基于益生菌及其代谢物的食源性致病微生物控制技术研究进展
2022-07-02励建荣檀茜倩崔方超吕欣然
励建荣, 檀茜倩, 王 丹, 崔方超, 吕欣然
(渤海大学 食品科学与工程学院/生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心, 辽宁 锦州 121013)
益生菌是在积累到一定数量的时候对人体健康产生有利影响的一类活的微生物,具有提高免疫力、增强肠道菌群调节能力、治疗代谢疾病和改善情绪等多种功能[1-4]。 益生菌种类很多,其中的乳酸菌,包括乳杆菌属(Lactobacillus)、明串珠菌属(Leuconostoc)、片球菌属(Pediococcus)、链球菌属(Streptococcus)、肠球菌属(Enterococcus)、乳球菌属(Lactococcus)、魏斯氏菌(Weissella)等[5]中的大部分,是公认的一类安全益生菌,其为革兰氏阳性、微需氧、不形成芽孢、接触酶阴性;另一类研究较多的益生菌为芽孢杆菌属中的一部分细菌,其菌株特点为革兰氏阳性、需氧或厌氧、可形成孢子,孢子的形成使其能更好地耐受高温、高渗透压和酸性环境,相比于乳酸菌在应用上更具优势[6];此外还有布劳特氏菌属(Blautia)的部分细菌,属毛螺菌科,多分布于哺乳动物的粪便和肠道,这个菌属的一些菌曾经一度被划分到梭菌属和瘤胃球菌属中,其生长特性为严格厌氧,不具有运动性,成对或者成链状出现,大部分菌株不形成孢子但对环境(酸、盐和氧化反应)具有较强的耐受性[7];一些与肠道共生的真菌比如鲍氏酵母菌也被认为具有益生功效[8]。 食源性致病微生物在食品中的存在不仅会加速食品腐败也会引发食用安全风险。 目前研究发现,很多益生菌及从其发酵液中分离的代谢物都具有抑制食源性致病微生物的作用,具有在食品安全和品质控制方面应用的潜力;并且由于其兼具益生特性,相对于其他食源性致病微生物的控制技术更具优势,有很好的开发前景[9-11]。 本研究拟分析现有益生菌及其代谢物对食源性致病微生物的抑制机制,基于目前益生菌及其代谢物在食品安全和品质控制中的应用研究实例,阐述其在果蔬、肉类等食品中的应用方式、应用条件、对货架期的延长效果,并对其应用研究趋势进行探讨。希望为基于益生菌及其代谢物的食品安全和品质控制技术开发和应用提供有益的借鉴和参考。
1 抑制食源性致病微生物的益生菌及其代谢物类型
益生菌及其产生的代谢物种类繁多,研究发现,很多益生菌及其代谢物具有抑制包括细菌、真菌、病毒在内的多种食源性致病微生物的功能。 益生菌对食源性致病微生物的抑制效果不仅取决于其种类、培养条件、分泌的代谢物种类,也与食源性致病微生物本身的特性(种类、耐药性、生物被膜形成能力、毒力因子产生能力)有关[12-13]。 表1 列出了部分对食源性致病微生物有抑制作用的益生菌及其代谢物类型。 益生菌的生长和代谢受到自身特性和多种环境因素的影响,不同来源的同种益生菌的抑菌谱也有差别。 益生菌的代谢产物成分复杂,对食源性致病微生物起到抑制作用的活性物质主要有肽类,如抗细菌肽和抗真菌肽,包括乳酸链球菌素(Nisin)、乳酸片球菌素(pediocin)、枯草菌素(subtilin)等;有机酸类,如乳酸、丙酸、醋酸、苯乳酸、吲哚乙酸等;此外还有过氧化氢、醇类、酚类、酶类和多糖类等多种物质。 可通过微生物的抑制实验结合分子生物学(基因检测或基因组分析)以及仪器分析[气相色谱-质谱或高效液相色谱-质谱(GC-MS 或HPLCMS)]等手段对这些活性物质进行探定和分离。
