葡萄糖酸铜和双硫仑对SARS-CoV-2 PLpro的抑制作用
2022-05-11柳晓春辛全娟赵广健袁文敏王一帆章思语徐锡明
王 娟,柳晓春,辛全娟,赵广健,袁文敏,王一帆,章思语,徐锡明
(1.中国海洋大学医药学院,山东 青岛 266003;2.青岛海洋生物医药研究院,山东 青岛 266100;3.日照市中心医院呼吸与危重症医学科,山东 日照 276800)
2019年底,新型冠状病毒肺炎暴发,并以迅猛的速度在世界范围内传播,对全球公民的健康造成很大影响[1]。新型冠状病毒通过人与人之间的飞沫或直接接触而传播,病毒感染后常见的症状是发热,并伴有咳嗽,呼吸困难,肌痛和疲劳[2]。SARS-CoV-2是单股正链RNA病毒,编码4种结构蛋白(刺突蛋白、包膜蛋白、膜蛋白和核衣壳蛋白)以及两种多聚蛋白(polyprotein 1a and polyprotein 1ab,pp1a和pp1ab),pp1a和pp1ab被两种蛋白酶:木瓜样蛋白酶(papain-like protein,PLpro)和3C样蛋白酶(3C-like protein,3CLpro)切割成16种非结构蛋白(non-structure protein,NSPs),这16种非结构蛋白是新型冠状病毒基因表达的关键功能性蛋白[3]。因此,PLpro和3CLpro是抗新型冠状病毒药物研发的重要靶点。PLpro是NSP3自裂解产生的一种非结构蛋白,可以水解病毒蛋白多聚体成为功能性的NSPs,在病毒复制过程中起关键作用。抗新型冠状病毒药物靶向PLpro不仅可以在病毒的复制中起作用,还可以抑制Ⅰ型干扰素的产生,从而抑制非特异性免疫反应[4]。另外,泛素(ubiquitin,Ub)和类泛素蛋白干扰素刺激基因(interferon-stimulated gene 15,ISG15)都在其C末端带有LXGG序列,PLpro能够识别和水解LXGG序列,将Ub和ISG15从宿主细胞蛋白中切除,帮助冠状病毒逃避宿主天然免疫反应[5]。
半胱氨酸蛋白酶是一类重要的蛋白酶家族,广泛参与人和动物的多种生理过程。半胱氨酸蛋白酶家族在结构上有着很高的同源性,其催化核心是一个三联体结构,影响半胱氨酸蛋白酶的蛋白水解活性,在决定酶的特异性方面起着重要作用[6]。PLpro是半胱氨酸蛋白酶家族的一类,具有半胱氨酸(Cys)、组氨酸(His)、天冬氨酸(Asp)催化三联体,其中半胱氨酸的巯基是酶活性的关键位点,化合物和巯基反应可抑制酶活性[7]。另外,新冠状病毒与严重急性呼吸综合征冠状病毒(SARS-CoV)、中东呼吸综合征冠状病毒(MERS-CoV)基因组有很高的同源性,尤其表达几种关键酶的基因非常相似,这为针对SARS-CoV和MERS-CoV的药物研究成果直接应用到针对新型冠状病毒的治疗中提供了理论依据。已有文献证明,金属离子和双硫仑可以抑制SARS-CoV和MERS-CoV的活性[8]。
