文山石漠化区坡柳水分利用效率与茎导管性状对坡位的响应
2022-05-27张宗领
张宗领,陈 静
(文山学院 冶金与材料学院,云南 文山 663099)
茎叶关系是植物生态学策略的主要维度之一,反映了植物在水分传输和光拦截之间的适应性选择[1],与植物碳水经济学密切相关[2]。在茎与叶的数量、大小关系中,叶片均依赖于茎的多种功能需求[2],因而茎数量-大小的变化能直接体现叶大小的优化目标,即达到水分利用效率最大化程度[3]。水分从植物主干传输到叶片层须经过茎木质部[4],茎木质部导管起支撑叶片和输导水、营养和同化物质的作用,是植物进行水分长距离运输的一条主要通道[5],其数量(茎导管密度)和大小(茎导管直径)性状决定了叶片的水分供应状况[6],故茎导管密度-大小表征了茎水力结构的变异。植物叶片水分利用效率是净光合速率(Pn)与蒸腾速率(Tr)之比,将植物碳循环和水循环结合起来,能解释植物内在生理的耗水机制[7]。植物导管内径越大,其木质部导水率越大[8],茎导管数量响应叶片气孔,对光合作用做出适时调整,使呼吸速率最小化、光合速率最大化,实现高效WUE以适应特定的生长环境。因此,WUE与茎导管性状的关系可反映植物体内碳水代谢的适应模式。坡位是制约土壤理化性质和小气候变化最重要的地形维度[9],从而影响着植物茎叶水力性状之间的平衡[10]。随坡位上升,土壤的厚度及营养物质、植冠微生境存在明显差异,为了维持SPAC系统终端部分叶片水分利用效率与茎导管有效供水之间的动态平衡,植物不仅调整茎的机械支持能力,而且还控制茎木质部导管结构变化,保证植物茎叶间水分传导的有效性与安全性[11],所以,研究不同坡位植株WUE与茎导管性状的关系对于探索植物输导系统对生境的适应机制具有重要意义。
坡柳(Dodonaea viscosa)又名车桑子,为热带和亚热带常绿灌木,耐干旱、耐贫瘠和适应性强,在水土保持和植被恢复工程中具有重要的生态价值。近年来,国内外学者对茎叶水力结构与环境因子的关系进行了较多分析[12-15],解释了茎叶水力结构在不同环境中的安全性与有效性。目前对坡柳的研究多集中在种群特性[16]、个体生长发育[17]、叶性状[18-19]、旱生解剖结构[20]等方面,解释了坡柳种群、个体、根茎叶对干旱胁迫和养分胁迫的响应特征,但缺少茎叶间的碳水传输效率研究,特别是WUE与茎导管密度、茎导管直径的关联性研究鲜少报道。本文通过探讨云南文山石漠化区不同坡位坡柳WUE和茎导管水力结构的变化,旨在理解植物光合效率和生物物理结构的关系,揭示石漠化地区茎叶碳水代谢的投资机理。
1 研究地区和研究方法
1.1 研究区概况
研究区位于云南文山东山乡石漠化地区(23°54'N,104°58' E),海拔1 315~ 1 788 m。年均温16.11 ℃,年均降水量989.30 mm左右,年均蒸发量1 780.20 mm,相对无霜期309 d。土壤以赤红壤和红色石灰土为主,pH值4.5~5.5。植被分布具有明显的垂直分异性,以灌木林和旱生草本植物为主。