应用脂肪酸组成研究热带东太平洋同域中上层鲨鱼营养生态位分化
2022-07-30李云凯
李云凯,徐 敏,贡 艺,4,*
1 上海海洋大学海洋科学学院, 上海 201306
2 农业农村部远洋与极地渔业创新重点实验室, 上海 200009
3 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室, 青岛 200071
4 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306
探究有限资源环境中物种多样性维持及物种共存机制是群落生态学的核心问题[1]。各物种特征的权衡及组合的不同决定了其生活史对策的不同,由此决定了各物种在群落中生态位的差异化[2]。其中,基于物种对食物资源利用差异的营养生态位分化是物种共存的必要条件之一[3]。
大洋中上层鲨鱼多处于食物网的顶端或近顶端,其可通过下行效应(top-down control)对食物网结构和功能起调控作用[4],但对同域分布中上层鲨鱼的物种多样性维持及生态共存机制仍不明晰,尤其在资源相对贫瘠的开阔大洋。热带东太平洋海域主要栖息有8种中大型中上层鲨鱼物种,包括大青鲨(Prionaceglauca)、镰状真鲨(Carcharhinusfalciformis)、长鳍真鲨(C.longimanus)、大眼长尾鲨(Alopiassuperciliosus)、浅海长尾鲨(A.pelagicus)、尖吻鲭鲨(Isurusoxyrinchus)、路氏双髻鲨(Sphyrnalewini)和锤头双髻鲨(S.zygaena)[5],为东太平洋的顶级捕食者。对其营养生态位的比较研究有助于揭示大洋同域分布顶级捕食者的共存机制。
脂肪酸组成分析是近年来在生态学领域兴起的一项新方法,在海洋生物研究领域,该方法已开始应用于头足类[2]和硬骨鱼类[6]的营养生态位研究,而对鲨鱼的研究较少。作为顶级捕食者,鲨鱼自身合成的脂肪酸极少,且主要为短链和去饱和度较低的种类[7],而其摄入的外源性脂肪酸通常以保守形态沉积于组织中,可整合一段时间内的摄食信息[8]。其中某些脂肪酸在生物的食性和营养关系研究中具有指示作用,如捕食者机体内Σ22+Σ24的含量可追溯其陆源植物食性,DHA/EPA(C22:6n3/C20:5n3)含量比可用于指示其营养级[9]。因此,脂肪酸组成可用于反映鲨鱼与其食物间的动态关系和营养流动[10],进而用于探究鲨鱼的营养生态位。Bierwagen等通过肌肉和血浆脂肪酸组成分析研究了澳大利亚东北海域两种同域礁鲨——灰三齿鲨(Triaenodonobesus)和钝吻真鲨(Carcharhinusamblyrhynchos)的资源分配及利用,结果表明,两种鲨鱼存在激烈的资源竞争;灰三齿鲨营养生态位较小,表现为其食性的特化[11]。本研究通过测定热带东太平洋8种中上层鲨鱼肌肉的脂肪酸组成,分析其食性差异、营养关系及营养生态位分化,探讨其资源分配方式及同域共存机制。