铜元素是人体内必需的微量元素之一,各种组织的代谢都需要铜元素,缺铜易引起贫血、白细胞减少、生长缓慢等症状,目前临床上大都使用硫酸铜治疗缺铜症。半胱氨酸蛋白酶的巯基可以与金属铜离子形成配位键,金属铜已经被证明对半胱氨酸蛋白酶家族的木瓜蛋白酶有很好的抑制效果[9]。双硫仑(disulfiram,DSF)是一种常见的戒酒药,是已知的很好的硫醇反应化合物,可以共价修饰甲基转移酶,脲酶的半胱氨酸残基。双硫仑对MERS-CoV和SARS-CoV的PLpro蛋白分别表现出一定的抑制作用[8]。在体内,双硫仑在酸性条件下或其二硫键被还原时分解,生成两个二乙基二硫代氨,在甲基转移酶作用下,生成二硫代氨基甲酸甲酯,也可通过细胞色素P450介导的亚砜作用最终形成S-甲基-N-二乙基硫代氨基甲酸酯砜[10]。近年来,有研究发现,双硫仑是一种有效的抗癌药,双硫仑联合铜可以增强抗肿瘤效果,且双硫仑的体内代谢产物如二硫代氨基甲酸甲酯等联合金属铜离子在临床前模型中显示出抗癌活性,能够杀伤包括黑色素瘤、直肠癌、肺癌、脑胶质瘤、乳腺癌等多种肿瘤[11]。本研究针对葡萄糖酸铜和双硫仑对SARS-CoV-2的PLpro蛋白的抑制作用展开实验,且对二者联合使用及其代谢物对PLpro蛋白的影响做了进一步探索。
1 材料与方法
1.1 药品与试剂96孔板(Costar Corning Incoporated)、默克millipore 10 K 15 mL超滤离心管(上海未熹生物科技有限公司)、TB培养基(金克隆生物技术有限公司)、Ni Smart Beads 6FF(常州天地人和生物科技有限公司)、E.coli BL21(DE3)菌株(上海昂羽生物技术有限公司)、质粒小提中量试剂盒(天根生化科技有限公司)、考马斯亮蓝染液(南京凯基生物科技发展有限公司)、底物Ub-AMC和Cbz-RLRGG-AMC(合肥科生景肽生物科技有限公司)。
1.2 仪器多功能酶标仪(SpectraMax I3,Molecular Devices,LLC),震荡培养箱(上海知楚仪器有限公司),冷冻式离心机(SIGMA 3-30K,创合盛科教仪器有限公司),超声波细胞粉碎机(宁波新芝生物科技股份有限公司),NanoDrop微型分光光度计(Thermo Fisher Scientifics,Gene Company Limited基因有限公司)。
1.3 方法
1.3.1新型冠状病毒PLpro蛋白的表达纯化 新型冠状病毒PLpro蛋白的基因根据其原始编码序列中G/C含量和适用于大肠杆菌原核表达系统密码子的应用进行优化,优化后的编码核苷酸序列如下所示:3′-GAAGTGCGTACCATTAAGGTGTTTACCACCGTTGATAATATTAATCTGCATACCCAGGTTGTTGAT ATGAGCATGACCTATGGTCAGCAGTTTGGCCCGACC TATCTGGATGGTGCAGATGTGACCAAAATTAAGCCG