水稻耐低氮基因OsTPP6的鉴定及功能分析
2022-05-21任秋韵田云录刘晓岚余珺唐伟杰陈高明林之希李静王春明万建民
任秋韵,田云录,刘晓岚,余珺,唐伟杰,陈高明,林之希,李静,王春明*,万建民,2
(1.南京农业大学作物遗传与种质创新国家重点实验室/江苏省现代作物生产协同创新中心,江苏 南京 210095; 2.中国农业科学院作物科学研究所,北京 100081)
目前我国氮肥的施用量一直居高不下,但仅有30%~50%的氮肥能被水稻吸收,其余的以N2O的形式排放到大气中,或者渗入水系统中造成水质富营养化[1-2],对环境产生极为不利的影响。此外,过量的氮肥对水稻产生诸多不利的影响,如抗折力下降,节间抗倒伏能力降低[3],单穗籽粒数、结实率和千粒重下降,单产降低[4],稻瘟病[5]、纹枯病[6]发生增加等。因此,挖掘水稻高效利用氮肥的新基因具有重要的应用价值。
氮高效利用是一个复杂的数量性状,由多基因以及环境因素共同控制[7]。目前,已有几种硝酸盐转运蛋白基因成功克隆并运用于水稻氮高效利用性状的改良[8-12]。海藻糖(trehalose)是一种稳定的非还原性二糖,已知的海藻糖生物合成途径至少有5种,但在植物中唯一存在的则是海藻糖 6-磷酸合成酶/海藻糖 6-磷酸磷酸酶(TPS/TPP)途径,它涉及TPS和TPP催化的2个步骤,TPS催化葡萄糖从UDP-葡萄糖(UDP-glucose)转移到葡萄糖6-磷酸(glucose 6-phosphate,G6P)形成海藻糖6-磷酸(T6P),然后TPP将T6P去磷酸化形成海藻糖[13]。植物中的TPP家族高度保守,对TPP功能的研究较少,主要与逆境胁迫相关[14-16]。拟南芥中的TPP被证明与硝态氮有关,在低氮和无氮胁迫下,AtTPPB、AtTPPJ、AtTPPF、AtTPPG、AtTPPH的表达均明显下调[17-20]。拟南芥中过表达TPP基因降低了植株体内T6P的水平,叶片中的AGPase氧化还原激活下降,淀粉含量下降[21]。但是,T6P不是通过影响AGPase来调节淀粉的合成和分解,而是通过调节植物的生物钟进行的[22],关于T6P影响淀粉代谢的机制尚不明晰。
为了发掘自然群体中影响水稻氮高效利用的优异等位基因,本研究通过全基因组关联分析和单倍型分析,鉴定了一个水稻氮素利用相关基因OsTPP6。OsTPP6响应硝态氮信号,影响硝态氮的吸收,该基因的突变增加了籽粒垩白率,使得淀粉合成相关基因表达上调。研究该基因功能为提高氮素利用效率改良籽粒品质提供理论依据。
1 材料与方法
1.1 试验材料
以南京农业大学作物遗传与种质创新国家重点实验室保存的117个不同来源的对氮素响应差异显著的水稻地方品种为材料,通过全基因组关联分析(genome wide association study,GWAS)挖掘氮素相关基因。117个品种包括55个粳稻、48个籼稻以及14个中间型材料,其中包括已报道的氮敏感品种‘南京 6号’和耐低氮品种‘千重浪2号’。所有供试材料于2019和2020年种植于南京农业大学土桥水稻育种基地,分正常氮田与低氮田种植,采用完全随机田间分组设计,正常田与低氮田各2组重复,每组每个地方品种种植4行,每行10株;T-DNA插入突变体种植野生型‘Dongjin’作为对照,正常田与低氮田各5组重复,每组种植4行,每行10株。正常田的基础氮水平为全氮1.86 g·kg-1,低氮田的全氮为1.40 g·kg-1。施用氮肥量正常田为300 kg·hm-2,低氮田为0 kg·hm-2,所有试验田的钾肥和磷肥的施用量均为100 kg·hm-2。OsTPP6基因T-DNA插入突变体由韩国庆熙大学作物生物技术研究所和基因工程实验室(https://orygenesdb.cirad.fr/cgi-bin/gbrowse/odb_japonica/)创制,插入的受体材料为‘Dongjin’(粳稻)。
1.2 试验方法
1.2.1 田间表型观测在水稻成熟期调查株高、有效分蘖数,去除边际效应植株,每组15株重复。通过计算低氮和高氮条件下的株高、有效分蘖数,将水稻生长发育的关键决定因素株高比(plant height ratio,PHR)和有效分蘖数比(effective panicle number ratio,EPNR)作为低氮耐受性指标。水稻收割后,考察穗长、粒长、粒宽、结实率、千粒重等农艺性状,每株3穗重复。
1.2.2 幼苗培养采用水培法,在人工气候室进行(16 h光照,30 ℃;8 h黑暗,20 ℃)。全营养素培养(包括硝酸盐和铵盐)至2叶1心期,在0、0.25、1.44、2.50 mmol·L-1KNO3营养液中继续培养10 d左右;取‘Dongjin’和OsTPP6T-DNA插入家系各15株,测量苗期的株高、地上部鲜重、地上部干重(鲜样置于 50 ℃烘箱1周烘干后测量)。其中用以氮饥饿转录组分析的幼苗,以尿素作为唯一的氮源培养2周后,将一半的幼苗置于无氮营养液中继续培养21 d后,取样。
1.2.3OsTPP6单倍型分析采用DNAsecure Plant Kit(版本号:DP171206)提取DNA。设计OsTPP6全长基因引物,在117个地方品种中扩增OsTPP6基因的全长序列。PCR产物测序结果通过Sequencher 5.4.5软件进行比对分析,得到117个地方品种中OsTPP6编码区的SNP,利用GraphPad Prism 8软件绘图。
1.2.4OsTPP6T-DNA插入片段检测采用CTAB法提取DNA,其中T-DNA检测的引物一端设计在基因上,另一端设计在插入片段上,用10 g·L-1琼脂糖凝胶电泳检测。
1.2.5 RNA的提取及反转录、实时荧光定量PCR用RNAprep Pure Plant Kit提取水稻根系RNA(淀粉相关基因定量提取花后18 d的籽粒RNA)。采用TaKaRa(大连)公司的反转录试剂进行反转录,得到 cDNA。用SYBR GREENProTaqHS预混型qPCR试剂盒,在ABI 7500定量PCR仪上进行基因的定量检测,以Actin(水稻微管合成蛋白基因)作为内参基因。所用引物见表1。

