传统豆制品腐败菌污染及抑制研究进展
2022-07-02陈红丽张鑫平黄雨洋朱秀清
曲 敏, 陈红丽, 王 宇, 张鑫平, 黄雨洋, 朱秀清
(哈尔滨商业大学 食品工程学院/黑龙江省普通高校食品科学与工程重点实验室/黑龙江省谷物食品与谷物资源综合加工重点实验室, 哈尔滨 150028)
传统豆制品包括豆浆、豆花、豆腐、豆干和豆腐皮等。 我国有着悠久的豆制品食用历史,近年来随着居民生活水平的提高,我国大豆制品消费量迅速增长。 据中国豆制品专业委员会报道,在2019年豆制品生产行业前50 强企业的投豆量达到了266 万t,2021年的投豆量约达317 万t。 在传统豆制品中,产量最大、影响最广的莫过于豆腐。 虽然不同豆制品的制作工艺与产品特点不同,但蛋白含量与水分含量高的产品特点为微生物提供了理想的繁殖条件,极易引起腐败变质,如发生胀袋、变酸、变臭、变软、变黏等。 如葡萄糖酸内酯豆腐在夏季的保质期只有1 ~2 d,冬季(0 ~4 ℃)不超过7 d[1];汪立平等[2]将未开封的豆浆在37 ℃放置约7 d,发现豆浆盒鼓起,产生酸败味;欧杰等[3]将白干、薄百叶于25 ℃贮藏3 d 后,产酸变黏。 腐败变质问题直接影响了传统豆制品的品质与食用安全。 鉴于传统豆制品尤其是鲜食豆制品保质期短、运输储存均需低温冷藏,而且豆制品生产企业多以生产基地为中心,向四周圆形辐射销售,其辐射距离与豆制品的保鲜时间和企业的运输能力呈正相关,严重影响了豆制品加工企业的产量和规模。 近年来,传统豆制品的腐败变质及腐败菌污染引起了国内外学者们的关注,确定腐败菌的来源和种类,分析不同腐败菌与豆制品品质变化的相关性,并采用安全有效的方法对其进行抑制,日益成为研究热点。 本文对引起传统豆制品腐败变质的原因、腐败菌的种类与腐败特点及抑制机理和方法进行了综述,并对控制腐败菌、延长保质期提出了建议。
1 引起豆制品腐败的原因
对豆制品造成污染的微生物主要有3 种来源途径:原材料自身、生产加工过程以及贮藏条件等。
1.1 原材料自身的影响
1.1.1 种植环境的影响
在大豆种植过程中,土壤中的微生物有可能是引起豆制品变质的腐败菌来源之一。 这些土壤中的微生物在大豆种植期间会受到相关因素的影响,比如发育期、降水量以及轮作的影响等。
大豆在种植前期、分枝期、结荚期、成熟期时,其根际土壤微生物数量存在着动态变化的规律,根际微生物总数由高到低依次为:细菌、放线菌、真菌。 细菌在种植前期数量最高,真菌在分枝期总数最多,放线菌总数在结荚期达到最大值,根际微生物数量在成熟期降至最低[4]。 降水量是决定细菌和真菌群落多样性和组成的主要环境因素之一[5],同时降水量增多促进了土壤有机质的转化,进而促进产生更多的细菌——真菌营养级相互作用。 如南方地区的降水量显著高于北方地区,导致南方地区大豆根际微生物共发生网络比在北方地区复杂,具有更多的细菌——真菌连接[6]。 大豆轮作后,根际微生物的细菌群落结构的多样性更加高。 根际土壤中细菌的操作分类单元(operational taxonomic units, OTU)数增加了6.85 倍,Ace指数增加了4.15 倍,Chao1 和Shannon 指数分别增加了5.45 倍和2.43 倍。 富集了假单胞菌属(Pseudomonas)、葡萄球菌属(Staphylococcus)、根瘤菌属(Rhizobium)、芽孢杆菌属(Bacillus)等。 