土壤中获取N的能力相对要优于生长在沙地生境中植物。已有研究表明,当叶片N/P大于16,植物生长对P元素添加的响应强烈;当叶片N/P小于14,植物生长对N元素添加的响应强烈;叶片N/P在14和16之间被认为则同时受N、P两种元素影响[30]。本研究中,沙地植物叶片N/P远小于14,说明植物生长对N元素的响应更为强烈;而盐碱地植物叶片N/P大于16,表明植物生长更容易受到P元素限制。研究区叶片C/N和C/P平均值均高于全球陆地系统植物平均水平(25.4,257.8)[28]。C/N和C/P可以反映植物的生长速度,一般具有较高的C/N和C/P的植物具有相对较低的生长速率[31]。研究区属于极端干旱区,气温高、蒸发强烈,植物为抵御恶劣环境,降低植物生长速率,从而导致叶片C/N和C/P高于全球陆地系统植物。
3.2 不同生境植物叶片性状相关性及主成分分析
在自然环境中能长期生存并繁衍的植物,既需要具有适应环境变化的能力,还需要具备获取与生长和繁殖有关资源的能力,如光、水和养分等,这必须通过植物多种性状间的协同与权衡实现[32]。因此,植物叶片性状间关系密切,并且这种关系可能会随着生境及物种变化发生特异性改变[33]。当LA减少时,SLA也随之减小,导致LTD增大,体现荒漠植物通过SLA和LTD共同作用以维持体内水分的平衡[33]。因此,在本研究中不同生境下植物的LA与SLA呈极显著正相关性(P<0.01),而LA、SLA均与LTD呈极显著负相关(P<0.01)。C被称为植物有机体内的“骨架”[34],是构成生物大分子结构的基本元素,且为N、P的协同元素,它们之间一般呈正相关关系[20],这在本研究中也得到了证实,如沙地植物的LOC与LN呈极显著正相关(P<0.01),盐碱地植物的LN与LP呈显著正相关(P<0.05)。此外,沙地植物的LA、SLA与LOC、LN、N/P呈正相关(P<0.05),LTD与C/N呈极显著正相关(P<0.01);且盐碱地植物的LT、LA、SLA与LOC呈负相关(P<0.05),LT与C/N、C/P呈极显著负相关(P<0.01),这与王敬哲等[30]对南方红壤侵蚀区不同植被、张剑等[35]对敦煌阳关湿地芦苇、聂明鹤等[29]对宁夏荒漠草原不同优势植物的研究结果较为一致,说明植物养分对于植物形态结构构建具有一定的作用。
通过对荒漠植物叶片性状进行主成分分析发现,本研究中,LA和N/P是不同生境下植物性状中综合排名前三的共同指标因子,表明LA和N/P是塔里木河下游荒漠植物相对最为重要的叶片性状。LA不仅影响植物接收光资源的程度,还对叶片温度和蒸腾速率起作用进而改变叶片热量和水分平衡[36];而N/P是判断植物营养状况的关键指标,关系到植物个体以及整个植物群落的生产力[30],因此LA和N/P是影响荒漠植物群落结构与性质的重要性状。
3.3 叶片性状与土壤因子的关系
土壤是植物水分和化学元素的重要来源,其水分、养分的分布和变化会直接影响植物的生长发育[37]。根据RDA分析,SWC、SOC和SN土壤因子对荒漠植物叶片性状的影响较大。已有大量研究表明在干旱地区,土壤SWC是影响荒漠植物正常生长发育的主要限制因素,本研究进一步揭示土壤SWC与荒漠植物叶片化学性状关系密切,如土壤SWC与叶片N/P、LOC和C/P等化学性状均呈正相关关系,说明SWC的增加能够促进荒漠植物对N、P的吸收以及对C的同化。这与李玉霖[38]、孙力[25]、张晓龙[4]、马剑英[39]等人在荒漠生态系统研究中所得结果相似,其研究结果表明在水分受限的地区,植物可能通过调控对养分的利用对策以适应水分的制约。除土壤水分外,土壤养分状况及变化也在一定程度上制约着植物体内各种生理生化反应。本研究显示荒漠植物的化学性状比结构性状更易受到土壤养分的影响,相关分析揭示研究区植物LOC、LP分别与土壤SOC、AP呈正相关关系,这与已有研究所得结果基本一致,即植物C、N、P含量与土壤C、N、P含量之间存在线性相关关系[29]。此外,植物N/P与土壤SOC之间也存在显著正相关,说明植物化学元素含量对土壤的响应可以通过生态化学计量比来调节。但是本研究中植物LN与土壤SN之间无显著相关性,这与俞月凤[40]、陶冶[41]等人研究中所得结果相似。这可能是因为研究区群落特征、土壤特性等对植物化学元素含量产生一定影响[42],如研究区气候干燥、风蚀严重,大大减弱了土壤对N的固存,导致土壤养分较为贫瘠,其土壤SN平均含量仅为0.078g/kg,远小于全国平均值(1.88g/kg)[43]。