表1 检测、测序及定量所用的引物Table 1 Primers used in detection,sequencing and quantification
1.2.615N-KNO3吸收速率分析水稻幼苗在全营养素营养液中培养20 d后,在无氮营养液中饥饿培养3 d,然后转移至0.1 mmol·L-1CaSO4溶液中培养1 min,分别用0.25和2.50 mmol·L-1Ca(15NO3)处理 10 min,处理结束后放入0.1 mmol·L-1CaSO4溶液中培养1 min,最后置于50 ℃烘箱1周,完全烘干后磨样过0.28 mm筛,使用稳定同位素质谱仪 Delta V Advantage(Thermo Fisher ScientificTM)测定地上部、地下部的15N 含量。
1.2.7 籽粒直链淀粉含量的测定水稻成熟籽粒去壳后,使用球磨仪磨成精米粉(过0.25 mm孔径的筛),按照《米质测定方法:NY 147—1988》测定直链淀粉含量。
1.2.8 籽粒尿素膨胀体积的定性分析配制不同浓度的尿素溶液(0、1、2、3、4、5、6、7、8、9 mol·L-1),称取20.0 mg精米粉置于1.5 mL的离心管中,分别加入不同浓度的尿素溶液1 mL,25 ℃培养24 h,多次振荡混匀,待精米粉彻底溶解后8 000g离心30 min,静置30 min后观察并拍照。
2 结果与分析
2.1 OsTPP6的鉴定
对117个地方品种进行大田氮高效相关性状的全基因组关联分析,2015和2016年分别对EPNR和PHR这 2个水稻耐低氮品种的表型与分子数据进行关联分析,在8号染色体上检测到1个重复的位点。EPNR和PHR分别与S8_19349877(-lgP=3.35,R2=0.132)和S8 1917_3413(-lgP=4.30,R2=0.111)关联,该候选区间为897 kb,包含49个基因,294个导致氨基酸变异的SNP位点(图1-A)。候选区间内基因中仅有Os08g0400300、Os08g0409100、Os08g0410900和Os08g04146004个基因响应氮饥饿,Os08g0400300和Os08g0410900均被注释为编码假定蛋白的基因,Os08g0414600被注释为编码含有TolB类似结构域蛋白的基因,TolB结构域在革兰氏阴性菌中极度保守,在植物中尚未有报道。Os08g0409100被注释为编码海藻糖 6-磷酸磷酸酶基因6(OsTPP6),因此将OsTPP6作为氮素利用相关的候选基因(图1-B)。