1 材料与方法
1.1 样品采集

图1 热带东太平洋鲨鱼样品捕获地点 Fig.1 Sampling location of pelagic sharks in the tropical Eastern Pacific
鲨鱼样本采集自2014年6月—11月我国金枪鱼延绳钓兼捕渔获。采样范围为8°—10°N,115°—125°W的东太平洋海域(图1)。共采集到鲨鱼72尾,具体生物学信息见表1。取鱼体臀鳍附近白肌样本,真空包装低温运至实验室,于-20 ℃冰箱冷冻保藏。
1.2 脂肪酸分析与数据处理
肌肉样品去除表皮,使用超纯水漂洗后放入冷冻干燥机(Christ Alpha 1- 4)-55 ℃干燥24 h,用冷冻混合球磨仪(Mixer mill MM440)磨碎成粉末(直径5 μm)。称取样品粉末200 mg于具塞离心管中,加入12 mL二氯甲烷-甲醇溶液(2/1,体积分数),室温下浸泡24 h。离心后取上清液,加入4 mL 0.9 %的氯化钠溶液洗涤,静置至溶液明显分层。收集下层二氯甲烷层至圆底烧瓶中,用氮气(氮吹仪)吹扫除去有机试剂后,加入4 mL氢氧化钠—甲醇溶液(0.5 moL/L)复溶,获得肌肉总脂提取液。
将盛有总脂提取液的圆底烧瓶连接水浴回流装置加热20 min后,加入4 mL三氟化硼-甲醇溶液(14%,w/v),在60 ℃水浴中进行甲酯化反应20 min,反应结束冷却至室温,再加入4 mL正己烷,震荡30 s后,加入饱和氯化钠溶液定容。正己烷层移入配有聚四氟乙烯瓶盖的后壁玻璃管。脂肪酸采用气相色谱/质谱联用仪(7890B/5977A,Agilent)进行分析。色谱条件:毛细管柱型号为AgilentHP- 88(60 m×0.25 nm×0.20 μm),载气为高纯氦气,分流比为10:1,进样口温度为250 ℃。升温程序:初始温度为125 ℃,以8 ℃/min升温至145 ℃并保持26 min,然后以2 ℃/min升温至220 ℃并保持1 min,最后以1 ℃/min升温至227 ℃并保持1 min。
以37种脂肪酸混标及内标十九烷酸甲酯标品作为标准,通过比对保留时间对脂肪酸进行定性分析,采用内标法(GLC 37,Nu-Chek Prep Inc,Elysian)对脂肪酸进行定量分析
