氮添加对巴音布鲁克高寒湿地土壤微生物量和酶活性的影响
2022-07-30丛孟菲买迪努尔阿不来孜愚广灵申志博杨再磊朱新萍贾宏涛
胡 洋, 丛孟菲, 陈 末, 买迪努尔·阿不来孜, 愚广灵, 申志博, 杨再磊,3, 朱新萍,3, 贾宏涛,3,*
1 新疆农业大学资源与环境学院, 乌鲁木齐 830052
2 新疆农业大学草业学院, 乌鲁木齐 830052
3 新疆土壤与植物生态过程重点实验室, 乌鲁木齐 830052
目前,通过化石燃料的燃烧、施肥和畜牧业等人为活动,大气氮沉降速率已增加了3—5倍[1],据预测到2050年氮沉降速率将会继续加倍[2]。我国已成为全球第三大氮沉降区,而氮沉降速率的快速增加将对陆地生态系统产生深刻的影响[3]。氮素进入土壤将打破土壤原有氮循环过程,进而影响生态系统结构和功能,并产生一系列生态环境问题[4—5]。氮沉降作为最为重要的全球变化问题之一,同样也深刻地影响着土壤微生物群落结构和代谢。土壤微生物在陆地生态系统物质循环和能量流动中具有重要的地位,土壤微生物量和酶活性是土壤微生物数量和活性的代表[6]。而氮添加会引起土壤微生物量和酶活性的显著变化[7]。
土壤微生物量主要包括微生物生物量碳和微生物生物量氮,其不仅代表参与土壤有机质转化和养分循环的微生物数量,而且也能够反映出养分循环的速率[8]。土壤酶主要来源于土壤微生物、植物根系分泌以及植物残体的分解[9],其可以反映土壤中进行的各种反应的强度和土壤微生物群落活性等[10]。已有研究表明氮添加对土壤微生物量与酶活性有显著影响,但在不同的生态系统土壤微生物量和酶活性对氮添加的响应往往有所不同[11—12]。有研究发现,氮添加对微生物群落具有负面影响[13],然而也有研究呈现相反的结果[14]。也有许多研究认为氮添加对土壤微生物量的影响因湿地类型而异。例如,氮添加对高寒小泊湖湿地土壤中的微生物量产生了促进作用,但是对高寒依克乌兰湿地却有抑制作用[15]。在不同陆地生态系统上,土壤酶活性对氮添加的响应也有所差异。例如,氮添加促进了山西太岳山油松林土壤中脲酶和磷酸酶活性,并降低了人工林中的蔗糖酶活性,但对天然林中的蔗糖酶无影响[16]。在藏北高寒草原土壤上的研究发现,氮添加通过增加土壤中易分解的碳氮资源,降低了碳相关水解酶和氧化酶活性,提高脲酶活性;氮添加提高了植物对磷的需求,增强了磷酸酶活性[17]。因此,尽管已经进行了大量氮添加对土壤微生物量和酶活性影响的研究,但是研究结果还有争议,且对氮添加通过土壤因子调节土壤微生物量和酶活性的直接和间接作用还不清楚。
高寒湿地是水陆相互作用形成的独特生态系统,也是全球变化的敏感区与信息库,对维持高海拔地区生态平衡、养分循环以及生物多样性等方面具有重要意义[18]。而巴音布鲁克高寒湿地作为我国中亚干旱区典型高寒湿地,其在维系我国西北干旱区流域生态环境安全等方面具有重要作用。高寒湿地生态系统相比于其他生态系统更加脆弱,对氮沉降等全球变化最为敏感,而在全球氮沉降快速增加背景下,氮沉降增加势必对高寒湿地土壤微生物量和酶活性产生影响,进而影响生态系统的物质循环和能量转换。为此,本研究以巴音布鲁克高寒湿地作为研究对象,布设野外原位氮添加试验,探讨巴音布鲁克高寒湿地土壤微生物量和酶活性对氮添加的响应特征,解析氮添加对高寒湿地土壤微生物量和酶活性的影响机制,结果可为中亚干旱区高寒湿地适应未来气候变化措施的制定以及高寒湿地可持续利用提供数据参考。