图1 OsTPP6的鉴定Fig.1 Identification of OsTPP6 A.关联位点及LD block分析;B. LD block内基因的转录水平对低氮胁迫的响应。EPNR:有效分蘖数比;PHR:株高比。A.The associated sites and analysis of LD block;B. Transcription level of genes in LD block responded to low nitrogen stress. EPNR:Effective panicle number ratio;PHR:Plant height ratio.
2.2 OsTPP6单倍型分析
候选基因OsTPP6CDS区内存在3个SNP位点,分别是19578943 C/G、19578964 G/A、19579049 T/C,并且导致编码的氨基酸发生变化,第8位亮氨酸(Leu)变为缬氨酸(Val),第15位缬氨酸变为甲硫氨酸(Met),第43位缬氨酸变为丙氨酸(Ala)(图2-A),根据这3个SNP将关联群体分为2种单倍型(HapA、HapB)。HapA包括‘水原264’‘IR36’和‘JC92’等品种,HapB包括‘越光’‘小葱稻’和‘嘉花1号’等品种。HapA的株高比、有效分蘖比均显著高于HapB(图2-B),即HapA为一种优势单倍型。
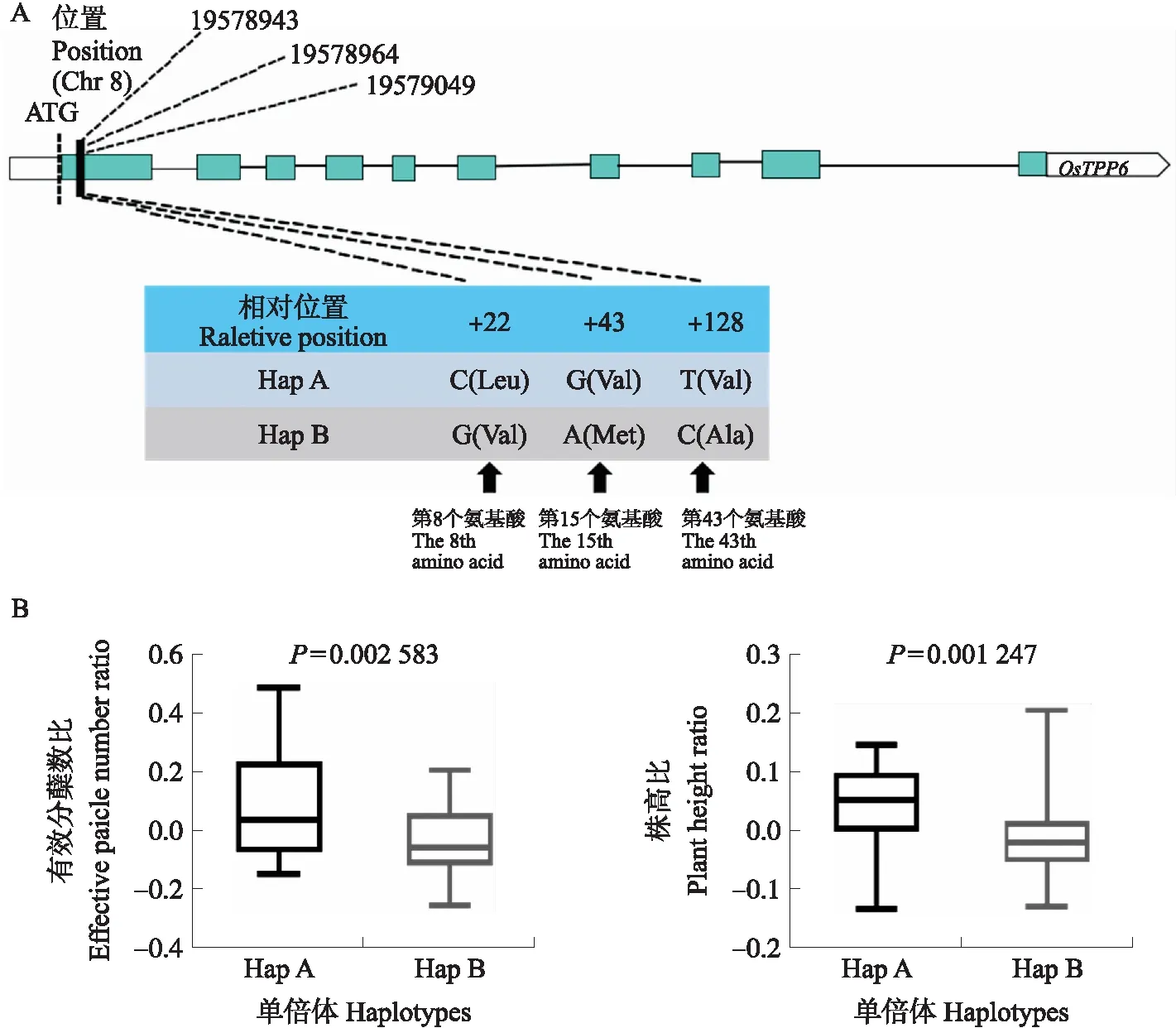
图2 OsTPP6的2种单倍型HapA和HapB分析Fig.2 The two haplotypes HapA and HapB analysis of OsTPP6 A. OsTPP6两种单倍型的SNP差异;B. OsTPP6两种单倍型的表型。差异显著性分析采用t测验。下同。A. The SNP differences between the two haplotypes of OsTPP6;B. Phenotype of the two haplotypes of OsTPP6. The significance analysis of the difference adopts the t test. The same below.
2.3 OsTPP6 T-DNA插入片段的验证
在野生型中可以扩增出OsTPP6基因全长的片段,但不能扩增出T-DNA插入部分片段,而在T-DNA插入突变体中,既可以扩增出T-DNA插入片段的部分片段,也可以扩增出OsTPP6基因上的片段(图3-A、B),推测该T-DNA插入突变体为杂合植株。T-DNA插入突变体中OsTPP6的表达水平相较于野生型显著下降(图3-C)。