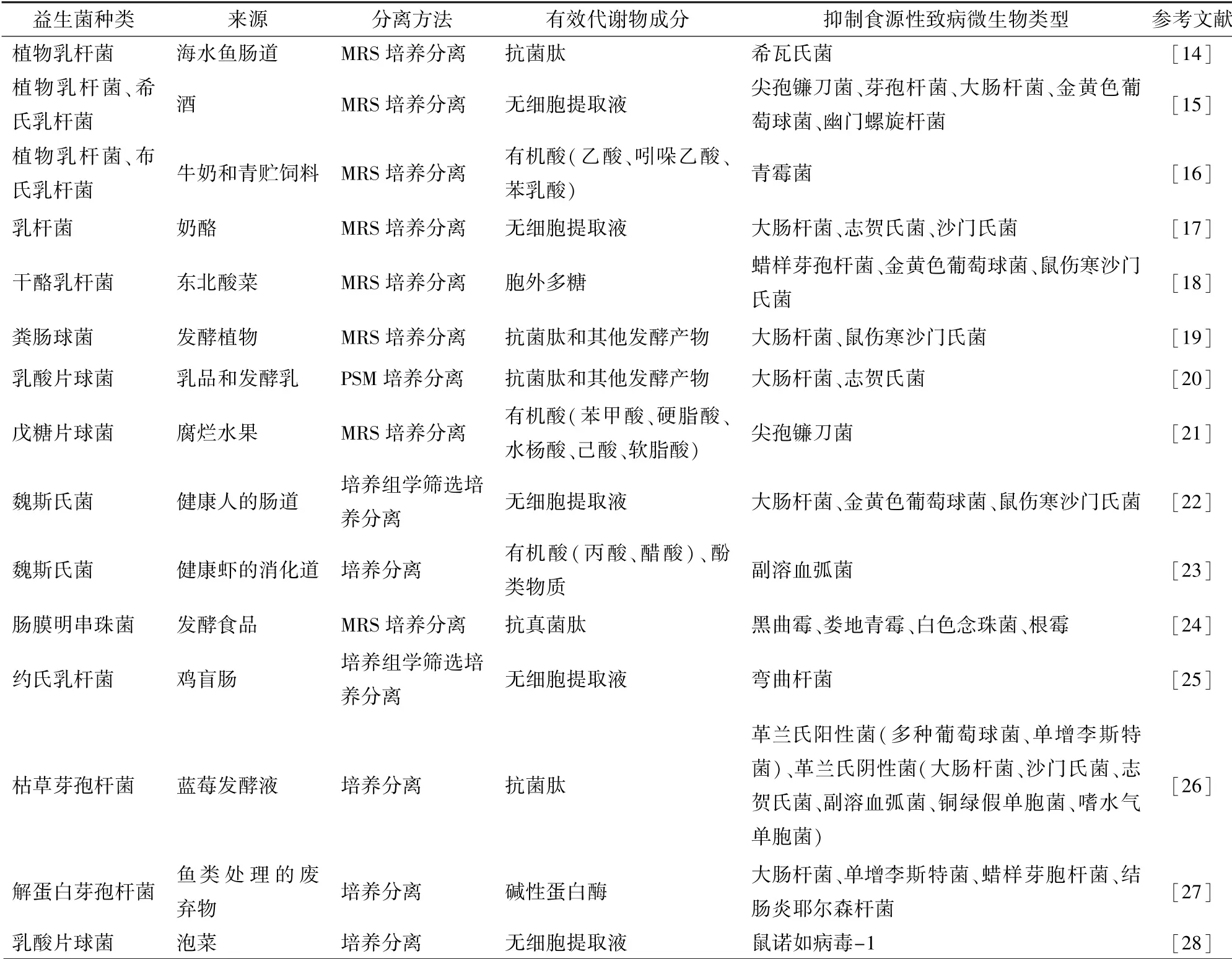
表1 抑制食源性致病微生物的益生菌及其代谢物Tab.1 Inactivation effect of probiotics and their metabolites against foodborne pathogens
目前大量具有抑制食源性致病微生物的益生菌资源尚待挖掘,因为大部分来源的样品菌群成分复杂[29]。 随着高通量测序技术的发展,常利用扩增子技术或宏基因组学技术对样品中的菌群结构进行分析,在确定其中具有益生潜力的菌株类型后可针对性采用合适的培养基对益生菌进行分离[30-31];同时,培养组学的发展也加速了益生菌株资源的挖掘和研究。 培养组学是将优化的培养条件应用于微生物群研究的一种技术,将基质辅助激光解吸/电离-时间飞行质谱(matrix-assisted laser desorption/ionization-time of flight mass spectrometry,MALDI -TOF MS)与16S rRNA 扩增子测序技术结合对菌株进行鉴定。 培养组学高通量的培养条件可获得大量菌株,尤其适宜一些对生长条件要求挑剔菌株的培养,常应用于新益生菌资源的挖掘中[32]。
2 益生菌及其代谢物对食源性致病微生物的抑制机制
2.1 对细菌的抑制机制
食源性致病菌常以两种状态存在,浮游态和生物被膜态。 细菌的生物被膜是细菌为了应对极端环境所采取的保护行为。 生物被膜的形成始于黏附,是一个涉及菌群结构、功能和组分之间变化的动态过程。 细菌会被大量的胞外多糖骨架包裹,附着在食品或者加工设备的表面,使其清除变得更加困难[33]。 益生菌及其代谢物对细菌的抑制机制主要包括破坏细胞结构,干预细菌内部DNA、RNA、蛋白、细胞壁和脂肪酸等生命活动所需要的关键物质合成,与细菌竞争关键性的营养物质,阻断关键的能量代谢通路,干扰细菌群体行为和破坏生物被膜结构等等。 这些机制与益生菌及其代谢物与食源性致病菌的类型和特性有关,常同时包含多种机制。 图1 列举了其中的部分机制。
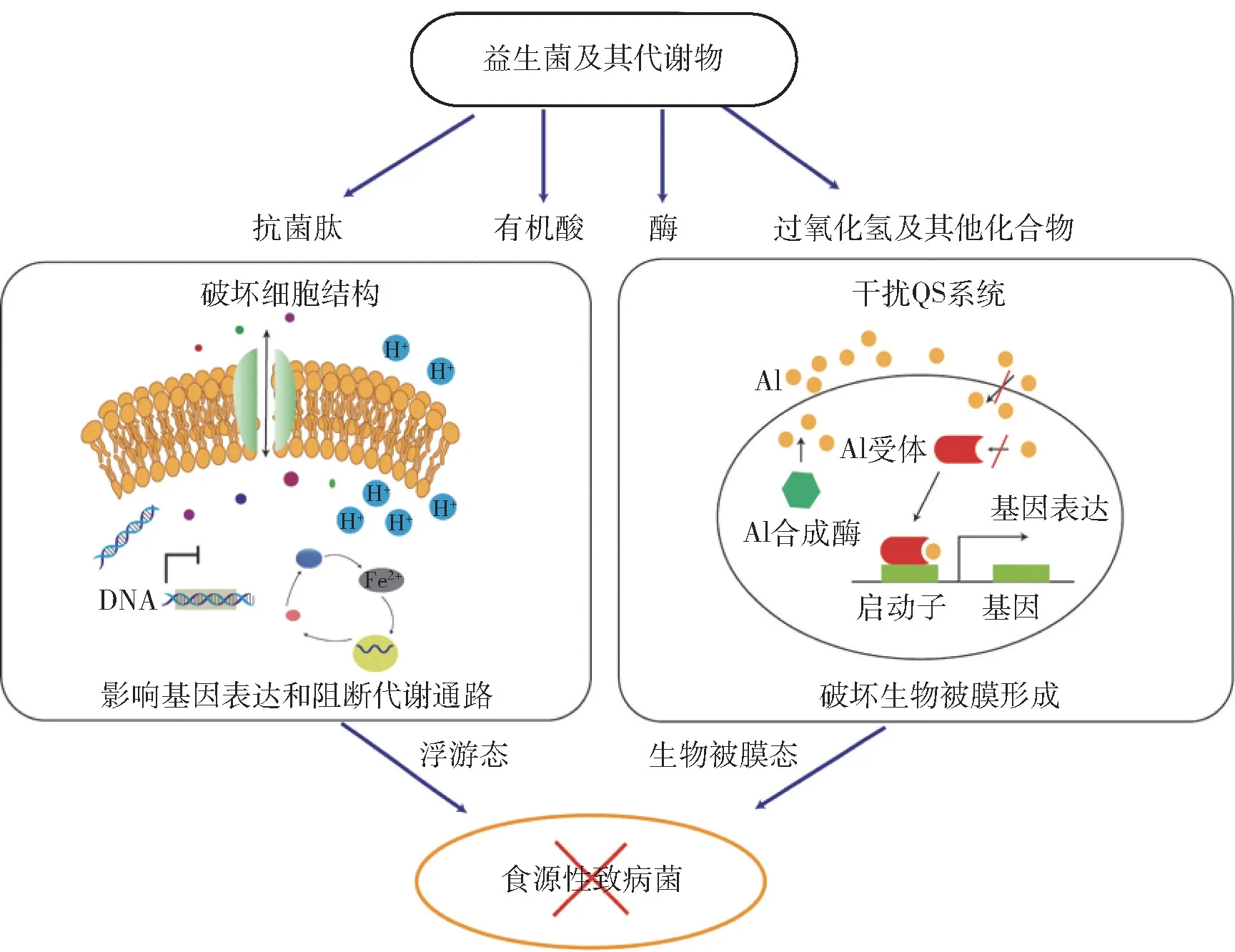
图1 益生菌及其代谢物对食源性致病菌的抑制机制Fig.1 Inactivation mechanism of probiotics and their metabolites against foodborne pathogens
2.1.1 对浮游状态细菌的抑制机制
2.1.1.1 破坏细胞壁完整性