灌木植物主要有坡柳(Dodonaea viscosa)、少花新樟(Neocinnamomum delavayi)、女贞(Ligustrum lucidum)、紫叶李(Prunus Cerasifera)、黄杨(Buxus sinica)、沙针(Osyris lanceolata)、白刺花(Sophora davidii)、青篱柴(Tirpitzia sinensis)、铁仔(Myrsine Africana)、云南蛛毛苣苔(Paraboea neurophylla)、酸藤子(Embelia laeta);草本植物主要有白茅(Imperata cylindrica)、马缨丹(Lantana camara)、鼠尾草(Salvia japonica)、羊耳菊(Inula cappa)、菝葜(Smilax china)、野拔子(Elsholtzia rugulosa)、类芦(Neyraudia reynaudiana)、金丝草(Pogonatherum crinitum)等。
1.2 样地设置与调查方法
于2020年8月10~20日,根据实地考察,在文山市东山乡东北部一个独立的石质山地,选择一个生长着坡柳人工同龄林的典型坡面,研究坡面为东南坡向,坡长310 m,平均坡度25.7°,自上而下设置上坡、中坡、下坡3个样地,每个样地划分成5 m × 5 m的样方,上、中、下坡位样地的样方数分别为28、43、66,总计137个样方,对所有样方进行林分特征调查(表1)。每一坡位随机选择10个样方,每个样方各选取大中小1株,共3株坡柳,在其植冠中部外侧东、南、西、北4个方位各选择1根枝条,每根枝条上标记3片生长良好的叶片后,进行以下步骤。

表1 坡柳样地的林分特征
1.3 测试项目和方法
1.3.1 环境参数的测定
选择晴朗天气的6:00~18:00,首先用手持光量子计(3415 F,Spectrum technologies,Aurora,USA)对每个供试植株的上方、中层和地表处进行光合有效辐射(PAR)测定,然后在相同位置使用手持气象站(NK4500,Kestrel,New York,USA)进行冠层温度(Ta)、相对湿度(RH)数据的采集,每2 h测量一次,计算坡柳PAR、Ta和RH的平均值,依据Ta和RH得到大气水汽压亏缺[21]。最后在各个样地进行土壤取样,重复3次,采用烘干法测定土壤含水量。
1.3.2 光合生理参数测定
选择与环境参数测定的相同时间,利用GFS-3000便携式光合测量系统(Heinz Walz GmbH,Bavaria,Germany)进行每个标记叶片Gs、Pn、Tr的测定,并用便携式激光叶面积仪(CI-202,CID,Camas,USA)测定叶面积,最后将叶片烘干至恒质量,记录叶干重。水分利用效率WUE=Pn/Tr;叶含水量(%)=(鲜叶重-干叶重)/干叶重。
1.3.3 茎水力学结构测定
从每个选定的枝条上各截取长度为1~2 cm左右的茎段作为样品,置于装有FAA固定液的小瓶进行固定保存。经乙醇对样品逐级脱水后,再通过二甲苯透明和石蜡包埋进行处理,每个样品采用切片机(Leica CM1900,Wetzlar,Germany)切取3个完整的横切面,将切片进行番红-固绿双重染色后用中性树胶封片,制成茎导管制片[22]。在光学显微镜(SMZ168-BL,Motic,Hong Kong,China)下观察并拍照,用木细胞图像分析系统(WinCELL Regent Instrument,SainteFoy,Canada)计算坡柳茎木质部导管特征(导管密度、导管直径、导管壁厚度和机械强度)。
1.4 数据分析
本研究中所有原始数据全部采用Microsoft Excel 2013整理。3个坡位坡柳植株性状平均值的差异比较采用单因素方差分析(α=0.05);茎导管密度、茎导管直径、WUE数据均经log10转换,然后由SMATR Version 3.0软件完成标准化主轴估计(SMA),由SPSS 20.0及oringin 8.5软件进行数据统计和绘图。