1 材料与方法
1.1 研究区概况
巴音布鲁克高寒湿地位于新疆维吾尔自治区巴音郭楞蒙古自治州和静县境内(82°59′—84°35′ E,42°40′—43°00′ N),属国家级湿地自然保护区,巴音布鲁克高寒湿地是中亚干旱地区一个典型而重要的湿地,是世界自然遗产地,在水量调节、蓄水和维持区域水平衡方面发挥着重要作用。海拔2300—3042 m,年平均气温-4.6℃,极端最高、最低气温分别为28℃与-48.1℃,年均降水量为273 mm,年蒸发量1250 mm,年均相对湿度60%。湿地水源主要来自冰雪融水和地下潜水,地下水埋深0.5—1.0 m[19]。
1.2 试验设计
本试验采用野外原位模拟的方法,基于前期设立的长期定位监测样带,并依据地表水分状况和植被群落将高寒湿地划分为:沼泽(S)、沼泽草甸(SM)和草甸(M)。其中沼泽土壤长期处于淹水状态,优势植被主要为大穗苔草(Carexrhynchophysa);沼泽草甸土壤冬季处于覆雪状态,春季冰雪融化及生长季持续降雨后会出现地表积水现象,在干旱季又会出现无积水现象,优势植被主要为黑花苔草(Carexmelanantha)和冰草(Agropyroncristatum);草甸土壤在冰雪融化及持续降雨时会出现短暂积水,优势植被主要为尖苞薹草(Carexmicroglochin)[20]。于2018年选择植物生长季结束后的10月,使用区组设计的方法在巴音布鲁克高寒湿地保护良好的3种湿地类型内分别布设了3块9 m×9 m的小区,共9块小区。试验开始前每个小区按3个施氮处理设置3个地块(2 m×8 m)。地块中设置1m缓冲区,以防止不同处理间的相互影响,并设置1m高的钢制栅栏以防止动物的干扰。根据巴音布鲁克高寒草原历史氮沉降量(8 kg hm-2a-1)来设置本研究施氮浓度[21],分别为CK(0 kg N hm-2a-1)、N1(8 kg N hm-2a-1)、N2(16 kg N hm-2a-1),每个处理3次重复。将尿素充分溶于蒸馏水中后均匀喷洒于各处理区域内,为消除土壤含水量的差异,对照也喷洒同样量的蒸馏水,共9个处理,3次重复。
1.3 样品采集
于施氮1年后的2019年10月使用土钻,采用五点取样法来采集各处理内0—10 cm的表层土样两份。一份土样装于自封袋再装于冰盒中带回实验室,取新鲜样品测定土壤微生物生物量碳(MBC)、微生物生物量氮(MBN)含量,后将剩余土壤样品平铺于白纸上阴干,阴干后将土壤分别过0.25 mm与1 mm标准筛,过1 mm筛的样品用于测定土壤pH、碱解氮、速效磷、速效钾以及土壤酶活性(包括过氧化氢酶、脲酶、碱性磷酸酶、蔗糖酶和蛋白酶活性),过0.25 mm筛的样品用于测定土壤有机碳、全氮、全磷和全钾含量。另一份土壤置于冰箱中保存备用。