益生菌分泌的代谢物会作用于细胞壁,在细胞壁上形成孔洞,引起内部物质溶出,最终导致细胞死亡。 很多抗菌肽对细菌的抑制多采用这种机制,且其抑菌谱与抗菌肽结构和食源性致病菌的类型有关。 与细胞壁上的受体结合是抑制过程的第一步,结合位点包括脂质Ⅱ和磷酸转移酶系统的甘露糖渗透酶(mannose permease of the phosphotransferase system,man-PTS)等。 脂质Ⅱ作为膜锚定的细胞壁合成前体,对细胞壁肽聚糖的生物合成至关重要。 Nisin 和很多羊毛硫类细菌素都以脂质Ⅱ作为其对接位点[34]。 脂质Ⅱ不是pediocin 与细胞结合的靶标位点,pediocin 分子与目标细菌接触后,其分子上的正电荷会与细胞壁上的磷脂壁酸的负电荷结合,随后pediocin 分子插入这些目标细菌的细胞质膜,形成孔洞[35]。 另一种由戊糖片球菌分泌的新型抗菌肽pentocin ZFM94 在细胞上形成孔洞同样不以脂质Ⅱ为受体,其具体结合位点还未明确,但这种抗菌肽抑菌谱很宽,对革兰氏阳性菌、革兰氏阴性菌和真菌都具有抑制作用,探明其结合机制对其应用有十分重要的作用[36]。 分离自植物乳杆菌SHY 21-2 的植物乳杆菌素LP 21-2 (分子质量为1362.82 Da,少于20 个氨基酸)也同样具有较宽抑菌谱,对包括金黄色葡萄球菌、单增李斯特氏菌、大肠杆菌、沙门氏菌、青霉等在内的细菌、真菌均具有抑制作用,扫描电镜和透射电镜分析发现其可在细胞壁形成孔洞和褶皱以及破坏细胞膜和原生质层的完整性,但具体结合位点和机制有待进一步探究[37]。
2.1.1.2 破坏细胞膜的通透性和完整性
细菌的细胞膜是一种选择透过性膜,通过与外界环境有选择性地交换物质为内部提供利于各种关键生物化学反应进行的条件。 细胞膜有选择性地允许保持细胞膜正常功能的K+、Na+、H+等离子通过。 研究发现,动物双歧杆菌BB04 分泌的抗菌肽bifidocin A 会使大肠杆菌细胞膜的通透性增加,造成细胞内K+和无机磷大量渗出导致细胞死亡[38]。细胞膜正常的选择通透性也对细胞维持正常酶活力和代谢平衡非常重要,细胞膜通透性的改变会造成铁离子失衡,进而影响到细菌能量代谢(转运、代谢调控、调节细胞压力和运动)有关的各种功能,导致细菌死亡[39]。
细菌的细胞膜还是细菌应对外界环境不利因素的屏障,细胞膜完整性的破坏会造成细菌死亡。 研究发现,鼠李糖乳杆菌代谢分泌的新型细菌素1.0320(表面带有负电荷、分子质量在1 ~3.3 kDa)具有破坏大肠杆菌UB1005 细胞膜的完整性和通透性的功能[40]。 芽孢杆菌JS-4 产生的抗菌肽(分子质量为3346.6 Da,分子序列与subtilin 类似,但15位的丙氨酸和24 位的亮氨酸分别被缬氨酸和异亮氨酸替代)也通过破坏细胞膜的通透性和完整性,造成细胞内外渗透压的失衡,进而间接影响代谢、复制、转录和翻译等一系列的生物学过程来抑制食源性致病菌。 研究证实此抗菌肽有很好的热和pH 稳定性,但是容易被蛋白酶降解,其结构特点可作为进一步探明其机制的基础[41]。
2.1.1.3 影响基因表达和阻断代谢通路
基因决定蛋白表达,对基因的破坏会阻断细菌某些重要酶和受体的合成,最终导致细菌死亡。 益生菌的一些代谢物会破坏细菌的遗传物质。 研究发现,苯乳酸可插入单增李斯特菌和大肠杆菌的基因组中,随后影响DNA 的复制、转录和表达,最后阻碍细菌生长导致细菌死亡,与溴化乙锭作用机制类似[42]。 益生菌的另一些代谢产物还可以通过影响基因表达和细菌正常代谢通路来抑制细菌。 研究发现,细菌素BM1157 可使大肠杆菌蛋白表达水平发生明显改变,其原生质组分和细胞膜组分显著降低,周质空间也相对减少,蛋白酶和蛋白质-DNA 复合物的表达水平上调;同时大肠杆菌与铁的结合能力降低,铁缺失进而使其催化酶和水解酶活性降低,碳水化合物代谢和转运受到显著影响,导致生物活性分子合成时所需的物质和能量供应不足,氨基酸转运和脂质代谢等过程进而受到阻断,细胞分化受到影响,最终导致细胞凋亡[43]。