土壤休眠孢子含量相对减少了77%,根瘤菌(Rhizobium)的丰度增加[7]。 研究表明:在不同产地、不同种植环境中采集的不同品种大豆,细菌总数和芽孢数总集中在一定范围内,细菌总数都集中在6 ~7 lg(CFU/g),芽孢总数集中在1 ~2 lg(CFU/g)[8]。柳玉[8]采集了来自黑龙江地区不同种植环境的大豆,发现枯草芽孢杆菌(Bacillus subtilis)和蜡样芽孢杆菌(Bacillus cereus)是原料中携带的豆制品的固定致腐菌。 根据蛋白电泳分析,大豆中的芽孢杆菌(Bacillus)与大豆生长和收获过程中的微环境存在较大关系,与大豆产地和品种无关。
1.1.2 种皮微生物与内源微生物的影响
渡边笃二[9]利用扫描电镜观察到大豆种皮表面具有大量坑凹,其凹陷深度延伸至表皮栅栏细胞的底部,形成气孔。 另外,大豆籽粒具有多层结构。 微生物可能栖息于气孔及籽粒多层结构的不同部位。 如细菌主要存在于大豆籽粒种皮的外表面以及气孔中,其中外表面细菌总数约为6.72 lg(CFU/g),气孔中的细菌总数约为4.32 lg(CFU/g)[8]。 研究发现,细菌在大豆表皮的柱状细胞上附着力较强,能引起豆制品质构变软、气味酸臭等的耐热芽孢杆菌(Bacillus)和肠球菌(Enterococcus)[2],如枯草芽孢杆菌(Bacillus subtilis)和屎肠球菌(Enterococcus faecium)等主要存在于大豆的外表面和表皮中,且简单水洗和脱皮不能将其除去,这为豆制品生产中微生物的去除增加了困难。因此,研究微生物在大豆中的分布部位,结合特性及其与豆制品的品质劣变和腐败的关系变得日益迫切起来。
内生菌存在于大豆、豌豆和苜蓿等豆科植物的很多组织器官中。 王玉霞等[10]从在不同地点采集的大豆的各种组织中分离得到了137 株内生细菌,发现其中2 株为芽孢杆菌(Bacillus)。 Kuklinsky等[11]从两个不同大豆品种中分离到了内生细菌和附生细菌。 部分分离株经16S rDNA 序列分析鉴定,其中的绝大多数为假单胞菌科(Pseudomonaceae)、伯克霍尔菌科(Burkholderiacea)和肠杆菌科(Enterobacteriaceae)。 这些分离株与分离季节、大豆生长期及获得分离物的组织相关的细菌种群密度方面存在着显著的差异。 Tuitemwong 等[12]的研究表明假单胞菌(Pseudomonas)和肠杆菌(Enterobacter)是使豆腐腐败变质的腐败菌,会使豆腐产生胺臭味、表面出现黄色素和变酸。
1.2 生产过程的影响
原料预处理的影响。 我国豆制品厂用的大豆原料都是从地里收割、没有经过任何特殊处理便投入生产,会沾有泥土、粪便等污染物,因此残留着大量的微生物,故原料大豆一次清洗、二次清洗及滤浆过程中细菌总数变化很小[8]。 石彦国等[13]发现大豆原料中所携带的一部分微生物通过简单的水洗和脱皮并不能将其除去,仍附着在大豆表皮的柱状细胞上。 无论利用干法或是湿法脱皮,交叉污染都会发生,很难起到减少大豆中微生物的目的。 柳玉等[8]发现,浸泡过程中细菌总数增加0.43 ~1.10 lg(CFU/g)。