图3 OsTPP6 T-DNA插入片段的验证Fig.3 Verification of OsTPP6 T-DNA insert A. T-DNA的插入位点和引物的设计;B. T-DNA插入位点的PCR检测;C.野生型和T-DNA插入突变体中OsTPP6转录水平。*P<0.05。A. T-DNA insertion site and primer design;B. PCR detection of T-DNA insertion site;C. Transcription level of OsTPP6 in wild type and T-DNA mutation.*P<0.05.
2.4 OsTPP6对氮素的响应分析
通过检测野生型与T-DNA插入突变体中氮素相关基因表达水平发现,铵态氮吸收利用相关的基因OsAMT1;2、OsAMT2;1、OsAMT2;2、OsAMT2;3、OsAMT3;1、OsAMT3;2(铵转运蛋白基因)、OsGDH1、OsGDH3(谷氨酸脱氢酶基因)、OsGOGAT2(谷氨酸合酶基因)、OsGS1;1、OsGS1;2(谷氨酰胺合成酶基因)的表达水平在野生型和 T-DNA 插入突变体中无明显差异,但与硝态氮利用相关的基因OsNIA2(硝酸还原酶基因)、OsNRT1.1、OsNRT2.1、OsNRT2.2(硝酸盐转运蛋白基因)的表达量在T-DNA插入突变体中均显著上调(图4-A),表明基因OsTPP6可能与硝态氮利用相关。