2.1.2 对生物被膜态细菌的抑制机制
益生菌及其代谢物可通过干扰细菌的群体感应信号通路来实现对生物被膜的控制,与杀死细菌的抑菌机制不同,这种机制以阻碍细菌的生长为目的,不容易使细菌产生耐药性。 群体感应(quorum sensing, QS)是细菌之间依靠其所分泌的信号分子(autoinducer, AI)的浓度进行交流的一种机制,与AI 的浓度呈现正向的相关性,能影响基因表达的调控,对细胞的运动、抗生素的合成、生物被膜的形成、毒力因子的产生都具有非常重要的作用[44-45]。 不同细菌QS 系统产生的信号分子种类不同,主要包括高丝氨酸内酯类(HSL)、寡肽(AIP)和AI -2 三大类[46-47]。 群体感应淬灭(quorum quenching, QQ)即通过干扰细菌QS 系统使其QS 现象消失,具有QQ 功能的物质可以称为QS 抑制剂,其抑制机制主要包括分解信号分子以及干扰信号分子与受体结合等。 在益生菌的代谢物中发现多种天然的QS 抑制剂[48-49]。 研究发现,乳酸片球菌产生的乳酸可以抑制铜绿假单胞菌的las 和rhl 两种QS 系统,影响了C4-HSL 等短链的HSL 的产生,进而抑制铜绿假单胞菌的群集和泳动行为,减弱其生物被膜以及其他毒力因子(弹性蛋白酶、绿脓菌素等)的形成能力[50]。 干酪乳杆菌CRL431 和乳酸片球菌CRL730发酵液的三氯甲烷提取物可抑制铜绿假单胞菌,显著影响其生物被膜形成、生物量以及代谢活动,同时可抑制其毒力因子的产生[51]。 部分芽孢杆菌和乳酸菌可分泌群体感应淬灭酶(QQ 酶),通过水解信号分子的内酯环,破坏信号分子酰基侧链的酰胺键,还原信号分子阻止其被受体识别等机制,影响相应食源性致病菌生物被膜的形成[52-54]。
另外益生菌及其代谢物还可以通过阻止致病菌黏附行为的发生,与致病菌竞争和外排作用来达到清除和抑制食源性致病菌生物被膜的目的。 研究发现,乳酸片球菌PA003 可抑制大肠杆菌、沙门氏菌、金黄色葡萄球菌和单增李斯特菌在不锈钢、聚氯乙烯和玻璃表面形成生物被膜,对已经形成的生物被膜通过竞争和外排作用破坏生物被膜结构达到清除生物被膜的目的[55]。 多株乳酸乳球菌也可通过抑制单增李斯特菌的黏附过程来抑制其生物被膜形成[56]。
2.2 对真菌的抑制机制
益生菌对真菌的抑制作用主要是通过其产生的抗真菌肽以及有机酸、羧酸等物质进行的,抑制机制与其对真菌细胞结构的破坏和与真菌互相竞争营养物质有关。 研究发现,芽孢杆菌(Bacillus siamensisLZ88)代谢产物中的两种肽类物质iturins 和macrolactin,以及挥发性的有机酸二甲基丁酸可使互隔交链孢霉菌丝发生明显的形态改变,使菌丝形成孔洞,并发生不规则萎缩和减少其上附着的分生孢子数目,这些不带分生孢子的菌丝随之会发生断裂、萎缩、瓦解和死亡[57]。 