1.4 土壤测定方法
土壤pH以水土比2.5∶1浸提后使用pH计测定;土壤有机碳采用浓硫酸-重铬酸钾外加热法测定,土壤碱解氮采用碱解扩散法测定,土壤速效磷和全磷采用钼锑抗比色法测定,土壤速效钾和全钾使用火焰光度计测定,土壤全氮使用元素分析仪测定[22]。土壤微生物生物量碳采用氯仿熏蒸,浓硫酸-重铬酸钾外加热法测定,微生物生物量氮采用氯仿熏蒸,茚三酮比色法测定[23]。土壤H2O2酶采用高锰酸钾滴定法测定、脲酶采用苯酚-次氯酸钠比色法测定、蔗糖酶采用3,5—二硝基水杨酸比色法测定、蛋白酶采用茚三酮比色法测定、碱性磷酸酶采用磷酸苯二钠比色法测定[24]。土壤微生物商用土壤微生物生物量碳含量与有机碳含量的比值表示[25]。
1.5 数据分析
采用Microsoft Excel 2010对数据进行统计整理、SPSS 26.0对数据进行双因素方差分析,分析各个指标对不同湿地类型、不同浓度氮添加及其两者的交互作用的响应;同时,利用LSD检验比较氮添加和湿地类型下土壤性质的差异(P<0.05)。使用R-corrplot软件包分析土壤基础化学性质与土壤微生物量和酶活性之间的相关性。使用构建结构方程模型(SEM)来解析不同湿地类型及氮添加对土壤微生物量和酶活性的直接和间接影响,首先使用R-vegan软件包实现PCA(主坐标分析)排序,使用PC1(第一主成分)参与SEM,最后采用SPSS Amos 24.0构建、验证并解析其结构方程模型。并使用SPSS Amos 24.0计算土壤因子对微生物量和酶活性的标准化总效应[26]。采用Origin 2018对土壤基础化学性质、微生物量和酶活性进行柱状图的绘制。
2 结果与分析
2.1 氮添加对高寒湿地土壤基础化学性质的影响
如表1所示,氮添加对3种湿地类型土壤pH、速效磷和全磷均无显著影响(P>0.05)。但是,与CK相比,N1处理显著降低了SM区土壤有机碳含量,减低了4.19%,而N2处理又无显著影响(P>0.05)。氮添加处理显著增加了M区土壤有机碳含量,较CK增加了15.95%—19.29%。氮添加处理显著增加了SM区土壤碱解氮含量(增加了18.32%—21.20%),随着氮浓度的增加M区土壤碱解氮含量逐渐增加,其中N1处理较CK显著增加了63.96%,N2处理较CK显著增加了93.30%。N2处理较CK显著增加了SM区土壤速效钾和S区土壤全氮含量,分别增加了12.16%和19.02%。从不同湿地类型间可以看出,S区土壤有机碳、碱解氮、全氮含量较其他2个区高;SM区和M区土壤pH和速效钾含量较高,SM区速效磷和全磷含量较高。
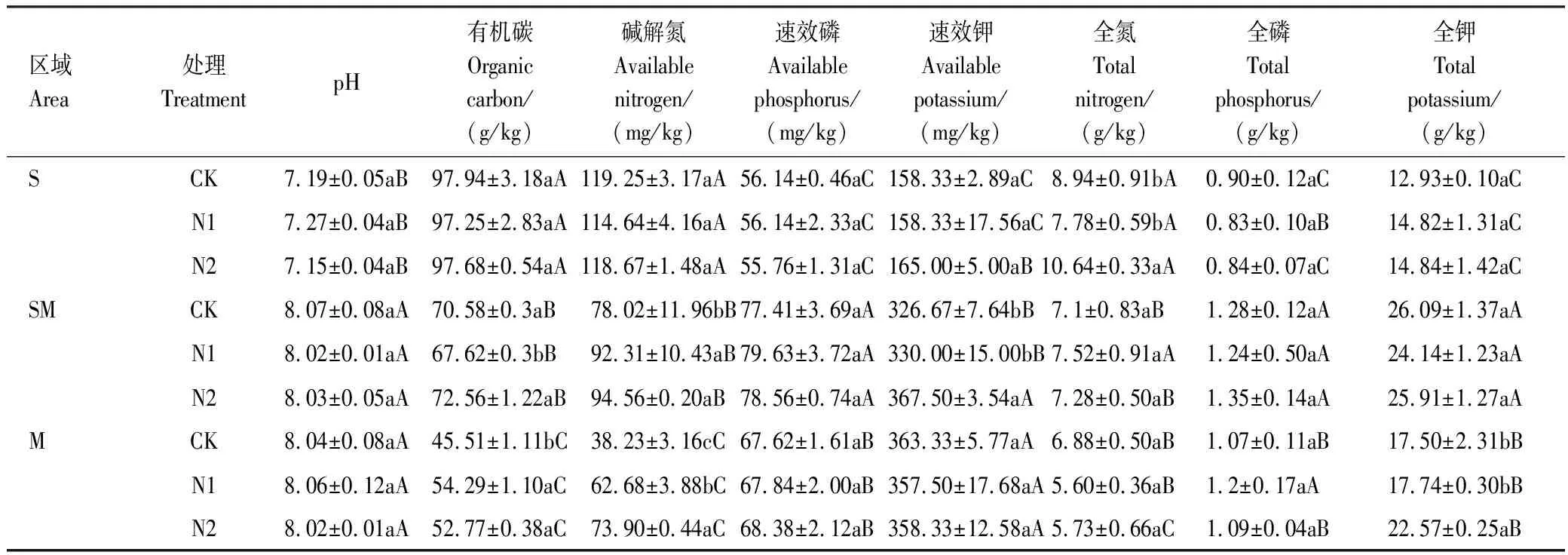
表1 氮添加对高寒湿地土壤基础化学性质的影响
2.2 氮添加对高寒湿地土壤微生物量的影响