图4 野生型和OsTPP6 T-DNA插入突变体对硝酸盐的响应Fig.4 Wild type and OsTPP6 T-DNA mutation response to nitrate*P<0.05,**P<0.01. The same below.
在低浓度硝酸盐处理下,OsTPP6在处理2 h表达量显著上升(图4-B),而在高浓度硝酸盐处理下,OsTPP6在处理32 h表达量显著上升(图4-C)。表明低浓度硝态氮能迅速诱导OsTPP6表达。
2.5 OsTPP6 T-DNA插入突变体15N吸收速率分析


图处理10 min后野生型和OsTPP6 T-DNA插入突变体15N吸收速率分析Fig.5 Analysis of 15N uptake rates in wild type and OsTPP6 T-DNA mutation treatment for 10 min
2.6 OsTPP6 T-DNA插入突变体苗期表型分析
用0、0.25、1.44、2.50 mmol·L-1KNO3分别处理野生型和T-DNA插入突变体,在1.44 mmol·L-1KNO3处理条件下,野生型和T-DNA插入突变体长势最佳、均有最大生物量,且T-DNA插入突变体地上部生物量显著高于野生型;在较低浓度(0、0.25 mmol·L-1)硝酸盐处理时,T-DNA插入突变体的株高相较于野生型显著降低,但地上部生物量无明显变化;在高浓度(2.50 mmol L-1)硝酸盐处理时,野生型和T-DNA插入突变体的株高和地上部生物量均无差异(表2)。此外,在4种处理条件下,T-DNA插入突变体中OsNIA2的表达量均较野生型显著上调(图6)。以上结果进一步表明OsTPP6响应硝态氮信号,且负调控硝酸还原酶基因的表达。

图6 不同硝酸盐浓度处理条件下野生型和OsTPP6 T-DNA插入突变体的相对表达水平Fig.6 Relative expression level of wild type and OsTPP6 T-DNA mutation under different nitrate concentration treatments

表2 4种硝酸盐浓度处理下野生型和T-DNA插入突变体苗期表型Table 2 Phenotypes of wild type and T-DNA mutation under treatment with four nitrate concentrations in seedling stage
用野生型和T-DNA插入突变体不同浓度和零浓度硝酸根处理条件下中不同性状(株高、地上部鲜重、地上部干重)差值与零浓度硝酸根处理时各生理指标的比值(HN-LN/LN),来表征对氮素响应程度,反映氮素利用效率。发现在0.25和2.50 mmol·L-1KNO3处理中,T-DNA插入突变体对硝酸盐的响应均显著高于野生型,说明氮素利用效率提高,在1.44 mmol· L-1KNO3时,T-DNA插入突变体对硝酸盐的响应达到最大值(图7),表明OsTPP6负调控水稻对硝态氮的利用。