2 结果与分析
2.1 不同坡位坡柳群落的环境特征
不同坡位坡柳群落的环境因子的变化如表2所示。单因素方差分析显示,随坡位的下降,空气相对湿度和土壤水分含量呈逐渐增大趋势,分别增加了23.08%、36.39%,二者在中坡位II和下坡位III均无显著差异(P>0.05);光合有效辐射、气温和大气水汽压亏缺呈逐渐减小趋势,分别减小了46.97%、12.71%和25.93%,气温和大气水汽压亏缺在中坡位II和下坡位III无显著差异(P>0.05)。表明低坡位的温湿度、水汽压亏缺和土壤水分状况优于高坡位,但光照条件变差。

表2 不同坡位坡柳群落的环境特征(平均值±标准误差,n=30)
2.2 不同坡位坡柳的叶性状变化
如表3所示,不同坡位坡柳叶片功能性状存在显著差异(P<0.05)。由上坡位I依次降到下坡位III,叶面积、叶片含水量、气孔导度、Pn和Tr均呈递增趋势,处于中坡位II的坡柳较上坡位I分别增加了27.63%、13.18%、45.83%、15.52%和32.90%,下坡位III的坡柳较中坡位II分别增加了17.45%、7.09%、17.14%、7.84%和16.94%。由此看出,低坡位植物单叶光合面积和水分含量大于高坡位,并且光合能力也更强。

表3 不同坡位坡柳的叶性状(平均值±标准误差,n=30)
2.3 不同坡位坡柳茎导管性状与水分利用效率
不同坡位坡柳的WUE和茎导管性状存在显著差异(P<0.05,表4)。从上坡位I到下坡位III,坡柳WUE、茎导管密度、茎导管壁厚度和导管壁机械强度呈递减趋势,中坡位II坡柳较上坡位I分别减少了13.43%、11.96%、11.58%和15.79%,下坡位III坡柳较中坡位II坡柳分别减少了8.62%、19.92%、20.89%和28.13%;茎导管直径呈递增趋势,中坡位II坡柳较上坡位I增加了15.95%,下坡位III的坡柳较中坡位II增加了10.82%。可见,WUE与茎导管密度、茎导管壁厚度和导管壁机械强度存在一致性的变化规律,而与茎导管直径有着相反的变化趋势。
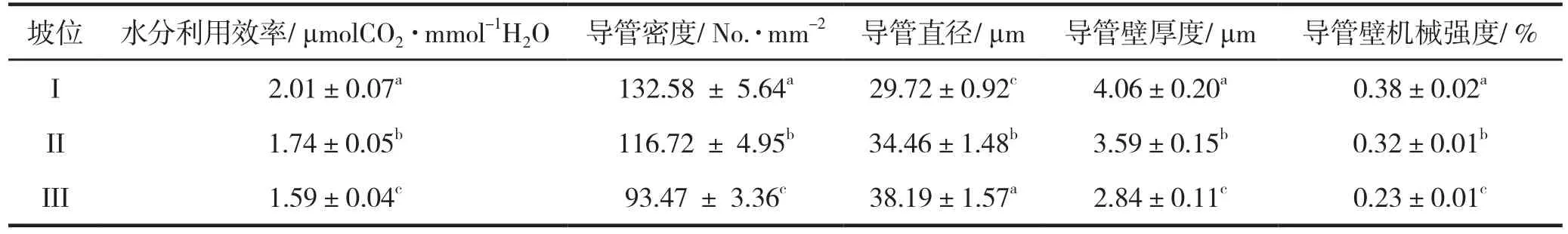
表4 不同坡位坡柳水分利用效率与茎导管性状(平均值±标准误差,n=30)
2.4 不同坡位坡柳水分利用效率与茎导管密度的关系
如图1所示,不同坡位的坡柳WUE和茎导管密度呈显著的正相关关系(P<0.05)。从SMA分析得到,坡柳在不同坡位间的SMA斜率存在差异,上坡位样地I为0.76(95%的置信区间:CI=0.59,0.81)、中坡位样地II为0.94(95%的置信区间:CI=0.86,0.99)、下坡位样地III为1.26(95%的置信区间:CI=1.02,1.49)。从上坡位I到下坡位III,SMA斜率依次呈现逐渐增大的趋势,上坡位I的SMA斜率显著小于1.0(P<0.05),中坡位II的斜率与1.0无显著差异(P>0.05),下坡位III的斜率显著大于1.0(P<0.05)。在不同坡位间存在着不同程度的异速关系,上坡位I的坡柳WUE的增加速度小于茎导管密度的增加速度;中坡位II接近等速关系;下坡位III的WUE增加速度大于茎导管密度的增加速度。表明在某一给定的WUE时,低坡位比高坡位坡柳对茎导管密度的投入减少。