氮添加对土壤微生物生物量碳(MBC)、微生物生物量氮(MBN)、微生物生物量碳/氮(MBC/N)和微生物商(QMB)的影响如图1所示,氮添加处理显著增加了3种湿地类型土壤MBC和MBN含量,在S、SM和M区内,土壤MBC分别增加了47.62%—61.90%、42.25%—19.00%、7.00%—20.93%,MBN分别增加了10.79%—18.26%、8.03%—8.15%、18.20%—38.26%。N1、N2处理较CK显著增加了S和SM区的MBC/MBN和QMB,且随着氮添加浓度的增加而逐渐增加。与CK相比,S区MBC/MBN分别增加了24.68%—46.08%,QMB值增加了48.50%—61.92%,SM区的MBC/MBN分别增加了31.72%—102.48%,QMB值分别增加了48.48%—113.10%。而N1、N2处理显著降低了M区的MBC/MBN,降低了8.93%—9.47%,N1处理显著降低了M区的QMB,降低了10.36%,N2处理又显著增加了4.27%。不同湿地类型土壤微生物量也存在显著差异,其中SM区土壤MBC较S区高128.12%,较M区高79.98%。S和SM区MBN含量显著高于M区,分别高出65.54%和67.33%。SM和M区MBC/MBN显著高于S区,分别高出215.78%和172.12%。SM区QMB含量最高,较S区高125.41%,较M区高109.64%。
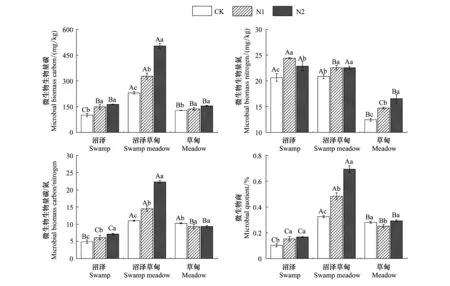
图1 氮添加对高寒湿地土壤微生物量的影响Fig.1 Effects of nitrogen addition on soil microbial biomass in alpine wetlandS:沼泽 Swamp;SM:沼泽草甸 Swamp meadow;M:草甸 Meadow; CK:对照 Control;N1:施氮8 kg N hm-2 a-1处理 Nitrogen application 8 kg N hm-2 a-1 treatment;N2:施氮16 kg N hm-2 a-1处理 Nitrogen application 16 kg N hm-2 a-1 treatment;不同小写字母代表同一湿地类型内不同浓度氮添加处理间差异显著,大写字母表示同一浓度氮添加处理下不同湿地类型间差异显著,P<0.05
2.3 氮添加对高寒湿地土壤酶活性的影响
氮添加对高寒湿地不同湿地类型土壤酶活性有着显著影响(图2)。氮添加显著增加了高寒湿地土壤的H2O2酶活性,在S区内增加了93.31%—157.27%,在SM区内增加了78.26%—81.45%,在M区内增加了34.70%—61.40%。而氮添加对高寒湿地土壤碱性磷酸酶活性无显著影响(P>0.05)。对于蛋白酶、蔗糖酶和脲酶活性而言,在S区内,N1处理对土壤蛋白酶和脲酶活性无显著影响,但是在N2处理下蛋白酶和脲酶活性显著增加,分别增加了7.69%和58.55%。N1处理显著增加了S区土壤蔗糖酶活性,增加了53.58%,N2处理蔗糖酶活性又降低了22.71%。