其中,Xi是待测样品中组分i的含量(mg/g);Ai是组分i的峰面积;m是样品质量;m19是内标物十九烷酸甲酯的质量;A19是待测样品中内标物的峰面积;Fi是组分i与内标物的校正因子之比,称为相对校正因子[12—13]。
计算各脂肪酸占总脂肪酸含量的百分比,整体分析时去除小于0.5%的数据[14]。

表1 鲨鱼采样信息及基础生物学参数
1.3 数据分析
鲨鱼脂肪酸组成测定结果以“平均值±标准偏差”(mean±SD)表示,采用单因素方差分析(analysis of variance, ANOVA)检验各脂肪酸含量是否存在显著种间差异。聚类分析(cluster analysis)用以分析数据之间的相似性,明确划分成组。主成分分析(principal component analysis,PCA)可用于探究多元事物的主要影响因素,通过降维将多个相互关联的数值转化成少数互不相关的指标。因此本研究进一步应用PCA将脂肪酸测定结果降至二维,并利用R语言中的SIAR软件包计算标准椭圆面积(SEAc),以表征各鲨鱼物种的营养生态位[15]。绘图使用Origin 2021 pro和R软件4.0.2操作。
2 结果与分析
2.1 脂肪酸组成
8种鲨鱼肌肉中共检测得脂肪酸35种,含量大于0.5%的共21种(表2)。饱和脂肪酸含量范围为39.29%—48.64%,种间差异显著(P<0.05),主要种类为C14:0、C16:0、C17:0、C18:0、C20:0、C22:0、C23:0和C24:0。其中,C16:0和C18:0含量最高,分别为13.22%—25.18%和15.22%—28.52%。两种饱和脂肪酸含量存在显著种间差异(P<0.05)。

表2 鲨鱼肌肉脂肪酸组成/%
单不饱和脂肪酸含量范围为8.75%—15.08%,种间差异显著(P<0.05),主要种类为C16:1n7、C18:1n9和C24:1n9。其中,C18:1n9含量较高,为3.11%—5.93%。长鳍真鲨肌肉中C18:1n9含量最高,锤头双髻鲨最低,种间差异显著(P<0.05)。
多不饱和脂肪酸含量范围为39.46%—49.38%,种间差异显著(P<0.05),主要种类为C18:2n6、C20:2、C20:3n3、C20:3n6、C20:4n6、C22:2n6、C20:5n3(EPA)和C22:6n3(DHA)。其中,C20:3n3、C20:4n6、C20:5n3和C22:6n3含量较高,分别为3.95%—9.13%、3.96%—9.20%、0.64%—3.34%和22.32%—33.89%,锤头双髻鲨肌肉中C20:3n3和C20:4n6含量最高,大青鲨肌肉中C20:5n3含量最高,浅海长尾鲨肌肉中C22:6n3含量最高,此4种脂肪酸含量种间差异显著(P<0.05)。