加热过程中的残留。 在大豆加工过程中,加热不仅能使蛋白质适度变性、使抗营养因子失活,还能杀死生豆浆中大量的微生物。 但过热会使豆浆黏度增加,不利于与凝固剂混合,影响其硬度。 因此,在豆制品的加工过程中,一般采用较为温和的加热煮浆方式,但革兰氏阳性芽孢杆菌和耐热菌会残留下。同时,某些芽孢杆菌在逆环境中会产生残留的耐热芽孢,待到环境适宜,重新萌发,产生毒素,食品安全遭到威胁。 例如枯草芽孢杆菌(Bacillus subtilis)可产生淀粉酶及枯草杆菌素,引起食品腐败变质,蜡样芽胞杆菌(Bacillus cereus)可引起食物中毒。 柳玉[8]发现大豆经加热煮浆过程细菌总数会较大幅度的降低,但在37 ℃保存时,残留细菌得以迅速繁殖,在24 h 后即达到8.14 ~9.07 lg(CFU/g)。 这些加热后仍残留的微生物将随后续加工步骤进入到最终产品,直接影响豆制品的卫生安全和保质期。 因此,原料中的初始细菌数量和加热方式直接影响大豆制品的食用安全和货架期。
加工辅料带入的安全隐患。 豆腐加工过程中,需要加入内酯、石膏、盐卤等凝固剂,空气和土壤中也存在着乳酸菌(lactic acid bacteria)和浮游芽孢杆菌(Bacillus)等,由于生产和保藏不当,极易在生产中将杂菌引入。 Fei 等[14]利用高通量测序技术对传统豆腐凝固剂——天然发酵豆腐乳清(naturally fermented tofu whey,NFTW)微生物多样性进行了分析,结果表明乳酸杆菌(Lactobacillus)占比95.31%,是NFTW 微生物群落中的优势属。 石彦国等[13]利用紫外线对豆腐中的凝固剂进行杀菌处理。 发现与未杀菌组对比,杀菌组豆浆中的细菌总数减少了近一半,说明对凝固剂进行杀菌处理可以有效控制豆腐中的微生物增长。
设备清洗不彻底等。 生产结束后,设备清洗不够彻底使表面仍附着有大量微生物,某些设备与豆浆、豆腐直接接触,营养物质丰富,微生物迅速繁殖,造成产品质量劣变。 吴丽樱[15]发现磨浆机和装豆浆的容器等清洗不彻底,易形成富含蛋白质和多糖的环境。 磨浆时,大豆中的内源微生物芽孢杆菌(Bacillus)会释放出来,在此环境下繁殖迅速,在容器壁上形成菌膜,污染豆腐。
1.3 贮藏条件的影响
在不同的贮藏条件下,复杂食品体系中的微生物之间会通过互惠共生、群体感应或者拮抗作用等相互影响,腐败菌代谢会产黏液、产酸、产腐败味等,导致颜色、质构、气味等显现出腐败特性。 如果某种细菌具有较强竞争力并且能适应此种环境,它就会大量生长繁殖成为优势菌,其他菌生长受到抑制。吴丽樱等[16]研究了内酯豆腐和简易包装老豆腐的腐败菌群变化。 结果显示,引起内酯豆腐腐败变质的主要是耐热的芽孢杆菌属(Bacillus),而简易包装的老豆腐初始菌相构成复杂,在4、25、37 ℃等不同温度储藏腐败后,与初始菌相差异明显,4 ℃下老豆腐腐败菌主要有假单胞菌(Pseudomonas)、芽孢杆菌(Bacillus)。 25、37 ℃储藏腐败后明串珠菌属(Leuconostoc)在数量上占据绝对优势。 李博等[1]研究了37 ℃下不同贮藏时间(0 d 和1 d)下的葡萄糖酸内酯豆腐中优势菌群的变化。 发现新鲜的贮藏0 d 的豆腐中的主要微生物是坚强芽孢杆菌(Bacillus firmus),在37 ℃存放1 d 的样品中优势菌群则以屎肠球菌(Enterococcus faecium)为主,可以说尽管初始芽孢杆菌(Bacillus)在新鲜豆腐中数量占优势,但相同条件下屎肠球菌(Enterococcus faecium)生长繁殖速度大于芽孢杆菌,故成为导致豆腐迅速发生腐败变质的腐败菌。