图7 野生型和T-DNA插入突变体的苗期生长指标对不同浓度硝酸盐的利用差距Fig.7 The difference in the utilization of nitrate at different concentrations of wild-type and T-DNA mutation seedling phenotype (HN-LN)/LN=(某硝酸盐浓度下的生长指标-零硝酸盐浓度下的生长指标)/零硝酸盐浓度下的生长指标。PH:株高;SFW:地上部鲜重;SDW:地上部干重。(HN-LN)/LN=(growth index under a certain nitrate concentration-growth index under zero nitrate concentration)/growth index under zero nitrate concentration. PH:Plant height;SFW:Shoot fresh weight;SDW:Shoot dry weight.
2.7 OsTPP6 T-DNA插入突变体大田表型分析
2.7.1OsTPP6T-DNA插入突变体在正常田中的表型分析在正常田中,T-DNA插入突变体的株高、有效分蘖数、穗长、粒长、千粒重、结实率相较于野生型均减小,粒宽则无明显差异(图8,表3)。在低氮田中,相比于野生型,T-DNA插入突变体也表现出株高、有效分蘖数、穗长、粒长、千粒重、结实率降低(图9,表3)。

图8 OsTPP6 T-DNA插入突变体在正常田的表型Fig.8 Phenotypes of OsTPP6 T-DNA mutation in normal field

表3 野生型和T-DNA插入突变体农艺性状Table 3 Agronomic traits of wild type and T-DNA mutation

图9 OsTPP6 T-DNA插入突变体在低氮田的表型Fig.9 Phenotypes of OsTPP6 T-DNA mutation in low nitrogen field
通过分析野生型和T-DNA插入突变体在正常田和低氮田中的表型,发现T-DNA插入突变体对氮素的响应比野生型强烈(图10),说明T-DNA插入突变体具有更高的氮素利用效率,与苗期结果基本一致(图7)。综上所述,OsTPP6的缺失增强了水稻对氮素的利用,提高氮素利用效率,但同样会影响水稻的生长和发育,多项农艺性状指标与野生型相比降低。

图10 野生型和T-DNA插入突变体对氮素的响应Fig.10 Responses of wild-type and T-DNA mutation to nitrogen (HN-LN)/LN=(正常田的性状-低氮田的性状)/低氮田的性状。(HN-LN)/LN=Calculation of the response of different phenotypes to nitrogen=(traits under normal field-traits under low nitrogen field)/traits under low nitrogen field.PH:株高 Plant height;EPN:有效分蘖数 Effective panicle number;TGW:千粒重 Thousand grain weight;SSR:结实率 Seed setting rate.
2.8 OsTPP6 T-DNA插入突变体籽粒的表型、理化性质分析及淀粉代谢相关基因的差异表达
与野生型相比,T-DNA插入突变体的垩白明显增多,籽粒胚乳呈粉质、不透明状,而野生型籽粒胚乳则呈现透明状(图11-A)。T-DNA插入突变体的直链淀粉含量相较于野生型极显著降低(图11-B);当尿素浓度为5 mol·L-1时,野生型籽粒中的淀粉发生明显膨胀,而T-DNA插入突变体则无明显变化(图11-C),表明OsTPP6的缺失改变了成熟籽粒中淀粉相关的理化性质。此外,淀粉代谢相关基因OsAGPL、OsAGPS(ADP-葡萄糖焦磷酸化酶基因)、OsGBSS(直链淀粉合成酶基因)、OsTPT(磷酸丙糖转运载体基因)、OsISA(异淀粉酶基因)、OsGWD(葡聚糖水合二激酶基因)在T-DNA插入突变体中均显著上调(图11-D),表明OsTPP6参与淀粉代谢,进而影响籽粒的品质。