益生菌也会和真菌竞争环境中的营养因子,尤其当环境中的营养素含量较低时,益生菌会优先利用营养素,其结果就是环境中剩下的营养物质不足以保证真菌孢子萌发,导致孢子的活性受到抑制。 虽然当环境中的营养素浓度增高达到孢子萌发所需要营养条件时其可以正常萌发,但研究发现其萌发率要远低于未经处理时的状态,说明益生菌代谢物对真菌孢子结构也造成一定程度的损伤[58]。 益生菌及其代谢物可清除和抑制真菌生物被膜,研究发现其可抑制多株白色念珠菌生物被膜的形成,其机制主要与阻止生物被膜的黏附以及同真菌竞争营养物质有关[59]。
2.3 对病毒的抑制机制
益生菌及其代谢物对病毒的抑制机制包括干扰病毒复制和阻止其与宿主细胞结合等,具体机制还有待于进一步研究。 已发现乳酸菌的一些无细胞提取物尤其是乳酸可抑制病毒的复制[60]。 还有研究发现,植物乳杆菌LBP-K10 分泌的一种环二肽可以抑制流感病毒(H3N2),其抑制机制与其丰富的手性结构有关,其中4 个可以改变立体化学和构象结构的位点被认为在抗病毒中起到比较关键的作用,并可能通过与人蛋白酶或者病毒血球凝集素裂解蛋白互作阻止病毒感染宿主细胞[61]。
3 基于益生菌及其代谢物的食源性致病微生物控制技术的应用
基于益生菌及其代谢物的食源性致病微生物控制技术是利用益生菌及其代谢物所具有的抑制食源性微生物功能特性,将其应用于食品来达到对食品安全和品质进行控制的技术。 这种技术会以从益生菌发酵液中分离提取的某些纯化或半纯化产物(单一代谢物)、益生菌无细胞提取液(多种代谢物的复合物)、益生菌(活菌)的形式应用到对食品的品质控制中。 益生菌及其代谢物的类型、应用方式、应用条件、食品基质的类型、食源性致病微生物的种类都会对其技术应用效果(食品安全性、货架期的延长和食品品质保持)产生不同的影响。
3.1 在果蔬安全与品质控制中的应用
果蔬的水分和糖含量较高,容易受到机械损伤,在食源性致病微生物污染以及酶的作用下发生品质劣变,同时因其食用方式多为生食,故存在引发食源性疾病的风险。 基于益生菌及其代谢产物的食源性致病微生物控制技术目前已经应用于果蔬从采后保藏到加工等多个环节,可以保证果实的鲜度和安全性。
益生菌及其代谢物可应用于果蔬的保鲜保藏,研究发现,pediocin 对莴苣、生菜、芹菜和菠菜等新鲜蔬菜具有很好的品质控制效果,可有效降低新鲜蔬菜在贮藏期间产生单增李斯特菌的数目[62]。 番茄和酸面团中分离的植物乳杆菌TR7 和植物乳杆菌TR71 发酵液可延长番茄的鲜度和贮藏时间,这两种益生菌无细胞提取液(活性成分为乳酸、乙酸、苯乳酸和吡嗪衍生物)可以很好地抑制扩展青霉和黄曲霉的生长, 霉菌最小抑菌浓度在6.3 ~100.0 g/L,在贮藏期间能很好地控制番茄品质,避免其腐败[63]。
益生菌及其代谢物可对鲜切水果中的食源性致病微生物进行控制,保证消费者食用的安全性和口感。 利用鼠李糖乳杆菌GG 直接喷涂到鲜切苹果的表面,可在贮藏期间不同程度降低单增李斯特氏菌和沙门氏菌的菌落数,虽未达到完全杀灭效果,但根据体外实验,两种食源性致病菌在模拟消化道内的生存能力下降,说明其对宿主致病力有所减弱;同时贮藏期间鲜切苹果的颜色、可溶性固形物和硬度可得到很好保持[64]。 将鼠李糖乳杆菌GG 喷洒于鲜切梨表面,可在贮藏期间显著控制单增李斯特氏菌的菌落数;鲜切梨果肉可溶性固形物和总酸含量在整个贮藏期基本保持稳定;风味物质(酯类、醇类、醛类、萜类、酮类、酸类)种类多样,醇类和酯类的特征物浓度随着贮藏时间延长而升高,比不处理以及采用氯化钙处理更有效地保证了梨的特征风味[65];鲜切梨的硬度、外观以及抗氧化性也比不采用任何处理和氯化钙处理的效果好[66]。