2 引起传统豆制品腐败的菌种类型
引起传统豆制品腐败的细菌种类及腐败特点见表1[17-28]。 腐败菌代谢导致蛋白质的分解,使豆制品产黏液、产酸,导致其颜色变红或变黄、质构特性指标下降,出现难闻的酸臭味等腐败特性。 在化学指标上,有人利用有机酸分析预测豆腐新鲜度[29]:酸度变化大,腐败程度越大。 在传统豆制品腐败菌的分离鉴定中,多用将传统的表型鉴定与现代分子遗传学鉴定相结合的方法来分离鉴定腐败菌,这有利于全面评估微生物多样性和动态[30]。
引起豆制品腐败的微生物主要有细菌和真菌。从表1 中可以看出,引起腐败变质的细菌主要有芽孢杆菌(Bacillus)、假单胞菌(Pseudomonas)、大肠杆菌(Escherichia coli)、沙门氏菌(Salmonella)、乳杆菌属(Lactobacillus)及金黄色葡萄球菌(Staphylococcus aureus)等。 其中,蜡样芽胞杆菌(Bacillus cereus)、大肠杆菌(Escherichia coli)、沙门氏菌(Salmonella)和金黄色葡萄球菌(Staphylococcus aureus)等为常见的致病微生物,会导致食源性疾病的暴发。 芽孢杆菌(Bacillus)主要作用于豆腐、豆浆和豆腐干,会使豆制品变酸、变臭、质构变软、产生黏液和酸臭味;假单胞菌(Pseudomonas)主要作用于豆腐,会产生黄色色素或荧光素,有胺臭味(腥臭味);大肠杆菌(Escherichia coli)主要出现在生产包装消毒不彻底、卫生条件差的豆制品中,会产酸、产气;乳杆菌(Lactobacillus)主要作用于豆腐和豆浆,会使其酸度增高;金黄色葡萄球菌(Staphylococcus aureus)主要出现在豆腐中,数量超标会致病。

表1 引起传统豆制品腐败的细菌种类及腐败特点Tab.1 Types of bacteria that cause spoilage of traditional soy products and their spoilage characteristics
引起腐败变质的真菌主要包括霉菌和酵母菌。韩翠萍等[31]发现来源于生产设备中的克假丝酵母(Candida krusei)会使豆腐发黏、胀袋以及质构变软。 欧杰等[3]发现诞沫假丝酵母(Candida zeylanoides)、季也蒙毕赤酵母(Pichia guilliermondii)、皮状丝孢酵母(Trichosporon cutaneum)、芸苔丝孢酵母(Trichosporon brassicae)是存在于传统豆制品白干与薄百叶中的腐败菌,会导致豆制品发生腐败。 其他一些真菌,如圆弧青霉(Penicillium cyclopium)和黄绿青霉(Penicillium toxicarum Miyake)等也在腐败变质的豆制品中被检出。
3 传统豆制品腐败菌的抑制机理与方法
3.1 腐败菌抑制的机理