在SM区内,N1、N2处理显著增加了土壤蛋白酶和脲酶活性,蛋白酶增加了7.25%—59.63%,脲酶增加了4.71%—12.78%,对蔗糖酶活性无显著影响(P>0.05)。在M区内,N2处理显著增加了土壤蛋白酶和脲酶活性,分别增加了29.91%和6.58%,N1、N2处理对M区土壤蔗糖酶活性无显著影响(P>0.05)。高寒湿地不同湿地类型土壤酶活性也存在显著差异,其中SM和M区土壤蛋白酶活性较高,较S区分别高出79.84%和64.62%。SM和M区土壤碱性磷酸酶活性较高,较S区分别高出36.85%和21.29%。M区土壤H2O2酶活性活性较高,分别较S区和SM区高41.57%和41.16%。M区土壤脲酶活性较高,分别较S区和SM区高219.74%和8.97%。而3种湿地类型土壤蔗糖酶活性无显著差异(P>0.05)。
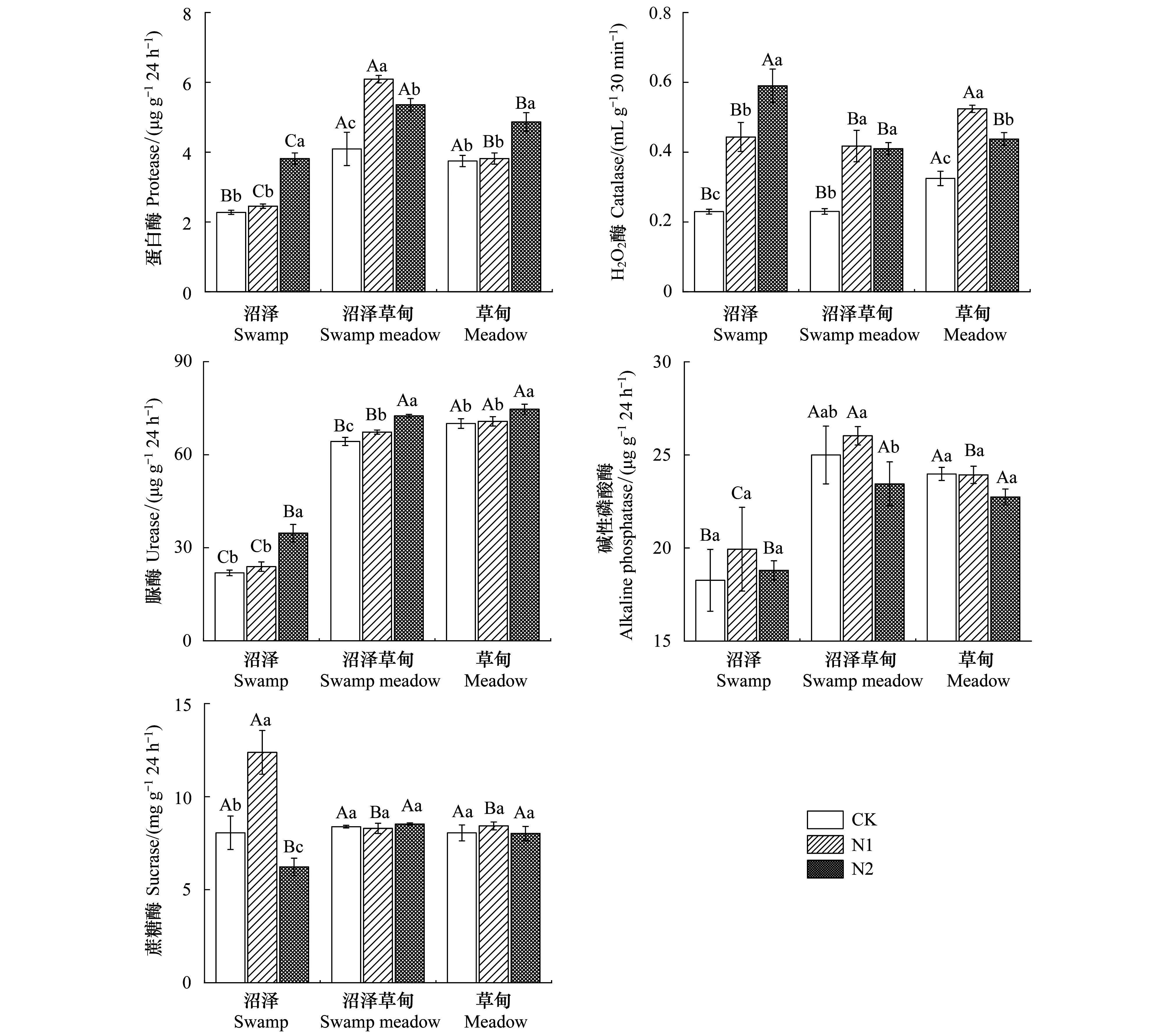
图2 氮添加对高寒湿地土壤酶活性的影响Fig.2 Effects of nitrogen addition on soil enzyme activities in alpine wetland
2.4 氮添加条件下高寒湿地土壤基础化学性质与微生物量和酶活性之间的关系
由图3的相关性分析可知,除土壤蔗糖酶与H2O2酶活性外,其余酶活性和微生物量与土壤基础化学性质之间均存在一定的相关性。其中MBC与速效磷、速效钾、全磷和全钾含量相关(P<0.05),MBN与pH、有机碳、碱解氮、速效钾和全氮相关(P<0.01),而MBC/N和QMB与pH、速效磷、速效钾、全磷和全钾相关(P<0.001)。酶活性中,碱性磷酸酶和脲酶活性与土壤基础理化性质、微生物量之间均有显著相关(P<0.001),蛋白酶与pH、有机碳、速效磷、速效钾、全磷、全钾相关(P<0.01),而蔗糖酶、H2O2酶与基础化学性质之间无显著影响(P>0.05)。
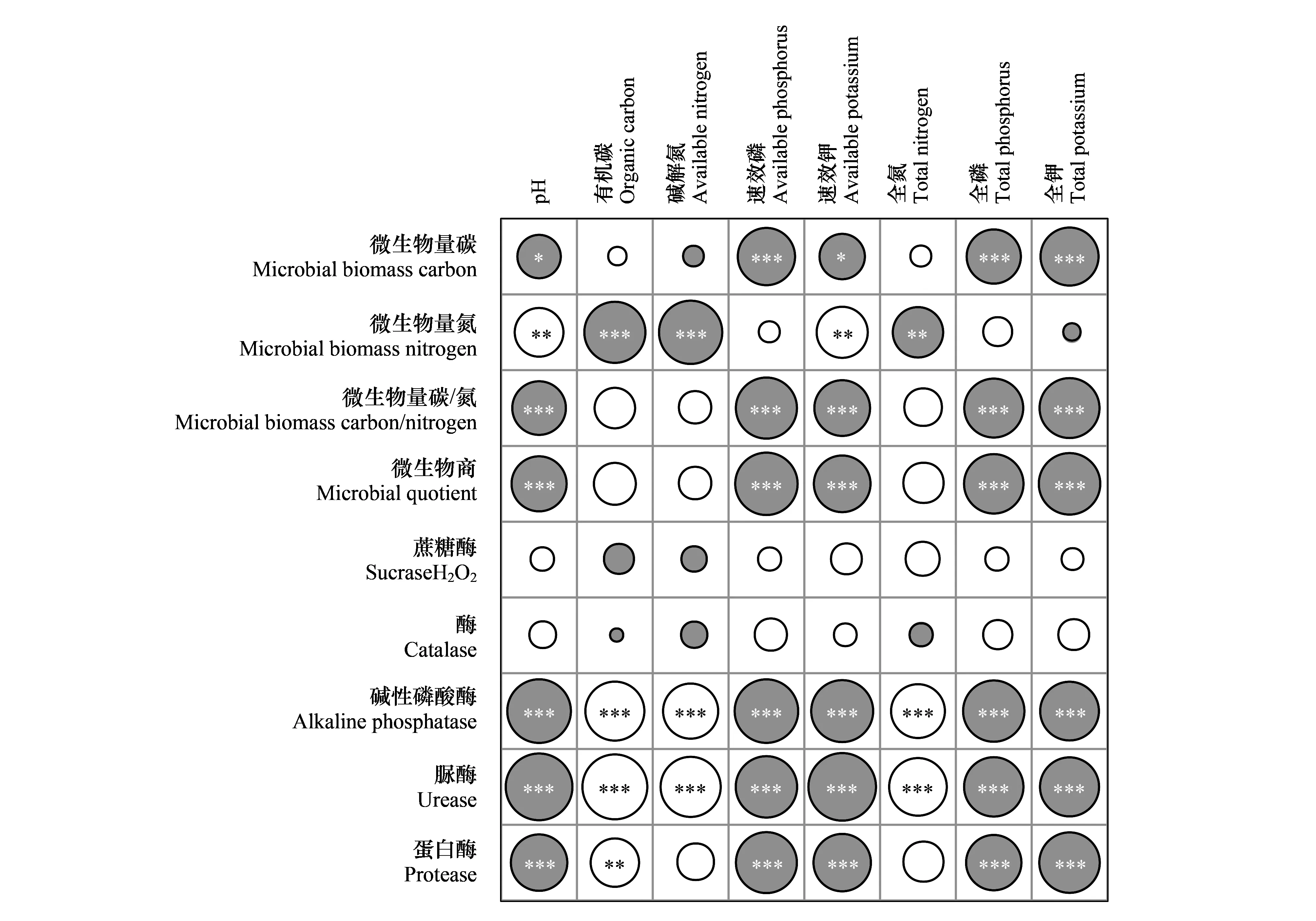
图3 土壤基础化学性质与酶活性、微生物量的相关性矩阵Fig.3 Correlation matrix of soil basic chemical properties with enzyme activities and microbial biomass图中灰色圆圈表示正相关,白色圆圈表示负相关关系,圆圈大小表示相关性R2的大小,圆圈内*表示P<0.05,**表示P<0.01,***表示P<0.001
由表2可知,土壤有机碳、碱解氮、速效钾、全钾、MBC、MBN、MBC/MBN、QMB和土壤蔗糖酶、H2O2酶、碱性磷酸酶、脲酶、蛋白酶活性均在不同施氮水平存在显著差异(P<0.05);且所有指标在不同湿地类型条件下存在显著差异(P<0.05);可以看出湿地类型对土壤化学、微生物量和酶活性的影响大于氮添加的影响。有机碳、碱解氮、速效钾、全钾、MBC、MBN、MBC/MBN、QMB、蔗糖酶、H2O2酶、脲酶和蛋白酶在不同氮添加水平和不同湿地类型的交互作用下存在显著差异(P<0.05)。