CATAATAGCCATGAAGGTAAAACCTTTTATGTTCTGC CGAATGATGATACCCTGCGTGTTGAAGCATTTGAATA TTATCATACCACCGATCCGAGTTTTCTGGGCCGTTAT ATGAGTGCACTGAATCATACCAAAAAATGGAAATAT CCGCAGGTTAATGGCCTGACCAGTATTAAGTGGGCA GATAATAATTGTTACCTGGCAACCGCCCTGCTGACC CTGCAACAGATTGAACTGAAATTCAATCCGCCGGCA CTGCAAGATGCCTATTATCGTGCACGCGCAGGTGA AGCCGCCAATTTTTGTGCACTGATTCTGGCCTATTG CAATAAGACCGTTGGCGAACTGGGCGATGTGCGTG AAACCATGAGTTATCTGTTTCAGCACGCTAATCTGG ATAGTTGCAAACGTGTTCTGAATGTTGTGTGCAAAA CCTGTGGTCAGCAGCAGACCACCCTGAAAGGCGTG GAAGCCGTGATGTATATGGGCACCCTGAGTTATGAA CAGTTTAAAAAAGGTGTGCAGATTCCGTGCACCTGC GGTAAACAGGCAACCAAATATCTGGTTCAGCAGGA AAGCCCGTTTGTTATGATGAGCGCCCCGCCGGCCC AGTATGAACTGAAACATGGTACATTCACTTGTGCAA GCGAATATACCGGCAATTATCAGTGCGGTCATTATA AACATATCACCAGTAAAGAAACCCTGTATTGCATTG ATGGTGCCCTGCTGACAAAAAGTAGTGAATATAAAG GCCCGATTACCGATGTGTTTTATAAAGAAAATAGCT ACACCACCACCATTTAA-5′。
选择pET-28a(+)质粒作为表达载体,酶切位点为NdeⅠ-XhoⅠ,N端带6× His-tag,将测序正确的pET-28a(+)重组质粒转化至E.coliBL21(DE3)中,挑单克隆菌落至10 mL的LB培养基中37 ℃摇床培养过夜,再转移至1 L的体系中37 ℃摇床培养至OD600为0.8-1.0,加入异丙基硫代半乳糖苷(isopropyl β-D-thiogalactoside,IPTG)至终浓度为0.2 mmol·L-1,16 ℃诱导过夜表达;菌液4 500×g4 ℃离心30 min,弃上清用磷酸缓冲盐溶液(phosphate buffer saline,PBS)重悬菌体,再次4 500×g4 ℃离心30 min,弃上清收集菌体。用裂解液(20 mmol·L-1HEPES,0.5 mmol·L-1NaCl,pH 7.4,200 mg·L-1溶菌酶和0.05% Triton X-100)[12]重悬菌体至50 mL离心管,置于摇床上4 ℃孵育30 min后冰上超声破碎,1 0000×g,4 ℃离心取上清备用。