当前,对腐败菌的抑制主要采用物理杀菌、化学抑菌和生物防腐技术,归纳总结腐败菌的抑制机理见图1。 由图1 可知,通过破坏细菌细胞壁的重要组成成分——肽聚糖,干扰细胞壁的合成,使菌体结构受损;通过损伤细菌细胞膜,使其穿孔,膜通透性和流动性得到增加,从而破坏细胞膜结构,氨基酸、离子等诸多内容物渗岀,最终使细胞失活;抑制细菌中大分子物质——核酸和蛋白质合成。 抑菌剂可与DNA 结合或作用于与DNA 合成有关的酶类,使细菌中相关基因的正常合成、表达受阻,抑制蛋白质合成,影响微生物生长代谢所需要的酶,从而抑制或杀死腐败菌;大分子量的抑菌剂可以变成一种生物絮凝剂,吸附在细胞壁表面,形成致密高分子膜,使细菌困在一个相对密闭的环境中,阻碍营养物质吸收,扰乱细菌的新陈代谢,实现抑菌的效果;抑菌剂与腐败菌竞争营养物质,使得运输到细菌内部的营养物质减少,机体的正常代谢紊乱,微生物生长受到抑制。
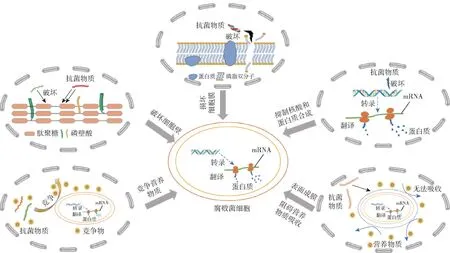
图1 传统豆制品中腐败菌增殖的抑制机理Fig.1 Mechanism of inhibiting proliferation of spoilage bacteria in traditional soy products
3.2 腐败菌抑制的方法
3.2.1 物理杀菌
传统的物理杀菌技术主要有低温保藏、辐照杀菌、超高压保鲜和超高温瞬时杀菌等。
利用低温保藏技术(4 ~10 ℃),抑制微生物的生长及代谢活动。 可较好的保持豆制品的原有风味,但耗能多、成本高,豆腐即使在冷藏条件下也很容易被微生物污染,致使低温难以有效控制耐低温的病原微生物;利用辐照杀菌技术可有效损害细菌细胞内的遗传物质DNA,使其化学键裂解不能继续复制。 是一种杀菌效果好、对食品质量影响不大,能有效延长食品保质期的冷杀菌技术。 目前,被用来进行食品杀菌和提高可储存性的是60Co 和137Cs 发射的γ 辐射技术。 徐远芳等[32]将湘氏休闲豆干放在不同吸收剂量的60Co γ 射线下照射。 发现8.6 kGy 的吸收剂量可以将细菌和霉菌数减少到10 CFU/g以下,达到贮藏和保鲜的目的。 但辐照食物可能引起的致畸、致癌、致突变性等安全问题存在的争议限制了该技术的使用。 利用超高压保鲜技术(液体或气体压力在100 MPa 以上称为超高压)可破坏微生物体内蛋白质的非共价键及其高级结构,使蛋白质凝固和酶失活。 还能造成细胞膜破裂,使微生物体内化学组分外流,对微生物具有致死作用。具有温度升高值很小,杀菌高效快速、均匀,能耗低的优点。 但超高压杀菌只能使用软包装材料,包装方式有限。 Préstamo 等[33]将豆腐样品在5 ℃、400 MPa 的超高压下分别放置5、30、45 min,发现高压处理可以减少豆腐中微生物的数量。 5 min 时菌落数值减少了0.31 lg CFU/mL,30 min 时数值减少了1.56 lg CFU/mL;在相同条件下,随着时间增长(45 min),细菌数量总体减少了2.38 lg CFU/mL;感官测试结果表明处理后的豆腐可以被消费者接受,加压样品的口感在良好范围内,高压处理过的样品比未经处理的样品口感更为细腻、适口感增加。 但高压处理加剧了豆腐中网状基质颗粒的分散,影响了豆腐的品质。 利用超高温瞬时杀菌技术将食品瞬间加热到135 ℃高温以上,食品中微生物的蛋白质肽链弯曲结构发生改变从而变性,3 ~5 s 可将微生物孢子完全杀灭,达到杀菌目的,而不影响食品的营养和色香味。 韦艳姿等[34]利用超高温瞬时杀菌技术将豆浆在(135 ±5)℃下保持3 ~5 s 后,发现豆浆中的所有细菌包括芽孢菌都被杀灭,产品口感保持良好。 结合无菌包装技术,豆浆可在常温下存放30 d 以上,极大延长了保质期。