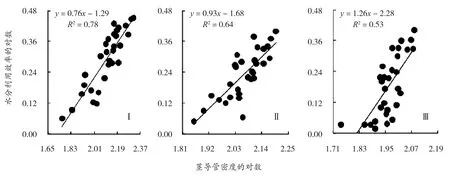
图1 不同坡位坡柳水分利用效率与茎导管密度的关系
2.5 不同坡位坡柳水分利用效率与茎导管直径的关系
如图2所示,不同坡位坡柳WUE和茎导管直径呈显著的负相关(P<0.05)。SMA分析显示,坡柳在不同坡位间的回归方程斜率存在明显差异,上坡位I为-1.20(95%的置信区间:CI=-1.24,-1.15)、中坡位II为-1.03(95%的置信区间:CI=-1.04,-0.97)、下坡位III为-0.64(95%的置信区间:CI=-0.70,-0.61),从I至III,SMA斜率的绝对值呈递减趋势,上坡位I的SMA斜率显著小于-1.0(P<0.05),中坡位II的斜率与-1.0无显著差异(P>0.05),下坡位III的斜率显著大于-1.0(P<0.05),即上坡位I坡柳WUE增加的速度大于茎导管直径减小的速度,中坡位II坡柳WUE增加的速度约等于茎导管直径减小的速度,下坡位III坡柳WUE增加的速度小于茎导管直径减小的速度。表明坡柳茎导管直径越大,WUE增加速率越小。
3 讨论
自然选择压力迫使植物叶片与茎之间始终维持着形态匹配和生物力学的动态平衡[2],茎通过形态生理结构调整来适应叶片水分运输的变化,确保异质环境中叶片光合与水分代谢间的协调统一[23]。本研究发现,对于每一坡位的坡柳,WUE与茎导管密度、茎导管直径间均存在不同的异速关系,WUE与茎导管密度呈显著的正相关(P<0.05),与茎导管直径呈显著的负相关(P<0.05)。这支持了Fan等[24]关于“植物较高的运输和输导能力需要有较粗的导管以及较低的导管密度”的发现。坡柳叶片WUE与茎导管性状的关系反映了茎导管水力结构对植物水分代谢的支持效率,以及维持碳水循环过程的生态机制。
3.1 上坡位坡柳水分利用效率与茎导管性状的关系
叶是茎端分生组织产生的侧生器官,其形态建成和生理功能受茎顶端生长和分化的影响[25]。坡柳群落居于上坡位时,生存空间和PAR增大,由于土层浅、被山体遮挡少的缘故,环境条件表现为强光、高温和干旱,坡柳增加茎导管密度,减少茎导管直径的资源分配,形成WUE的增加速度小于茎导管密度的增加速度,而大于茎导管直径减小的速度的异速关系。主要原因有:(1)强光和高温带来过强的光生物化学反应,严重限制了植物细胞的分裂和生长,使之面临光抑制的风险,为了自我遮阴需要,坡柳减小叶面积的生物量分配而将更多的资源投入于叶片数量的增长。一方面,小而厚的叶片往往具有薄的叶边缘,易与外界环境进行热量和物质交换,减轻光环境胁迫;另一方面,大量厚重的叶片造成单位枝茎的机械负载增大,继而导致茎木质部密度和单位导管数量的增加。(2)大气水汽压亏缺较大,坡柳茎导管内存在极低的负压容易产生崩裂,植物试图减少叶片含水量,增大水蒸气进出表皮阻力[26],从而降低茎、叶导水率,同时减小茎导管直径的资源分配以增大细胞壁厚度和机械强度,维持植物体导水系统的稳定性与安全性,利于其在胁迫环境下更具生存优势[27]。(3)该坡位土层浅薄,土壤含水量较低,叶片吸收水功能受抑增加了细胞脱水伤害风险[28],为减少植物体的水分散失,坡柳增加木质部导管密度使得水分传导可选择的路径增多,通过淘汰运输不畅的气蚀化木质部管道,经由安全路径将水分输送到蒸腾部位的细胞[29],从而实现以最小的水分散失来获得最大的碳同化速率[30],导致WUE上升。综上所述,WUE与茎导管性状间的相关性体现了先锋植物重要的生态学意义,处于上坡位的植物同时面临着光胁迫和干旱胁迫,坡柳为避免光抑制和水分亏缺的风险[31],通过高投资慢生长,保持高效的水分利用率,达到茎叶水分运输通道安全性的目的。