使用结构方程模型(SEM)来解析氮添加和湿地类型对土壤酶活性和微生物量的直接和间接影响。用不同湿地类型土壤含水率来表示不同湿地类型。使用主成分分析参与SEM的构建,其中用土壤碱解氮、速效磷、速效钾含量来表示土壤速效养分,PC1解释了83.04%的变异。用土壤全氮、全磷、全钾来表示土壤全量养分,PC1解释了76.33%的变异。用土壤MBC、MBN、MBC/MBN和QMB来表示土壤微生物量,PC1解释了88.77%的变异。用土壤蛋白酶、碱性磷酸酶、蔗糖酶、H2O2酶、脲酶活性来表示土壤酶活性,PC1解释了68.06%的变异。由图4可知,氮添加和湿地类型可直接或间接影响土壤基础化学性质来影响土壤酶活性和微生物量。其中,氮添加直接显著增加了土壤全量养分、土壤酶活性和微生物量,其中对酶活性的影响较大。而随着湿地类型(S-SM-M)的改变,土壤酶活性显著增加,还通过增加土壤有机碳含量降低了土壤酶活性。同时湿地类型的改变还通过降低土壤pH和有效养分,间接增加了土壤微生物量。

表2 不同湿地类型和不同氮添加水平对土壤化学性质、微生物量和酶活性的双因素方差分析
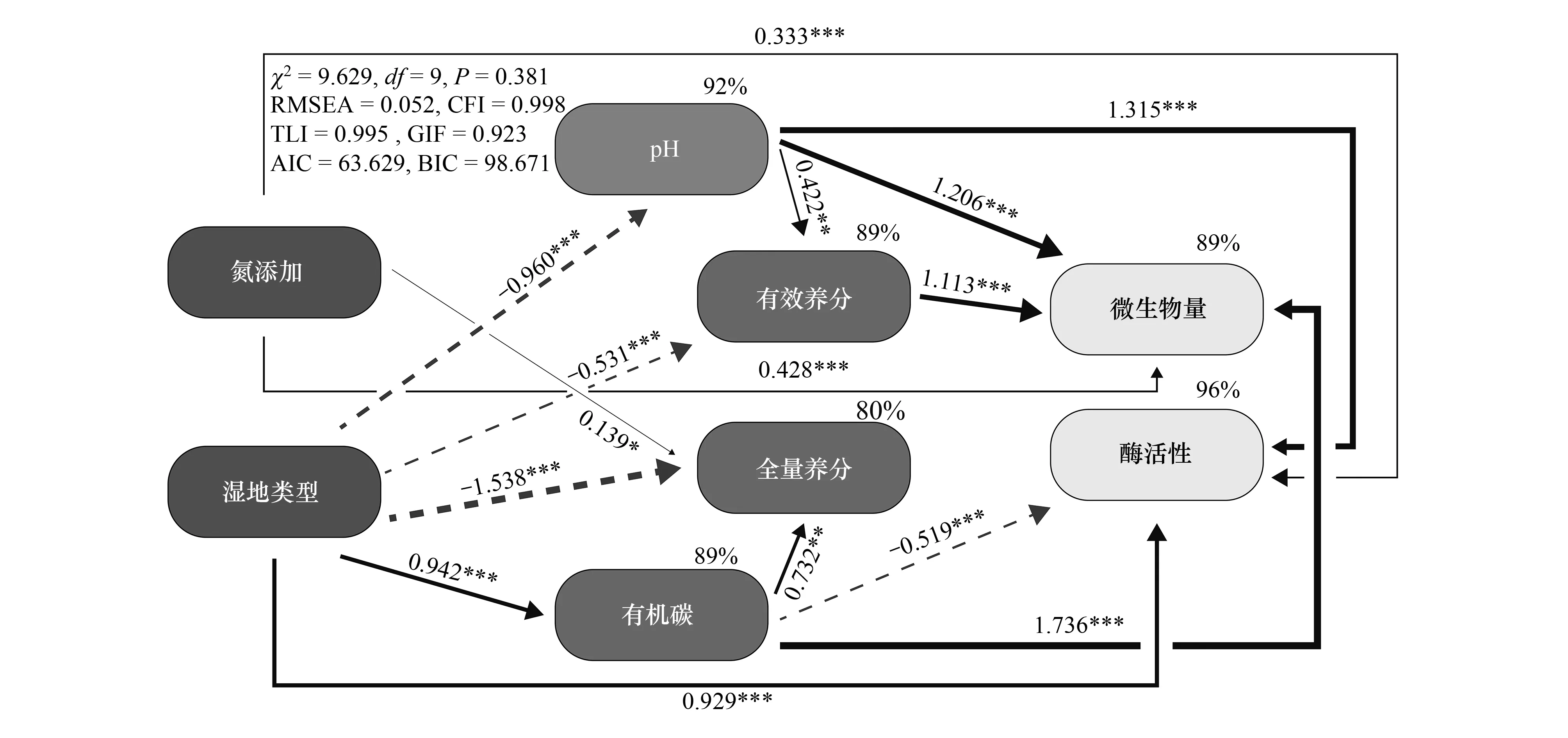
图4 各因子对土壤微生物量和酶活性的结构方程模型分析Fig.4 Structural equation model of soil microbial biomass and enzyme activity for each factor χ2:卡方 Chi-square;df:自由度 Degrees of freedom;RMSEA:近似误差均方根 Root-mean-square error of approximation;CFI:比较拟合指数 Comparative fit index;TLI:Tucker-Lewis指数 Tucker-Lewis coefficient;GIF:拟合优度指数 Goodness-of-fit index;AIC:赤池信息准则指数 Akaike information criterion;BIC:贝叶斯信息准则指数 Bayesian information criterion;图中路径旁边的数字为标准化的路径系数,括号内为对应P值,灰色虚线为负效应,黑色实线为正效应,只有显著的路径被保留,矩形上分百分比为方差解释量(R2)
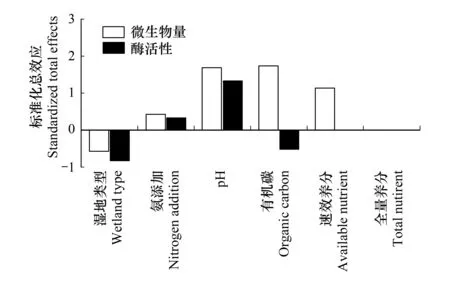
图5 各因子对土壤微生物量和酶活性的影响 Fig.5 Effects of various factors on soil microbial biomass and enzyme activities
由图5可知,pH和有机碳对土壤微生物量的影响较大,而全量养分对微生物量影响的标准化总效应为零。pH对土壤酶活性的影响也较大,而速效和全量养分对酶活性影响的标准化总效应为零。
3 讨论
3.1 氮添加对高寒湿地不同湿地类型土壤微生物量的影响