低温等离子体技术是一种新型绿色的非热杀菌技术,近年来在食品加工业中的应用正在崭露头角。利用低温等离子体在放电过程中激发气体产生的紫外线破坏了微生物的结构,同时在放电过程中等离子蚀刻诱导产生含氧氮活性基团破坏了微生物的遗传物质及细胞膜与细胞壁结构,造成细胞质外泄,细胞死亡。 低温等离子体技术具有杀菌效率高,低温处理时间短、无毒、运行成本低的优点。 并且能够非热灭活潜在的腐败微生物[35]。 低温等离子体冷杀菌技术正在成为食品杀菌研究领域的热点,随着食品杀菌技术的快速提高,该技术具有广阔的应用和发展空间。 但目前多用于生鲜肉制品的保鲜杀菌中,尚未应用在豆制品中。
3.2.2 化学抑菌
目前被广泛应用于食品防腐中的措施是直接向豆制品中添加化学防腐剂,用来控制食品微生物的生长。 常用的化学防腐剂包括苯甲酸及其盐类、山梨酸及其盐类。 Koo 等[36]研究了大肠杆菌(Escherichia coli)、沙门氏菌(Salmonella)、李斯特菌(Liste-ria monocytogenes)和蜡样芽孢杆菌(Bacillus cereus)在豆腐上接种后,采用硫胺素二月桂基硫酸盐(thiamine dilauryl sulfate,TDS)对豆腐进行腐败菌抑菌实验。 结果表明,添加w=2%的TDS 显著降低了4 种菌的数量水平。 Shin 等[37]研究了异硫氰酸盐(isothiocyanates,ITCs)在10 ℃条件下对在贮藏的0、2、4、6、8、10 d 的豆腐腐败微生物影响。 结果表明:200 mg/kg 的ITCs 对储存10 d 的豆腐中的微生物生长有明显的抑制作用,颜色、味道、气味、咀嚼性和过度接受度等感官属性的偏好较高。 在贮藏初期,添加ITCs 对豆腐的感官性状(特别是口感和气味)有轻微的影响,而在10 ℃下贮藏10 d 后,豆腐的感官性状保持了良好。 但化学防腐剂滥用和过量使用会造成一系列食品安全问题,对人体健康存在着潜在威胁,使用范围受限。
3.2.3 生物防腐
从动植物、微生物中提取天然抑菌剂具有资源丰富、天然、高效的特点,在确保食品的微生物安全性、功能性的同时,还能赋予食品更高的营养价值,是一个有待开发的食品防腐新领域。 生物防腐剂包括动物源抑菌剂、植物源抑菌剂和微生物源抑菌剂。
3.2.3.1 动物源生物抑菌剂
动物源生物抑菌剂是一类氨基酸类、天然肽类、高分子糖类等物质。 直接从动物体中分离提取而来,对多种微生物具有抑菌活性。 郑卫等[38]发现w=1%的壳聚糖溶液可以抑制豆腐中大肠杆菌的生长,将豆腐保质期延长至3 d,保证豆腐的口感。王敏等[39]采用具有抑菌活性作用的蜂胶作为非发酵豆制品中的保鲜剂,结果显示w=0.4%的蜂胶可以明显抑制腐败菌的生长,使豆制品保质期延长。Kim 等[40]发现添加w=0.05%和w=0.1%牡蛎壳粉的豆腐制品,具有口感好,硬度适中,豆味较低的特点,可延长货架期。 然而,动物源抑菌剂的抗菌特性及其潜在的食品应用仍处于起步阶段,具有深远的潜力和应用空间。
3.2.3.2 植物源生物抑菌剂
植物源抑菌剂是指植物产生的具有抗菌作用的化合物,包括香辛料(精油)及中草药提取物(醛、酮、酯、醚、酸、萜类等)和茶多酚等,具有保护和延续物种及防御疾病的作用。 Cui 等[41]利用脂质体包裹的丁香油对豆腐中的金黄色葡萄球菌进行抑制研究,结果表明处理24 h 后,金黄色葡萄球菌的数量减少了99.87%。 卫强等[42]发现栀子叶挥发油可使豆腐保鲜期延长6 d,对豆腐中的菌群有较强的抑制作用。