此外,∑22+∑24含量范围为27.35%—38.38%,种间差异显著(P<0.05),大青鲨低于其余7种鲨鱼。鲨鱼肌肉中DHA/EPA含量的比值,尖吻鲭鲨(50.68)>浅海长尾鲨(33.72)>大眼长尾鲨(33.12)>镰状真鲨(21.34)>锤头双髻鲨(15.21)>路氏双髻鲨(14.05)>长鳍真鲨(9.76)>大青鲨(8.38)。
2.2 聚类分析
为进一步探究8种鲨鱼的脂肪酸组成特性关系,依据含量大于0.5%的21个脂肪酸指标进行的聚类分析结果显示(图2),树状图在距离8—10内分成两组,第一组为大眼长尾鲨、浅海长尾鲨和尖吻鲭鲨,属鼠鲨目;第二组为大青鲨、镰状真鲨、长鳍真鲨、路氏双髻鲨和锤头双髻鲨,属真鲨目,其中,大青鲨、镰状真鲨和长鳍真鲨属真鲨科,路氏双髻鲨和锤头双髻鲨属双髻鲨科。同一组鲨鱼在聚类树状图上的欧氏距离相距较短,表明其肌肉脂肪酸组成较为相似。

图2 聚类分析图Fig.2 Cluster analysis of fatty acid profilesBSH:大青鲨 blue shark;SPL:路氏双髻鲨 scalloped hammerhead shark;SPZ:锤头双髻鲨 smooth hammerhead shark;FAL:镰状真鲨 silky shark;OCS:长鳍真鲨 oceanic whitetip shark;BTH:大眼长尾鲨 bigeye thresher shark;PTH:浅海长尾鲨 pelagic thresher shark;SMA:尖吻鲭鲨 shortfin mako shark
2.3 主成分分析
由主成分散点图可知(图3),3种鼠鲨散布在相同区域,表明其脂肪酸组成相似。但3种鼠鲨与5种真鲨基本散布在不同区域,表明其脂肪酸组成存在差异。负荷图可反映造成差异的主要脂肪酸种类。依据负荷图,3种鼠鲨肌肉中C16:0、C16:1n7和C22:6n3含量更高,而5种真鲨C18:0、C20:4n6、C24:1n9和C20:5n3含量更高。主成分分析结果与聚类分析结果相似。营养生态位宽度及重叠见表3,大青鲨和镰状真鲨占据较大的营养生态位宽度,尖吻鲭鲨和路氏双髻鲨占据的营养生态位宽度较小。3种鼠鲨与5种真鲨营养生态位存在明显分离或有较小重叠。大眼长尾鲨和浅海长尾鲨营养生态位重叠程度较高。
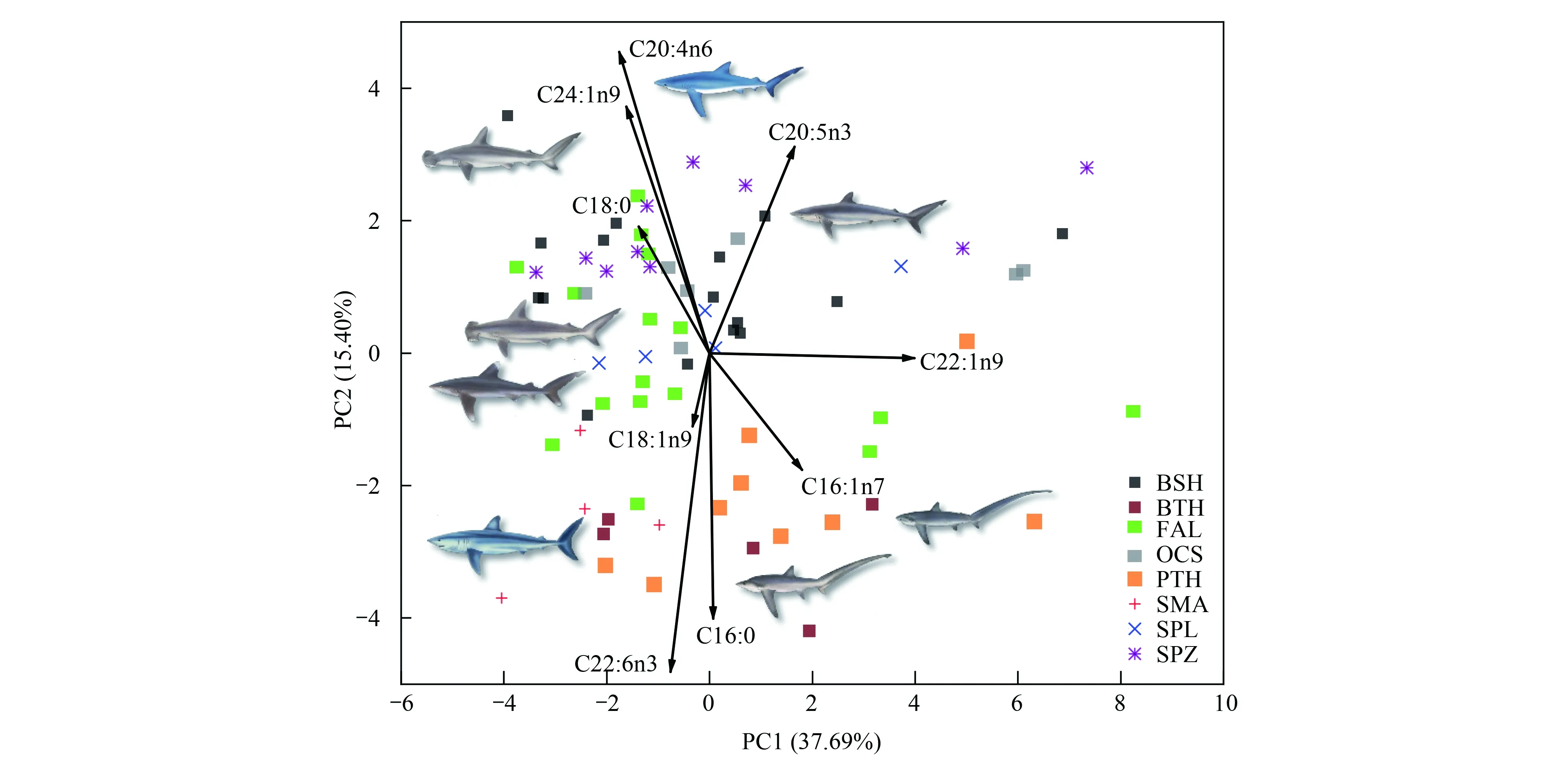
图3 主成分分析图Fig.3 Principal component analysis of fatty acid profiles