土壤微生物是土壤生态系统的一个重要组成部分,其参与诸多土壤生态过程,可作为研究和预测陆地生态系统对全球变化响应的对象,但土壤微生物受多种生态因子的影响,氮素又是土壤中最常见的限制性元素,因此氮添加可能会显著影响土壤微生物活性和代谢[27]。土壤微生物量碳(MBC)、生物量氮(MBN)可作为土壤微生物群落活性表征指标,土壤微生物商(QMB)又在一定程度上反应了土壤有机碳的利用效率,所以土壤微生物量和微生物商可作为研究土壤环境变化的敏感性指标[28]。目前有关氮添加对土壤微生物量及微生物商的影响具有很大的不确定性,例如,在长白山森林土壤的研究发现氮添加可显著增加土壤MBC[29]。但是,也有研究发现氮添加显著降低了草原和森林土壤的MBC,增加了MBN含量[30—31]。一项Meta分析发现,氮添加降低了森林和草地土壤的MBC和MBN,而对农田和湿地土壤有着正向影响,这与生物群落、氮类型、实验持续时间和气候因素等都有关系[32]。本研究中,氮添加总体上增加了高寒湿地土壤MBC、MBN、MBC/MBN和QMB值。一方面,可能是由于氮添加缓解了土壤中氮的限制,促进了微生物活性,使微生物量增加[33]。另一方面,可能是氮添加提高地上凋落物和死亡根系的输入量,缓解了土壤微生物对碳的限制[32]。本研究巴音布鲁克高寒湿地沼泽(S)、沼泽草甸(SM)、草甸(M)土壤的微生物量和酶活性有着显著的差异。其中,SM区处于季节性积水状态,土壤水分含量更加适宜微生物生长,所以SM区土壤具有较高的微生物性状。
3.2 氮添加对高寒湿地不同湿地类型土壤酶活性的影响
土壤酶是物质循环和能量流动不可替代的参与者,是土壤生态系统中最活跃的组分之一,其中脲酶、磷酸酶、蔗糖酶等水解酶活性能够表征土壤氮、磷、碳等养分的循环状况,过氧化氢酶活性与土壤腐殖化过程、生物呼吸强度紧密相关,蛋白酶活性与有机碳和氮循环相关[34]。关于氮添加对酶活性影响的研究众多,氮添加会改变土壤环境,影响微生物合成许多重要的酶[35]。在天山云杉林土壤和华西雨屏区光皮桦人工林土壤上的研究发现,氮添加对土壤脲酶、蔗糖酶和磷酸酶活性有促进作用[36]。前人Mate分析发现,氮添加使土壤脲酶活性提高了18.6%[37]。而本研究中,氮添加显著增加了土壤脲酶、蛋白酶和H2O2酶,这是由于氮添加增加了底物浓度刺激了与氮相关酶活性(脲酶和蛋白酶),同时氮添加使土壤有效氮含量增加,促进植物生长,增加凋落物和死亡根系输入量,进而使土壤微生物活性增强,增强了土壤H2O2酶活性[38]。随着施氮浓度的增加,S区土壤蔗糖酶活性呈现先增加后降低的趋势,这可能是由于S区土壤与碳循环相关的微生物受养分添加的响应较为敏感,适量氮素添加刺激微生物活性,而过量则抑制其活性[39]。而氮添加对土壤碱性磷酸酶活性无显著影响,这可能是由于本研究区中磷素充足,施氮未影响微生物分泌磷酸酶以获取磷素[40]。
3.3 氮添加下高寒湿地土壤微生物量和酶活性的影响因素