益生菌及其代谢物也可以应用于非发酵型果汁饮料品质的控制。 鼠李糖乳杆菌GG 能控制低温和常温贮藏期间沙棘果汁饮料中大肠杆菌的生长以及保证果汁品质,其控制效果与益生菌的适宜生长温度有关。 相对高的温度下鼠李糖乳杆菌的代谢旺盛,可快速分泌有机酸和抗菌肽,因此会使大肠杆菌的数量在储藏前期即发生非常明显下降;而低温贮藏条件下大肠杆菌数目则在贮藏中期开始下降[67]。利用益生菌对非发酵果汁进行品质控制的过程中应合理评估益生菌与致病菌的生长互作和果汁风味保持之间的关系。
3.2 在肉类安全与品质控制中的应用
肉及肉类制品的味道鲜美、品种和营养丰富,在人们日常膳食中占比较大,是获取蛋白质和脂肪等营养素的主要来源。 肉类容易被大肠杆菌、单增李斯特氏菌、沙门氏菌、副溶血弧菌等致病微生物污染,在食用过程中杀菌不彻底以及致病菌产生的毒素会引起很大的食用安全隐患。 受到日式和地中海饮食方式的影响,消费者对一些红肉和水产品(三文鱼、金枪鱼、牡蛎)会倾向采用生食方式,根据益生菌及其代谢物的特点,人们对其在肉类安全与品质控制方面也开展了大量研究。
益生菌可以作为发酵剂保证发酵肉类品质。 采用植物乳杆菌和德氏乳杆菌作为发酵剂生产萨拉米香肠,可以使香肠中的梭状芽胞杆菌的菌落数下降2 lg CFU/g 左右。 虽然萨拉米香肠在制作过程中的环境(较高的脂肪含量和熟化过程不断增高的盐浓度)对益生菌生长不利,益生菌抑菌效果略低于化学防腐剂,但是益生菌的加入可减少香肠生产过程中的亚硝酸盐添加量,提高发酵肉类品质,满足当前消费者的绿色、健康、微加工、无添加食品选购理念[68]。 益生菌的添加既作为发酵剂,也兼具生物保护剂的作用,已经被应用于包括香肠、熏肉等多种肉品安全和品质控制中。
除了作为发酵剂,也可以将鲜肉浸泡于益生菌液处理一定时间来达到保证鲜度和安全性的效果,在应用中需考虑益生菌的初始活菌数目以及贮藏条件等因素。 研究发现:初始活菌数目为7.5 lg CFU/g 的干酪乳杆菌和弯曲乳杆菌混合液的浸泡处理可以显著降低低温真空贮藏牛肉样品中的致病微生物以及TVBN 值;处理后牛肉表面菌落复杂程度明显低于未处理组,其中腐败微生物肠杆菌、假单胞菌和热杀索丝菌数目显著降低。 在品质方面,未经处理的牛肉样品于贮藏中后期TVBN 开始上升迅速,在贮藏末期达到15 mg/100 g,已超中国冷鲜肉卫生标准GB2707—2016;而益生菌处理可使货架期延长10 d,且贮藏末期TVBN 值只有10 mg/100 g 左右,满足国家标准[69]。 还有研究利用干酪乳杆菌CTC494 控制低温贮藏的真空包装烟熏三文鱼中的单增李斯特氏菌,来保证烟熏三文鱼微生物安全性和品质。 研究发现,烟熏三文鱼的品质控制效果与益生菌菌株有很大关系,相对于干酪乳杆菌CTC494,弯曲乳杆菌CTC1742 对烟熏三文鱼中单增李斯特菌的控制能力相对较弱[70]。