用5倍柱体积的双蒸水清洗镍柱,10倍柱体积平衡液(20 mmol·L-1HEPES,0.5 mmol·L-1NaCl,10 mmol·L-1imidazole,pH 7.4)平衡,随后加入菌体上清,4 ℃孵育1 h,弃流出液,加入洗杂液(20 mmol·L-1HEPES,0.5 mmol·L-1NaCl,20 mmol·L-1imidazole,pH 7.4)洗杂蛋白,加入洗脱液(20 mmol·L-1HEPES,0.5 mmol·L-1NaCl,250 mmol·L-1imidazole,pH 7.4)洗脱并收集目的蛋白。通过SDS-PAGE、考马斯亮蓝染色方法分析各组分的纯度,收集纯度较高的蛋白至Millipore Amicon Ultra-15超滤管中离心浓缩除盐,即得纯化好的重组PLpro蛋白。
1.3.2PLpro米氏常数的测定 Ub-AMC和Cbz-RLRGG-AMC是去泛素化酶的一种荧光底物,AMC是一类稳定性好、位移性好的香豆素类荧光染料,底物与去泛素化酶孵育释放AMC,使得荧光值增加来检测酶的活性(Ex./Em.=360/460 nm),用这种方法能够评价去泛素化酶的活性或筛选去泛素化酶的抑制剂[13-14]。PLpro是一类去泛素化酶,在PLpro的作用下,Ub-AMC和Cbz-RLRGG-AMC水解产生去淬灭AMC,通过测定荧光强度的变化来评价PLpro的活性。在酶总浓度为100 nmol·L-1、荧光底物浓度为0.125~40 μmol·L-1的条件下,测定新型冠状病毒PLpro的酶活。新型冠状病毒PLpro活性测定的缓冲体系为:20 mmol·L-1磷酸盐缓冲液(pH 6.8)。在25 ℃的孵育温度下,加入100 nmol·L-1的PLpro,迅速加入荧光底物Cbz-RLRGG-AMC或Ub-AMC,每隔1.5 min时间记录一次荧光读数,共测定30 min。根据实验得到不同底物浓度下的反应速率值,运用GraphPad Prism 8中的Michaelis-Menten模型非线性拟合出Michaelis-Menten方程曲线,并得到拟合值Vmax和Km值,Michaelis-Menten方程式如下:
其中,Vmax表示酶被底物饱和时的反应速率,Km值表示米氏常数,是酶促反应速率V为最大反应速率一半时的底物浓度。
1.3.3抑制剂葡萄糖酸铜和双硫仑IC50值和Ki值的测定 测定葡萄糖酸铜和双硫仑IC50值的实验在黑孔96孔板中进行,总体系为100 μL,50 μL的PLpro的浓度为40 nmol·L-1,在25 ℃的孵育温度下,加入20 μL浓度为1~200 nmol·L-1的葡萄糖酸铜或0.02~10 μmol·L-1的双硫仑,室温震荡孵育15 min,迅速加入30 μL荧光底物Ub-AMC(2.4 μmol·L-1),进行化合物的IC50检测。