在陆地生态系统中,土壤微生物受到多种因素的影响,如气候、植被和土壤因子等[41—42]。有研究发现,在氮添加条件下,人工林土壤中SOC、TN和TP的含量及其化学计量比均对土壤微生物群落具有一定的影响[43]。还有研究发现,在森林生态系统上,氮添加条件下,土壤酶活性主要与土层深度、土壤pH以及森林生态系统内的碳氮磷等限制元素显著相关[44]。除氮添加外,不同湿地类型的差异也可以通过植被类型和土壤因子等来改变土壤微生物量和酶活性,但是目前在高寒湿地生态系统上的影响机制还不清楚。在本研究中,不同浓度氮添加和湿地类型的差异均显著影响着土壤微生物量和酶活性,其中氮添加直接影响着土壤酶活性和微生物量。这可能与土壤中氮素的有效性有关[34],氮添加可能提高了土壤净氮的矿化速率使土壤中有效氮含量增加[45],补充了土壤中的氮限制,刺激了微生物活性,促进了微生物生长和代谢。在本研究中,随着湿地类型的改变(沼泽-沼泽草甸-草甸),直接增加了土壤酶活性,还通过增加土壤有机碳含量微弱地间接降低了土壤酶活性。同时,还通过降低土壤pH和有效养分含量来增加土壤中微生物量。这与本研究中不同湿地类型的土壤pH值和有机碳含量的差异有关系,S区由于其常年处于积水状态,土壤处于厌氧环境,植物凋落物和死亡根系不易矿化分解,导致有机碳含量较高,同时不断积累的凋落物和死亡根系进一步腐殖化后形成弱酸性腐殖质,导致pH降低[46—47]。而pH和有机碳又是影响微生物群落的关键土壤因子[48]。与pH和有机碳相同,土壤有效养分也是土壤微生物的一个关键因子。本研究中,SM和M区分别处于季节性和临时积水状态,土壤水分的差异会直接影响养分的运输、扩散和有效性[49]。加之植被群落类型和生长状况的差异,影响植物对养分的吸收,故不同湿地类型土壤有效养分含量有所不同[50]。由此可知,在氮添加条件下,氮添加直接促进了微生物量和酶活性,而在不同湿地类型条件下,土壤pH、有机碳和有效养分的差异是导致土壤微生物量和酶活性变化的主要因素。
4 结论
(1)氮添加和不同湿地类型显著影响着土壤酶活性和微生物量,且其交互作用显著影响着微生物量、蔗糖酶、H2O2酶、脲酶和蛋白酶活性。沼泽草甸土壤的MBC含量较沼泽和草甸土壤平均高出110.79%。沼泽和沼泽草甸土壤的MBN含量较草甸土壤平均高出66.44%。沼泽草甸和草甸土壤蛋白酶活性较沼泽平均高出72.23%,碱性磷酸酶活性高出29.07%。草甸土壤H2O2酶较沼泽草甸和沼泽平均高出41.37%,脲酶活性高出114.36%。
(2)氮添加直接影响了土壤酶活性和微生物量,土壤MBC显著增加了7.00%—119.00%,MBN增加了8.03%—38.26%。脲酶、蛋白酶和H2O2酶活性分别增加了7.25%—59.63%、4.71%—58.55%和34.70%—157.27%。N1处理增加了沼泽土壤蔗糖酶活性(增加了58.58%),N2处理显著降低了22.72%。氮添加对沼泽草甸和草甸土壤蔗糖酶活性无显著影响。
(3)湿地类型可直接或间接(pH)影响土壤酶活性,并可通过pH、有机碳和有效养分间接增加土壤微生物量。