3.3 技术应用研究趋势
3.3.1 开发高产高效抗菌肽
抗菌肽是目前在食品品质控制中应用较多的一种益生菌代谢物,其中Nisin 已被多国批准作为食品添加剂使用,因为受到纯化技术的影响,Nisin 的一种半纯品Nisaplin 的工业应用比较常见[71]。 虽然抗菌肽的分离困难,使用成本较高,部分抗菌肽的抑菌谱较窄,但是近年来还是受到了持续关注和广泛研究;人们还结合数据科学等技术针对已有抗菌肽抗菌特性开发了相应的数据库,利用基因组学数据的自动识别和筛选评估其安全性[72],已筛选到很多具有相对宽抑菌谱的抗菌肽。 提高抗菌肽产量的手段包括对益生菌的培养条件进行优化、对菌株进行基因工程改造以及人工合成等,同时需要配合加速下游益生菌纯化技术的开发[73-74]。 高产高效抗菌肽的开发可为益生菌代谢物在食品安全和品质控制中的工业化应用提供一定基础。
3.3.2 提升益生菌抑制微生物性能
对益生菌培养条件的优化可提升其抑制微生物的性能,以达到更好地控制食源性致病微生物的效果。 益生元的添加可以提高益生菌有效代谢物产量并在一定程度上增强其抑制食源性致病微生物的能力。 研究发现,岩藻多糖的添加可提高鼠李糖乳杆菌GG 代谢产物的产量和抑菌能力。 岩藻多糖可促进鼠李糖乳杆菌GG 有机酸的产生,傅里叶变换红外光谱(FT-IR)分析发现,鼠李糖乳杆菌GG 细胞的磷脂双分子层受到岩藻多糖的影响,膜蛋白二级结构发生改变,细胞形态变长;低质量浓度(8 mg/mL)相对于高质量浓度(80 mg/mL)岩藻多糖的添加对鼠李糖乳杆菌GG 抑菌性能的提升作用更强,这可能与高浓度益生元添加造成的高渗透压和快速生长分泌的有机酸影响了鼠李糖乳杆菌GG 的某些信号通路有关[75],其具体机制有待于进一步研究。 另外将益生菌在比较温和的压力下培养也可以增强其抑菌性能[76],但其机制以及在食品品质控制中的应用效果也有待于进一步的研究证实。
3.3.3 人工合成多功能益生菌
可考虑采用基因工程手段构建具有多重功能的益生菌。 有研究构建了一种可同时分泌γ-氨基丁酸(GABA)(抗氧化剂)以及抗菌肽Nisin 的功能益生菌F44/GadB1C1,使其发酵液同时具有控制食源性致病微生物和抗氧化作用。 这株菌的发酵过程采用pH 调节的两段式发酵,控制GABA 以及Nisin 的生物合成,发酵过程结束后F44/GadB1C1 菌的GABA产量可以达到9.12 g/L,是恒定pH 值发酵过程GABA产量的2.2 倍。 F44/GadB1C1 冻干的发酵产物(具有抑菌剂和抗氧化剂的双重功能)可显著降低猪肉冷藏期间表面微生物水平,对大肠杆菌和乳酸菌的抑制效果高于山梨酸钾,并可很好地控制猪肉脂质氧化,猪肉样品的TBARS 值在贮藏期间上升缓慢,与添加叔丁基羟基茴香醚对脂肪氧化的抑制效果类似;同时F44/GadB1C1 的发酵液也可以显著降低草莓腐烂率,比单独采用Nisin 具有更好的品质保持效果,与添加苯甲酸钠和山梨酸钾的防腐效果相当[77]。
3.3.4 设计益生菌活性包装