每隔1.5 min记录一次荧光读数,共测定30 min。以葡萄糖酸铜浓度和双硫仑浓度的对数值为横坐标,对应的抑制率的值为纵坐标,用GraphPad Prism 8中的非线性回归(曲线拟合)模块中的四参数法制作曲线,并分析计算出葡萄糖酸铜和双硫仑的IC50值。用Dixon作图法求葡萄糖酸铜和双硫仑的Ki值。实验在黑色96孔板中进行,总体系为100 μL,50 μL的PLpro的浓度为40 nmol·L-1,在25 ℃的孵育温度下,分别加入20 μL浓度为0、25、50、100 nmol·L-1的葡萄糖酸铜或0、1.25、2.5、5 μmol·L-1的双硫仑,室温震荡孵育15 min,迅速加入30 μL荧光底物Ub-AMC(2.5 μmol·L-1或者10 μmol·L-1),每隔1.5 min记录一次荧光读数,共测定30 min。以不同的抑制剂浓度为横坐标,酶活反应速率的倒数为纵坐标作图,两种不同底物浓度的曲线交点的横坐标即为该抑制剂的Ki值。
1.3.4抑制剂葡萄糖酸铜和双硫仑的酶动力学研究 用Lineweaver-Burk作图法判断葡萄糖酸铜和双硫仑对PLpro的抑制作用类型[15]。实验在黑孔96孔板中进行,总体系为100 μL,PLpro总浓度为40 nmol·L-1,葡萄糖酸铜用去离子水分别配制成浓度为0、25、50、100 nmol·L-1的浓度梯度,双硫仑用去离子水分别配制成浓度为0、1.25、2.5、5 μmol·L-1的浓度梯度,在25 ℃的孵育温度下,分别加入不同浓度(5~80 μmol·L-1)的荧光底物Ub-AMC,对新型冠状病毒PLpro酶活进行测定。每隔1.5 min记录一次荧光读数,共测定30 min。根据实验结果,以底物浓度的倒数为横坐标,以反应速率的倒数为纵坐标绘制Lineweaver-Burk双倒数曲线,由此判定抑制剂的抑制类型。若曲线相交于Y轴,抑制剂的抑制类型为竞争性抑制剂,若曲线相交于X轴,则抑制剂的抑制类型为非竞争性抑制,若曲线相互平行不相交,则为反竞争性抑制剂[8]。
用Yonetani-Theorell作图法进一步探究葡萄糖酸铜和双硫仑对PLpro蛋白作用的相互影响。实验在黑孔96孔板中进行,总体系为100 μL,PLpro总浓度为40 nmol·L-1,将不同浓度的葡萄糖酸铜(0~500 nmol·L-1)和300 nmol·L-1的双硫仑联合使用,在25 ℃的孵育温度下,加入2.4 μmol·L-1荧光底物Ub-AMC,对新型冠状病毒PLpro酶活进行测定。每隔1.5 min记录一次荧光读数,共测定30 min。根据实验结果,以葡萄糖酸铜的浓度值为横坐标,以反应速率的倒数为纵坐标绘曲线,并通过两条曲线的关系得到评价两种抑制剂相互作用的常量α,其定义了一种抑制剂与蛋白的结合对另一种抑制剂对蛋白亲和力的影响。若曲线形成一系列平行线,则α=∞,表示两个抑制剂之间是互相排斥的,作用于同一结合位点;若曲线相交于原点,则α=1,表示两个抑制剂之间完全独立,互不影响;若曲线相交于X轴上方,则α<1,表示一个抑制剂和蛋白的结合增加了蛋白对另一个抑制剂的亲和力,两个抑制剂被认为是协同的;若曲线相交于X轴下方,则α>1,表示一个抑制剂和蛋白的结合降低了蛋白对另一个抑制剂的亲和力,两个抑制剂则被认为是拮抗的。