图11 OsTPP6 T-DNA插入突变体籽粒的表型、理化性质及淀粉代谢相关基因的差异表达分析Fig.11 The analysis of phenotype,physical and chemical properties grains and differential expression analysis of genes related to starch metabolism in OsTPP6 mutation A.野生型及T-DNA插入突变体外观表型;B. 野生型和T-DNA插入突变体籽粒中直链淀粉的含量;C.野生型和T-DNA插入突变体籽粒淀粉在不同浓度尿素中的膨胀体积,红框表示尿素浓度为5 mol·L-1;D.野生型和T-DNA插入突变体中淀粉代谢相关基因的表达情况。A. Grain appearance of wild type and T-DNA mutation;B. Amylose content of wild type and T-DNA mutation;C. The starch expansion volume of wild type and T-DNA mutation in different concentrations of urea,the red box indicates the 5 mol·L-1 urea concentration;D. Expression level of genes related to starch metabolism in wild type and T-DNA mutation.
3 讨论
TPP作为海藻糖生物合成途径中的关键酶,目前在水稻中的相关研究主要围绕干旱、寒冷、厌氧等逆境胁迫展开[23-26]。同时水稻中关于氮素利用效率(nitrogen-use efficiency,NUE)的研究大多集中在双亲本QTL的检测和同源基因的克隆。并且发现Aus and Aromatic稻中存在的OsTCP19-H是调控水稻分蘖响应氮素的关键基因[27]。本研究中基于全基因组关联分析,利用2年的大田表型数据鉴定了一个海藻糖 6-磷酸磷酸酶基因(OsTPP6),作为NUE的候选基因。通过分析野生型和T-DNA插入突变体中氮素利用相关基因的表达差异、15N吸收速率差异,明确了该基因对硝态氮的响应,表明OsTPP6参与水稻硝态氮利用的相关途径。
本研究通过比较野生型和T-DNA插入突变体对不同浓度硝态氮的利用差异,发现T-DNA插入突变体对硝态氮的利用水平显著高于野生型。连续2年的大田试验也发现,在施用相同氮肥的条件下,T-DNA插入突变体的株高、有效分蘖数、千粒重的增加量均显著高于野生型,OsTPP6的缺失增加了水稻对氮素的利用,具有提高NUE的潜力。
T6P可能是通过激活ADP-葡萄糖焦磷酸化酶(AGPase)的氧化还原活性来调节淀粉的合成[21],它是植物体内一种重要的生长调节物质,在白天T6P通过调控PEP羧化酶翻译后修饰影响光合同化物在蔗糖、有机质和氨基酸之间的分配;而在夜晚调控叶片(源)中瞬时淀粉的降解,以及种子等(库)中淀粉的合成、降解及转运以满足植物生长对蔗糖的需求[28]。本研究发现,OsTPP6T-DNA插入突变体籽粒胚乳呈粉质、不透明状,垩白增多。垩白的形成与淀粉的代谢异常有关,通过分析淀粉代谢相关基因表达水平发现,在T-DNA插入突变体中,OsAGPL、OsAGPS、OsGBSS、OsISA和OsGWD的表达均显著上调,表明OsTPP6的缺失导致淀粉代谢途径相关基因的表达紊乱,淀粉代谢异常。同时,直链淀粉含量和尿素膨胀体积的差异也表明淀粉代谢异常,导致水稻米粉的理化性质出现了差异。OsTPP6的缺失影响了水稻籽粒中淀粉的正常代谢。
本研究通过对野生型和OsTPP6T-DNA插入突变体硝酸盐15N吸收速率分析以及NRT家族表达水平的测定,发现OsTPP6负调控水稻对硝态氮的吸收。在不施硝酸盐条件下OsTPP6T-DNA插入突变体表现出明显生长缺陷的表型(株高变矮,地上部干重减少),而随着硝酸盐的加入,可以弥补OsTPP6缺失导致的生长缺陷表型,并且在高浓度硝酸盐时生长状态要优于野生型的表型。而田间条件复杂,OsTPP6的缺失导致T6P的代谢紊乱,使OsTPP6T-DNA插入突变体的农艺性状都较野生型差。
综上所述,OsTPP6作为一个海藻糖 6-磷酸磷酸酶,参与水稻硝态氮利用相关途径和籽粒淀粉代谢相关途径,对研究水稻高效利用氮素、籽粒品质改良具有一定的应用价值。