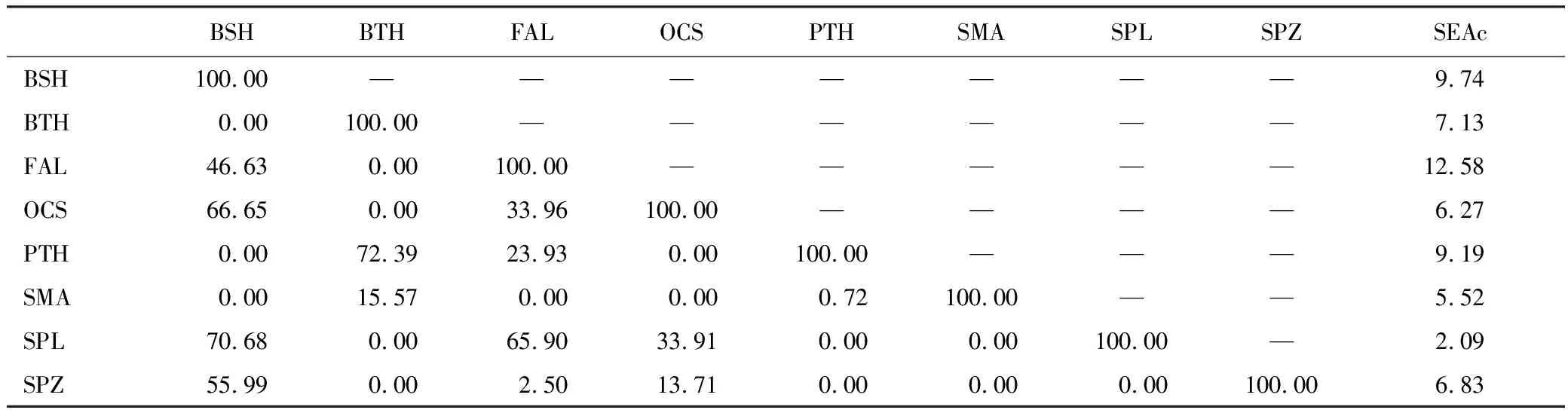
表3 东太平洋8种鲨鱼肌肉营养生态位面积(SEAc)及重叠面积百分比/%
3 讨论
本研究以东太平洋海域8种中上层鲨鱼为研究对象,通过分析肌肉组织脂肪酸组成探讨其营养生态位分化。8种鲨鱼肌肉中饱和脂肪酸和多不饱和脂肪酸占比较大,而单不饱和脂肪酸占比较低。这与Cárdenas-Palomo等对加勒比海鲸鲨(Rhincodontypus)及Davidson等对南非东海岸9种大型鲨鱼的研究结果一致[16—17]。鲨鱼多为顶级捕食者,其自身合成的脂肪酸种类较少,主要有C14:0、C16:0和C18:0以及它们在Δ9去饱和酶作用下产生的C14:1n5、C16:1n7和C18:1n9脂肪酸[7]。必需脂肪酸(essential fatty acid)如C20:5n3、C22:6n3和C20:4n6等仅能由初级生产者或细菌合成,并沿食物链保守性储存于捕食者体内,可用于指示鲨鱼的摄食信息[10]。其中,C20:5n3主要由硅藻自身合成,且在头足类体内含量较高[18—19],本研究中大青鲨和镰状真鲨肌肉中C20:5n3含量较高,表明这2种鲨鱼所处食物链的初级生产者可能为硅藻,且偏好捕食头足类,这与2种鲨鱼的胃含物分析结果一致[5]。C22:6n3是甲藻的特征脂肪酸,也可指示肉食性程度[20—21],路尖吻鲭鲨,浅海长尾鲨和大眼长尾鲨肌肉中较高含量的C22:6n3表明其可能使用由甲藻主导的营养途径且肉食性程度较高。路氏双髻鲨肌肉中20:4n6含量较高,而C20:4n6多由双壳类和甲壳类选择性的保留在体内,可用于指示捕食者对甲壳类的摄食[22]。Flores-Martínez等研究发现,路氏双髻鲨存在垂直运动模式,捕食底栖硬骨鱼类和甲壳类[23]。Σ22+Σ24含量可一定程度上反映陆源植物食性[24],大青鲨肌肉中Σ22+Σ24含量远低于其余7种鲨鱼,说明其可能因终生的大洋性生活史特征,受陆源因素影响较小。研究发现,鱼类肌肉中DHA/EPA比值与营养级成正比[20]。本研究中尖吻鲭鲨比值最高,为50.68,而大青鲨仅为8.38。Silveira等利用稳定同位素技术分析了加利福利亚西南海域5种鲨鱼的营养级,发现尖吻鲭鲨营养级最高,而大青鲨则相对较低[25]。