1.3.5双硫仑代谢物对PLpro的作用 选用DSF的代谢物二硫代氨基甲酸甲酯和S-甲基-N-二乙基硫代氨基甲酸酯砜作为研究对象,将其配置为10~1 500 μmol·L-1的阶梯浓度,测定新型冠状病毒PLpro的酶促反应的初速率,实验体系和条件与上述IC50值测定保持一致。
2 结果
2.1 PLpro蛋白纯化重组的PLpro蛋白在大肠杆菌中表达,经镍柱亲和层析纯化后,得到纯度较高的PLpro重组蛋白,通过SDS-PAGE检测蛋白纯度,结果如Fig 1所示,PLpro蛋白的纯度高,且得到的SARS-CoV-2 PLpro重组蛋白的分子量在35~40 ku之间,符合PLpro蛋白38.5 ku的理论分子量[16],纯化的蛋白储存在在50 mmol·L-1的Tris-HCl中,将其分装并于-80 ℃保存。

Fig 1 SDS-PAGE analysis of SARS-CoV-2 PLpro
2.2 PLpro米氏常数测定根据实验的得到不同的底物浓度下的反应速率值,运用GraphPad Prism 8中的Michaelis-Menten模型非线性拟合出Michaelis-Menten方程曲线并得到拟合值Vmax和Km值,如Fig 2所示,对于荧光底物Cbz-RLRGG-AMC,PLpro的米氏常数Km值为32.39 μmol·L-1,最大反应速率Vmax为1.26 nmol·L-1·s;对于荧光底物Ub-AMC,PLpro的米氏常数Km值为7.322 μmol·L-1,最大反应速率Vmax为0.519 nmol·L-1·s。实验结果表明对于两种不同的荧光底物,Ub-AMC与PLpro蛋白计算得到的Km值较小,因此,相比较Cbz-RLRGG-AMC,Ub-AMC有更好的亲和力,灵敏度更好,在后续的实验中选择Ub-AMC作为PLpro蛋白的底物进行一系列实验研究。
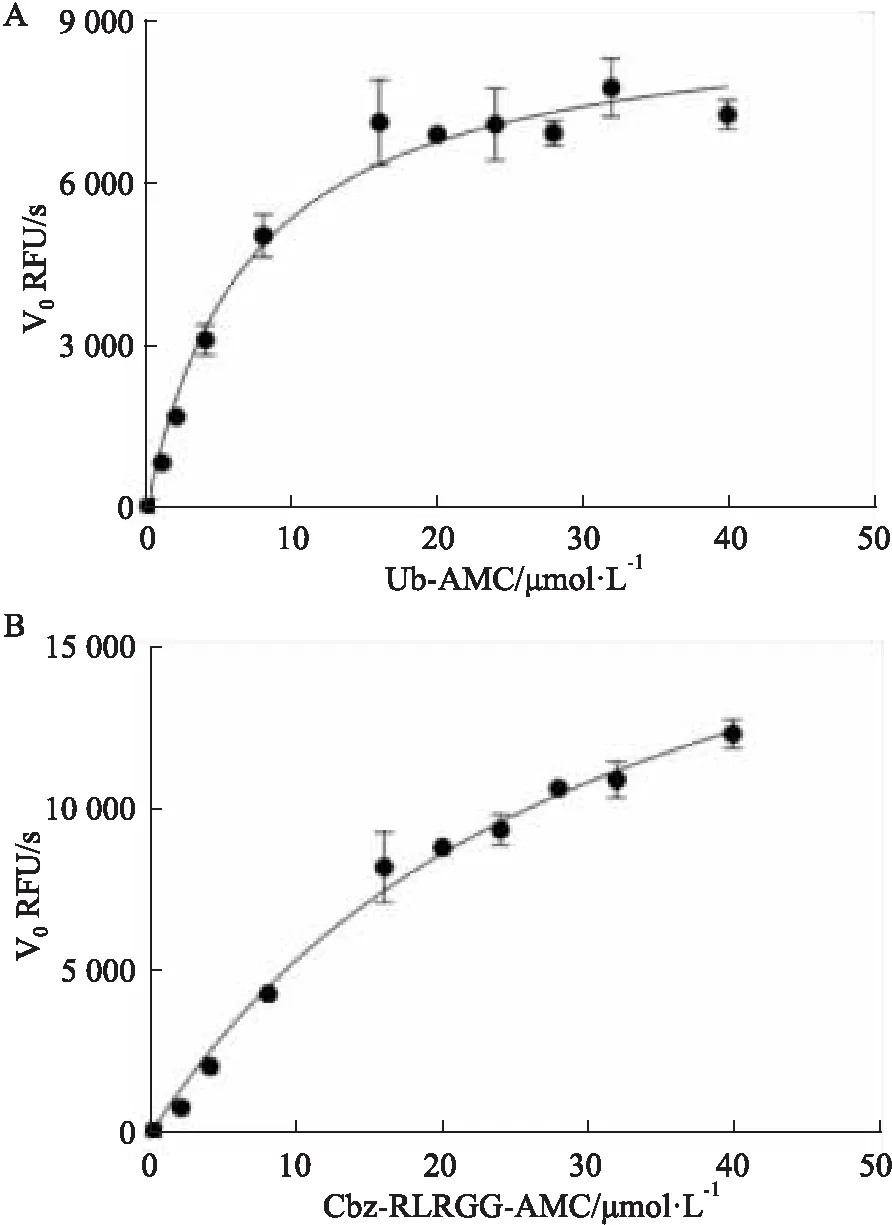
Fig 2 Michaelis constant curve of SARS-CoV-2 PLpro
2.3 葡萄糖酸铜和双硫仑的IC50值以及Ki值测定以葡萄糖酸铜浓度和双硫仑浓度的对数值为横坐标,对应的抑制率的值为纵坐标,用GraphPad Prism 8中的非线性回归(曲线拟合)模块中的四参数法制作曲线,并分析计算出葡萄糖酸铜和双硫仑的IC50值。荧光底物Ub-AMC浓度为2.5 μmol·L-1时,葡萄糖酸铜的IC50为33.02 nmol·L-1(Fig 3A),双硫仑的IC50为480.4 nmol·L-1(Fig 3B),且在相应的浓度范围以内显示出剂量依赖性。