在食品腐败生态系统中,细菌行为受到细菌群体感应(quorum sensing,QS)系统的控制。 来自食品加工环境中的病原体和腐败细菌定植在食品表面定植形成了生物膜。 大部分革兰氏阴性菌的生物膜形成是由QS 调控的。 香兰素具有明显的抗QS 表型抑制作用。 Li 等[43]研究了植物源抑菌剂——香兰素对食品中腐败菌蜂房哈夫尼菌的抑菌机理。 我们将Li 的研究归纳出作为植物源抑菌剂香兰素的抑菌机理,见图2。 由图2 可知,香兰素可与靶蛋白相互作用,与靶蛋白竞争结合;可以使相关基因表达水平下调,干扰QS 系统主要信号分泌分子——酰化高丝氨酸内酯(n-acyl-homoserine lactone,AHL)的合成从而阻止生物膜的形成。 这些发现将有助于拓宽香兰素作为QS 抑制剂的应用。 目前,植物源抑菌剂仍处于研究探索阶段,尚未达到商业化应用。
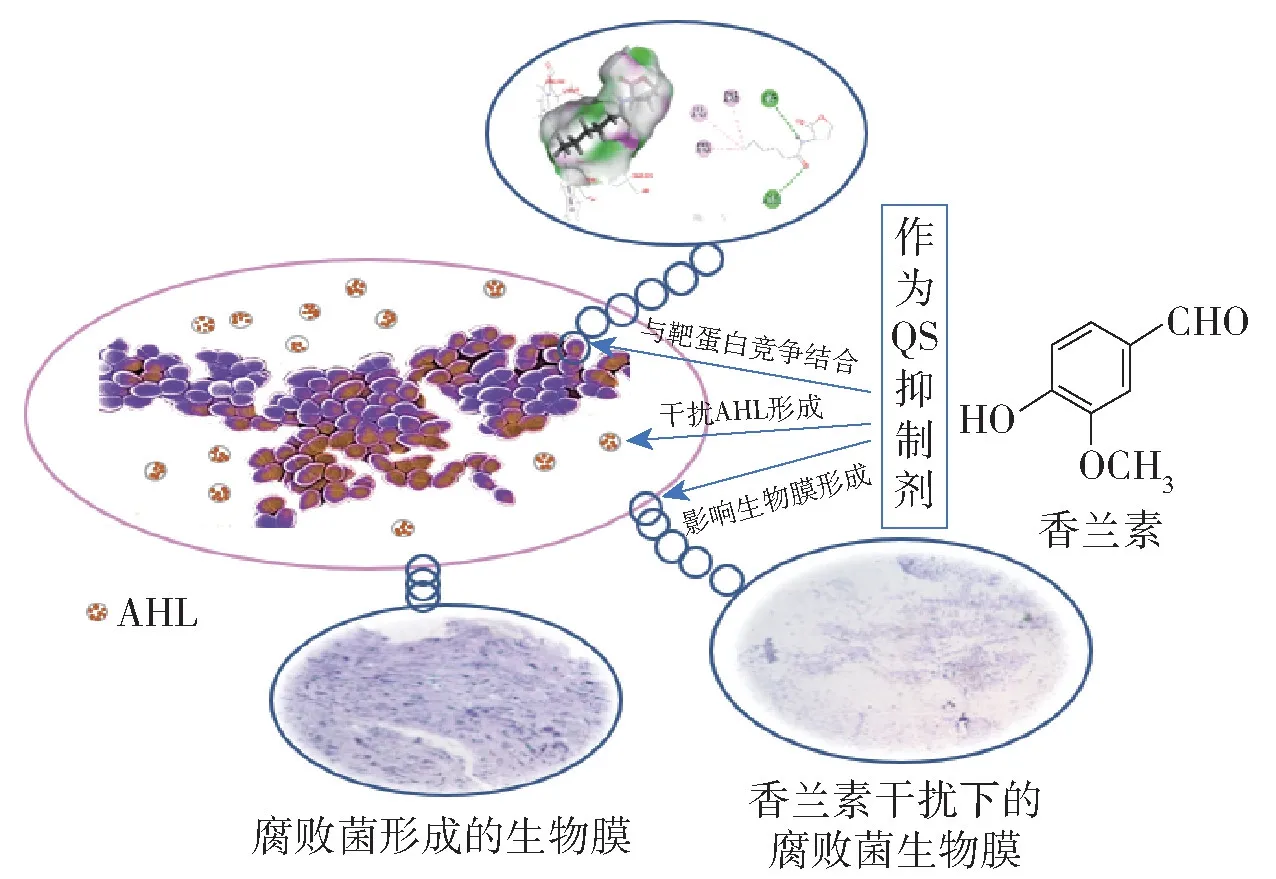
图2 植物源抑菌剂香兰素的抑菌机理Fig.2 Bacteriostatic mechanism of vanillin as a plant-derived bacteriostatic agent
3.2.3.3 微生物源生物抑菌剂
微生物源生物抑菌剂是指某些微生物本身或由其产生的代谢产物,具有抑制微生物生长繁殖的作用。 常用的微生物源抑菌剂有4 种(见图3):乳酸链球菌素(Nisin)、纳他霉素、ε-聚赖氨酸和普鲁兰多糖,微生物源生物抑菌剂的微生物源、作用特点及抑菌有效范围不同。 由乳酸菌产生的抗菌肽、乳酸链球菌素,可抑制革兰氏阳性菌的生长;由那他链霉菌产生的纳他霉素,对几乎所有霉菌和酵母有效;由放线菌产生的抗菌活性功能多肽、ε-聚赖氨酸,可破坏微生物的细胞膜;由出芽短梗霉产生的胞外中性多糖、普鲁兰多糖,与其他抑菌剂使用可协同增效。

图3 4 种常用的微生物源抑菌剂Fig.3 Four commonly used microbial-derived bacteriostatic agents
将这些微生物源抑菌剂的作用机制进行归纳,见图4。 由图4 可知,抑菌物质靠近细菌细胞膜后在静电力的作用下,插入到磷脂双分子层中,慢慢在细胞膜上形成微孔,导致膜通透性增加,离子通道遭到破坏和能量产生系统受到干扰,造成细胞内的Ca2+、K+等外流,细胞内容物(氨基酸、维生素等小分子物质)渗出,新陈代谢发生紊乱,破坏了腐败菌细胞膜屏障,导致菌体死亡。 Ye 等[44]认为ε-聚赖氨酸可能的抗菌机制是当ε-聚赖氨酸与大肠杆菌O157:H7 接触时,通过静电吸引与细菌膜表面结合。 这种相互作用的积累造成膜扰动,导致细胞膜结构的变化而断裂。 因此,ε-聚赖氨酸通过细胞膜断裂进入细胞质。 ε-聚赖氨酸诱导活性氧(reactive oxyen species,ROS)和DNA 损伤反应(SOS 反应)的产生,影响细胞死亡,调控相关基因的表达水平。 这些因素的协同作用或累积影响细胞,最终导致细菌的死亡。 Punyauppa 等[45]认为乳酸链球菌素的抑菌机理为:使用N 端结合基序与脂质Ⅱ的碳水化合物焦磷酸部分结合。 这使得乳链球菌素的C 端片段能够插入细胞膜。 多个Nisin-脂质Ⅱ复合物在靶细胞细胞膜上组装形成一个稳定的直径为2 nm 的孔隙。 它可能导致膜透性的增加,导致膜电位的耗散,流出的小胞质内容物如核苷酸和离子等。 因此,受损的细胞既不能产生能量也不能产生重要的大分子,致使细胞死亡。