营养生态位重叠反映了物种间对资源利用的相似程度及物种间潜在的竞争关系,而营养生态位分化可表征种间食性差异及栖息地分化[10]。浅海长尾鲨和大眼长尾鲨营养生态位高度重叠,反映了浅海长尾鲨和大眼长尾鲨可能存在较为激烈的资源竞争关系[26]。种间摄食空间隔离是缓解食物资源竞争的有效方法[3],大眼长尾鲨奇网(rete mirabile)发达,可维持颅内温度[27],较浅海长尾鲨更适应在深海捕食。尖吻鲭鲨、大眼长尾鲨和浅海长尾鲨与其他5种鲨鱼营养生态位存在明显分离或仅有较小重叠,表明其种间食物资源及栖息地利用的竞争程度较弱。三种鼠鲨目鲨鱼中,尖吻鲭鲨属温血性(endothermic)鲨鱼,体温可较环境温度高达7℃,捕食的环境适应性更强,区域更广[28]。胃含物分析也同样发现,尖吻鲭鲨和大眼长尾鲨可在深层捕食硬骨鱼类,镰状真鲨和大青鲨则主要捕食中上层头足类[29]。
生态位宽度反映了生物对食物资源和栖息地利用的竞争能力[10]。大青鲨和镰状真鲨占据较大的营养生态位宽度,表征大青鲨和镰状真鲨对食物资源和栖息地利用的竞争能力较强,食性相对泛化[26]。Estupián-Montao等发现,1—2龄的大青鲨游向近岸觅食,而3—6龄大青鲨更倾向于远洋觅食,对食物资源的利用范围相对较广[30]。胃含物分析表明,大青鲨主要以头足类、硬骨鱼类和甲壳类为食[5],这也一定程度上解释了大青鲨营养级相对较低的原因。Flores-Martínez等研究显示,镰状真鲨的摄食存在性别间差异并在生长发育过程中存在食性转化,雌性镰状真鲨幼鲨喜好捕食密斑刺鲀(Diodonhystrix),雄性喜好捕食茎柔鱼(Dosidicusgigas),而成年雄性捕食巴鲣(Euthynnusaffinis)[23],本研究中的镰状真鲨同时存在幼体和成体样本,可能对结果有一定的影响。尖吻鲭鲨营养生态位宽度较窄,是由于本研究中的尖吻鲭鲨个体较其他鲨鱼相对较小且均未达到性成熟,可能对食物资源的需求较为单一,胃含物分析也显示,尖吻鲭鲨幼鲨喜好捕食硬骨鱼类,如剑鱼(Xiphiasgladius)和颌针鱼(Belonebelone)等[5,31]。
综上所述,本研究量化分析了东太平洋8种中上层鲨鱼的资源分配方式,探讨了其同域共存机制。其中,3种鼠鲨与5种真鲨存在食性差异或栖息地隔离。大青鲨与镰状真鲨生态位宽度较大,表征其对环境的可塑性较强,而尖吻鲭鲨和路氏双髻鲨生态位宽度较小,表现为其食性的特化。尽管脂肪酸组成分析为研究种间资源利用模式,探究同域物种共存机制提供了新的方法。但鱼类生理差异(个体大小、游动速度及体内关键代谢酶活性等)可影响外源性脂肪酸在其体内的同化程度,进而影响脂肪酸组成[32—33]。目前在摄食生态学中,脂肪酸组成分析仍主要起定性或半定量的辅助功能,未来应优化脂肪酸组成定量分析方法,结合稳定同位素和胃含物DNA条形码技术从多维度完善大洋性鲨鱼摄食生态研究内容,揭示这一特殊捕食群体的共存机制。