已有文献报道,双硫仑对SARS-CoV的PLpro的IC50为6.5 nmol·L-1[8],相比较可以看出,双硫仑对SARS-CoV-2的PLpro蛋白抑制效果更好。根据实验结果可以看出,葡萄糖酸铜和双硫仑都对PLpro蛋白表现出很好的抑制活性,其抑制常数Ki值通过Dixon作图法求得,两条曲线交点横坐标的负值即为Ki值。如Fig 4所示,葡萄糖酸铜的Ki值为55.3 nmol·L-1,双硫仑的Ki值为821.6 nmol·L-1。Dixon作图法得到的Ki值在一定程度可以反映抑制剂的抑制类型,葡萄糖酸铜的两条曲线相交于X轴上方,则为竞争性抑制剂,双硫仑相交于X轴,则为非竞争性抑制剂,为了进一步验证其抑制类型以及两者相互作用的影响,设计了酶动力学实验做更深的探索。
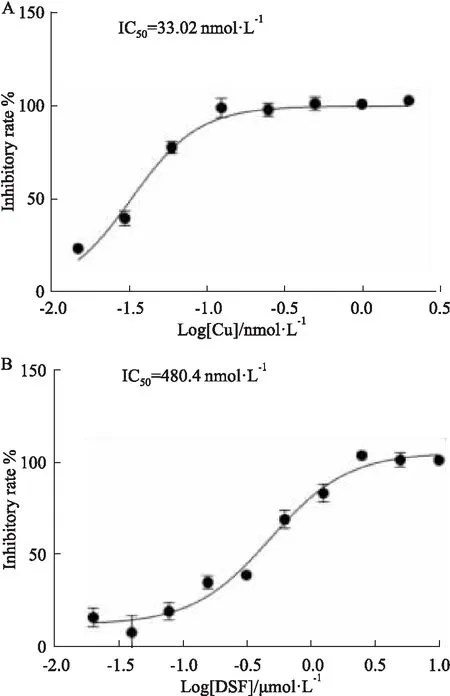
Fig 3 IC50 curves of copper gluconate and disulfiramagainst SARS-CoV-2 PLpro

Fig 4 Dixon Plot for copper gluconate and disulfiram attwo different substrate concentrations
2.4 葡萄糖酸铜和双硫仑的酶动力学测定葡萄糖酸铜和双硫仑都对PLpro蛋白有很好的抑制作用,但是关于两种抑制剂对于PLpro蛋白的抑制类型需要进一步的实验测定。将实验结果用Lineweaver-Burk双倒数作图法判断葡萄糖酸铜和双硫仑的抑制类型,如Fig 5所示,葡萄糖酸铜的4条不同浓度的曲线相交于y轴,具有固定的Vmax值,不同的Km值,表现出的抑制类型为竞争性抑制,而相反,双硫仑的四条不同浓度的曲线相交于X轴,具有不同的Vmax值,相同的Km值,表现的抑制类型为非竞争性抑制。
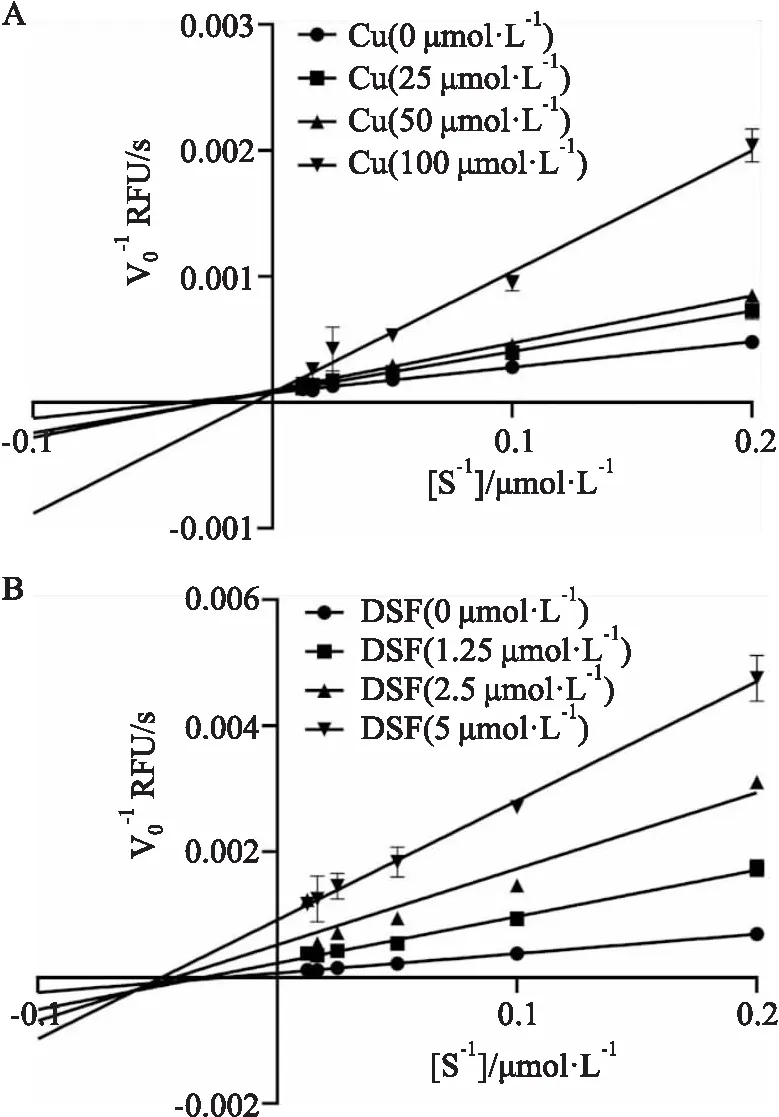
Fig 5 Lineweaver-Burk double reciprocal curveof copper gluconate and disulfiram
为了阐明葡萄糖酸铜和双硫仑与PLpro之间相互作用的动力学机制,采用Yonetani-Therell方法对两种抑制剂相互作用对PLpro蛋白影响做了分析。在不同浓度的葡萄糖酸铜和一定浓度的双硫仑存在下测量PLpro蛋白的水解活性,数据用Dixon图展示。如Fig 6所示,我们发现葡萄糖酸铜和双硫仑对PLpro蛋白表现出协同抑制作用。在葡萄糖酸铜存在和不存在的两种情况下,Yonetani-Theorell图中的两条线在X轴上方相交,α值<1,这表明葡萄糖酸铜和双硫仑对PLpro具有协同抑制效应,一种抑制剂与PLpro蛋白的结合增加另一种抑制剂和PLpro蛋白的亲和力。