图4 微生物源抑菌剂抑菌机理Fig.4 Bacteriostatic mechanism of microbial bacteriostatic agents
葛子榜[46]将微生物源保鲜剂添加到豆腐,研究其对豆腐中腐败菌的影响。 发现将普鲁兰多糖、ε-聚赖氨酸和双乙酸钠3 种保鲜剂进行复配,对腐败菌的抑制有明显的协同增效作用。 对豆腐的色香味以及品质无太大影响,可替代化学防腐剂,豆腐保存天数得到延长。 Schillinger 等[47]应用低剂量的乳酸链球菌素来防止豆腐中李斯特菌的繁殖。 Chen等[48]将产细菌素的魏斯氏菌(Weissella hellenica)D1501 作为发酵剂和凝固剂加入到新型乳酸菌豆腐中,发现其能够抑制豆腐中的腐败菌和致病菌的生长,提高了豆腐的安全性和货架期。
同时,如何保持微生物源抑菌剂的抑菌活性也成为当前的另一个研究热点。 张恒等[49]发现在豆浆生产过程中加入浓度为1.0 g/kg 的Nisin,能良好保持豆腐原有的品质与风味,保鲜效果明显。 葛子榜在豆腐制作过程中点脑的环节时将微生物源抑菌剂复配物加入,发现普鲁兰多糖与ε-聚赖氨酸组合的抑菌效果最好[46]。 其他方法-包埋微生物源抑菌剂制备微胶囊。 由于乳酸链球菌素、溶菌素、纳他霉素等微生物源抑菌物质在食品储存或加工过程中存在容易降解的问题,降低了抗菌活性,限制了使用范围,微胶囊技术可增加稳定性、防止失活[50]。 Wu等[51]发现壳聚糖——Nisin 微胶囊比单独使用Nisin对微生物生长的抑制效果高,保存效果较好。 因此,依据豆制品致腐微生物的种类,通过对不同抑菌特性的微生物源保鲜物质进行合理组合,再进行微胶囊化,增强抑菌效果,达到稳定、高效、广谱抑制鲜食豆制品腐败菌的目的。
4 展望
由于制作传统豆制品的原料大豆、加工过程、储存环境等环节均是产生豆制品腐败变质的来源。 故在原材料和产品的加工和储存方面,要加强卫生管理,制定精确、可实现的工艺参数,规范处理流程。高标准和严格的车间卫生要求才可确保产品质量。传统豆制品在贮藏过程中,在不同贮藏期的腐败菌不同,随着贮藏时间的延长,在保存环境中适宜生长的部分细菌会抑制其他几种细菌的生长而成为优势腐败菌,腐败菌群体系呈现出不同优势菌群的动态菌相变化。 在应用传统方法结合现代技术对引起传统豆制品腐败变质的腐败菌进行系统的分离和鉴定的基础上,针对特定腐败菌进行抑制成为延长传统豆制品保质期的关键之一。 同时,在传统豆制品生产加工过程中,针对不同的传统豆制品品质及腐败菌的特点,采用超高温瞬时杀菌技术和等离子体杀菌技术等物理方法近年来日益成为热点。 同时,微生物源抑菌剂因其安全高效、抑菌效果好等特点日益受到广泛关注,并逐渐应用于食品保鲜。 但目前该类抑菌剂在传统豆制品保鲜中的应用多停留在实验室研究阶段,且一种或一类微生物源抑菌剂存在抗菌谱窄、作用范围小等问题,限制了研究的深入和应用范围的拓宽。 将物理方法结合生物方法抑菌剂对传统豆制品进行保鲜将成为新的方向。 为有效延长传统豆制品保质期提供独特的优势,未来的应用前景可期。