益生菌的活力大小以及其活力是否可以在贮藏期间得到保持是影响其对食源性致病微生物品质控制效果的一个重要因素。 可考虑将益生菌和大分子可食用材料相结合来达到保证其活力的目的,因此开发含有益生菌的活性包装产品也成为目前益生菌在保鲜领域应用和活性包装材料研发的热点。 该技术在保证益生菌具有抑制食源性致病微生物活力的同时还可以避免将益生菌直接作用于食品对食品的风味可能造成的影响,同时这种活性包装材料还具有保水、阻隔、抗氧化等性能。 有研究制作了益生菌植物多糖活性包装膜,并将其应用于水果和蔬菜的品质控制。 该方法利用木瓜、亚麻和罗勒的种子(含有不同多糖)包裹益生菌,制备了有高保湿性、热稳定性和高机械强度的可食用膜,发现这种可食用膜不仅具有很强的阻隔性,并能同时保证鼠李糖乳杆菌GG 的活力;用其包装草莓、香蕉、黄瓜和圣女果等鲜果可使鲜果免受紫外线照射,保持水分(黄瓜和圣女果水分的保持分别能持续7 d 和30 d)、维持良好外观以及避免褐变等[78]。 另有研究制作了含有多种植物乳杆菌(植物乳杆菌LP3、植物乳杆菌AF1 和植物乳杆菌LU5)的魔芋基可食用膜,对鲜切猕猴桃品质及其真菌进行控制。 该研究发现,随着贮藏天数的增加,可食用膜中的益生菌仍保持较高的存活率(7 lg CFU/g 左右),且植物乳杆菌LP3 的存活率要高于其他菌株;与对照组相比,鲜切猕猴桃的腐烂和颜色变化得到了很好的控制,霉菌和酵母菌的数量显著减少,叶绿素和抗坏血酸的含量基本没有发生变化,且总酚含量和抗自由基活性也比对照组高,其感官的接受度也更高,说明其食用安全性和品质都得到了很好的保持[79]。
3.3.5 联合其他食品安全和品质控制技术
益生菌及其代谢物还可以和其他食品安全和品质控制技术结合达到协同增效的效果。 研究发现,使用益生菌和EDTA 及黑胡椒提取物复配后,相对于分别单独采用罗伊氏乳杆菌、干酪乳杆菌、EDTA和黑胡椒提取物,复配物可将对大肠杆菌的抑制能力提升2 倍,也使低温条件贮藏的牛肉中微生物数量显著降低,并可抑制牛肉在贮藏过程中的脂肪氧化,保证牛肉的食用安全和品质[80]。 利用超高压技术结合乳酸乳球菌对肉类进行处理后可有效减少变形菌门类腐败细菌的数目,可以在很大程度上减少和避免亚硝酸盐的使用,降低消费者食用后的致癌风险[81]。 与其他技术的联用扩大了益生菌及其代谢物在食品安全和品质控制中的应用范围。
4 展望
基于益生菌及其代谢物的食源性致病微生物控制技术在食品安全和品质控制上的应用具有很大优势,但其工业化仍然面临很多问题。 首先要考虑的就是益生菌生物及其代谢物的食用安全性问题,目前已经批准在食品中添加的益生菌及其代谢物的种类较少,且研究发现某些抑菌物质对人体内正常益生菌群存在抑制作用,他们之间的关系也需要进一步研究。 未来的研究需要侧重对益生菌及其代谢物的安全性评价,为其工业应用提供理论基础和依据;同时还需要进一步对具有益生和抑制食源性致病微生物功能的益生菌菌株资源进行挖掘,结合生物信息学、计算生物学、数据科学和现代仪器分析等手段确定合适的分离培养方案。 益生菌抑菌活性物质的确定和分离也是目前研究的一个瓶颈,需要开发敏感度更高和更有效的识别和分离方法;同时相关菌株及其代谢物抑制食源性致病微生物的机制、环境对益生菌抗菌性能影响等机制也有待于进行更深层次的研究,为更好地实现基于益生菌及其代谢物的食源性致病微生物控制技术的开发应用提供理论依据。