3.2 下坡位坡柳水分利用效率与茎导管直径的关系
茎作为根与叶片连接的通道,在植物对光、水资源的获得和能量平衡等方面具有重要作用[32]。研究显示,下坡位空气温湿度和土壤水分状况良好,但光照条件变差,坡柳趋向于WUE增加速度大于茎导管密度的增加速度,而小于茎导管直径减小的速度。原因在于:(1)随着坡位下降到III,山体遮挡时间渐长,加上坡柳个体相对高大,冠层内枝叶的自我遮荫现象严重,造成PAR急剧下降,光驱动力减弱限制了叶片对环境资源的可获得性,植株为了最大限度截获光辐射,趋向于简化导管结构,降低茎导管密度,将有限资源更多投入于叶片的拓展,增大了叶片捕光面积,利于光的穿透和光能的吸收[33],抵御弱光环境胁迫。(2)样地III浓密树冠增大了对雨水的截流作用,且减少了土壤表层的无效蒸散,有效改善植物种群的土壤水分条件[23],加上大气水汽压亏缺适中,使叶片始终保持着较高的含水量,这些都有助于避免植物体导管内连续水柱断裂的风险,即使导管壁薄、机械抗压强度低的大导管也不至于发生气穴化和阻塞,坡柳选择增加茎导管直径,增大叶片气孔导度和蒸腾速率,提高植物水分运输的有效性以促进植物光合能力。(3)该样地坡柳株高相对较高,势必要求植物产生较大的蒸腾拉力和Tr,以克服高生长带来的额外阻力和重力,同时大而薄的叶片拉近了CO2和水分等从气孔到达叶绿体的距离,缩短了水分运输时间,获得较高Tr,使得WUE下降。因此,石漠化区的植物随着坡位的递减,解除了叶片光合作用的水分胁迫,坡柳选择较低的茎导管密度也能满足蒸腾作用中的水分需求。植株降低茎导管结构的复杂性,扩大茎导管直径,以牺牲WUE为代价满足低坡位植株较高的水分需求量,保证水分供给的有效性,提高了整个植株的光合收益。
植物对水分防御和输导构造上的资源权衡投资直接决定了叶片水分代谢及其效率[34]。中坡位是由上坡转向下坡的生态交错区域,群落小气候、地表土壤性质存在着明显过渡性质,种间的空间、光照、水分等资源竞争相对缓和,坡柳拥有相对适宜的种群生态位,Gs、Pn和Tr均居中等,增强了茎木质部水分运输的自我调节能力,可以兼顾水分输导的有效性和安全性,达到提高水分利用效率的目的,坡柳采取折中茎导管数量和茎导管大小的均衡适应模式,不仅有效地拓展植物个体生存空间,增加了植株的光合收益,而且有利于缓解或避开无谓的种内竞争,维护种群的稳定。因此,生长在中坡位的坡柳WUE增加速度接近茎导管密度的增加速度,而约等于茎导管直径减小的速度,反映了植物在水分运送和输导结构上的稳定性生存策略。
4 结论
植株水分利用效率和茎导管性状之间的关系存在明显坡位差异。处于高坡位的坡柳植株趋向于减少茎导管直径的资源分配形成大量细径的茎导管,通过结构复杂的管道维持水分运输路径的安全性,提高水分利用效率来应对环境胁迫,这是植株所选择的一种高投入慢生长的保守性生存策略;生长在低坡位的植株构建少量粗径的茎导管,确保植物水分运输的有效性,从而最大限度地增加植物的光合收益,这源于坡柳采取的低投入快生长的竞争性生存策略。茎叶间物质与能量传输通道的耦联,体现了植物碳—水投资器官如何协调水力结构和光合效率的匹配问题。水分利用效率与茎导管性状的关系可能还受到区域气候背景、土壤性质和物种遗传特性的影响,仍需进一步的深入研究。