根据葡萄糖酸铜的抑制类型的分析结果,以及已经有文献证明金属离子可以与半胱氨酸蛋白酶的活性位点结合,葡萄糖酸铜可能与PLpro蛋白活性中心的催化三联体相互作用,如Fig 7A所示,其中催化三联体中的Cys的-SH部分被修饰,导致蛋白的性质发生变化,最终PLpro活性被抑制。双硫仑的抑制类型为非竞争性抑制剂,且可以与葡萄糖酸铜协同抑制PLpro,双硫仑或许可以与PLpro蛋白活性口袋以外的结合位点相互作用,比如锌指结构,如Fig 7B所示,锌离子不稳定,由4个保守的半胱氨酸残基四面体配位,对PLpro蛋白结构的正确折叠起着至关重要的作用。双硫仑可能通过与半胱氨酸的巯基结合破坏了锌指结构域的四面体结构,导致PLpro蛋白的活性发生变化。
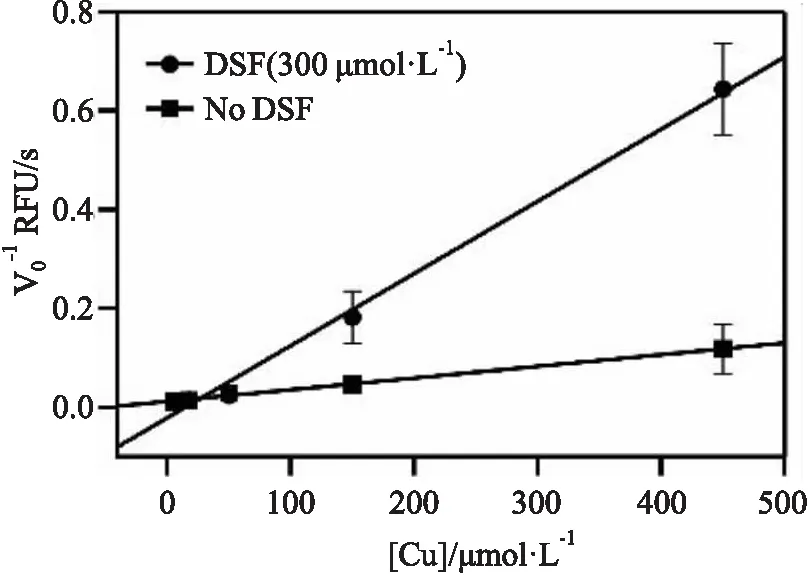
Fig 6 Yonetani-Theorell Plot for copper gluconateand disulfiram(α<1)
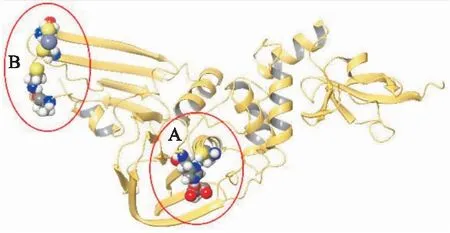
Fig 7 Crystal structure of SARS-CoV-2 PLpro (PDB 7cmd) representing two domains
2.5 双硫仑代谢物对PLpro蛋白影响分析双硫仑的代谢物在低浓度的时候对PLpro抑制效果不佳,浓度达到1 000 μmol·L-1时才能对PLpro蛋白表现出一定的抑制作用(Fig 8)。上述结果表明,与双硫仑代谢产物在抗肿瘤的研究结果不同的是,在对新型冠状病毒的研究中,双硫仑代谢物对PLpro蛋白无明显抑制活性。
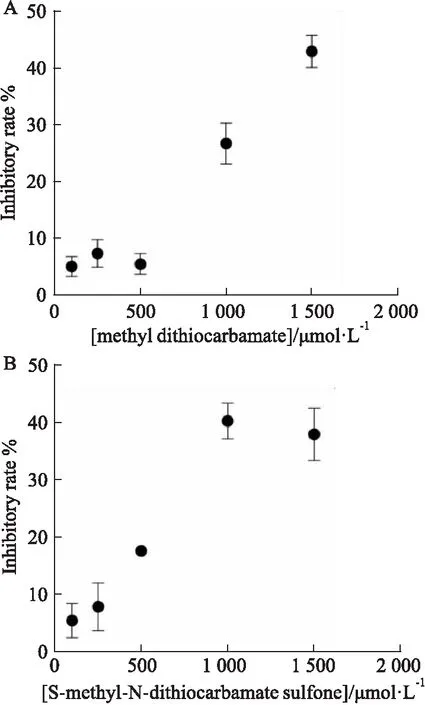
Fig 8 The inhibitory rate of methyl dithiocarbamate andS-methyl-N-diethylthiocarbamate sulfone againstSARS-CoV-2 PLpro
3 讨论
PLpro参与新型冠状病毒的复制、翻译和水解过程,在新型冠状病毒生命周期至关重要,通过研究发现葡萄糖酸铜和双硫仑对新型冠状病毒PLpro具有较强的抑制活性,葡萄糖酸铜可以竞争性抑制PLpro的活性来阻断新型冠状病毒的生命周期,进而成为一种预防或者治疗新型冠状病毒感染潜在药物,葡萄糖酸铜还可以单独作为一种原料药,或与其他已知的药物、化合物或载体相配合制成口服剂、注射剂、涂剂、洗剂、气雾剂、油制剂或透皮贴剂等各种形式的药剂用来预防或者治疗新型冠状病毒感染。双硫仑非竞争性抑制PLpro,是一种已知的戒酒药,临床使用多年,药动学和安全性有保障。葡萄糖酸铜和双硫仑单独使用对新型冠状病毒的PLpro具有较好的抑制效果,联合使用对PLpro的抑制活性增强,具有协同抑制效应,综上所述,葡萄糖酸铜和双硫仑有望成为抗新型冠状病毒的候选药物。
