亚热带天然林转变为毛竹林和茶园对土壤胞外酶活性的影响
2022-05-05张海阔张宝刚周钟昱梁佳辉吴家森吕玉龙解宏图蔡延江
张海阔,张宝刚,周钟昱,梁佳辉,吴家森,吕玉龙,解宏图,蔡延江*
(1.浙江农林大学省部共建亚热带森林培育国家重点实验室,杭州 311300;2.浙江农林大学环境与资源学院,杭州 311300;3.浙江农林大学林业与生物技术学院,杭州 311300;4.安吉县自然资源和规划局,浙江 安吉 313300;5.中国科学院沈阳应用生态研究所,沈阳 110016)
土壤胞外酶是一种来源于土壤微生物、植物根系或死亡动植物残体且具有催化作用的蛋白质,其在土壤中主要以游离态或矿物结合态等形式存在。土壤胞外酶参与土壤物质循环的关键步骤,可以催化有机质的分解和矿化,直至有机质分解产物可以被微生物同化利用。土壤胞外酶活性(Extracellular enzyme activities,EEAs)对环境变化或人类活动所引起的土壤扰动响应灵敏,因此,EEAs常被视为土壤生态系统生物地球化学过程发生改变的敏感指标。土壤中多种胞外酶对物质循环存在功能分化,即多种不同的酶可催化同一种物质或同一种酶可催化土壤中多种物质的分解。在若干种土壤胞外酶中,与碳循环相关的β-葡萄糖苷酶(β-1,4-glucosidase,BG)、纤维二糖水解酶(Cellobiohydrolase,CBH)、β-木糖苷酶(β-1,4-xylosidase,BX)、酚氧化酶(Phenol oxidase,POX)和过氧化物酶(Peroxidase,PER),与氮循环相关的N-乙酰-β-D 氨基葡萄糖苷酶(β-1,4-N-acetylglucosaminnidase,NAG)、亮氨酸氨基肽酶(Leucine aminopeptidase,LAP),以及与磷循环相关的土壤酸性磷酸酶(Acid phosphatase,ACP)得到了广泛研究。
人类活动的加剧正在破坏着越来越多的天然林,在1980 年至1990 年间,人们受经济收益和粮食需求的驱使而大规模砍伐森林,仅全世界热带地区就有83%的新增农业用地是由天然林直接改造而来。将天然林转变为人工林或种植园的现象在我国热带及亚热带地区也频繁发生。由天然林转变而来的人工林或种植园常伴随着施肥、耕作、林下植被去除等集约经营措施,这些管理措施会通过影响pH、含水量、有机质积累、底物有效性和微生物生物量等土壤性质而影响土壤EEAs。例如,在中国亚热带的四川洪雅林场,将天然林转变为杉木人工林后,由于底物有效性的降低,土壤BG 和NAG 的活性均显著降低;在中国亚热带的浙北地区,将天然常绿阔叶林转变为集约经营的毛竹人工林后,土壤ACP、脲酶和蛋白酶的活性受施肥和林下植被去除等影响均显著降低;在印度热带的Hathinala 地区,将天然林转变为木豆种植园等农用地后,由于化肥的施用而降低了土壤ACP 活性。此外,土壤EEAs 还受到多种生物因素和非生物因素的调控,众多研究者的研究结果也不尽相同。因此,深入探究由天然林转变为不同人工林或种植园后土壤EEAs的变异特征及其关键驱动因子,具有十分重要的意义。
综上可知,大量研究均证实天然林转变为农业用地可显著影响土壤的EEAs,由于天然林转变为农业用地后大都采用了不同的经营管理措施,这些管理措施可直接改变土壤环境因子,土壤环境因子的改变又直接关系到土壤EEAs的变化。例如天然林转变为集约经营的毛竹人工林后的经营措施是通过改变土壤活性氮、磷的含量而间接改变土壤氮、磷获取酶的活性;天然林转变为杉木人工林的措施则是通过改变土壤微生物群落结构间接降低土壤碳、氮获取酶的活性。位于中国亚热带地区的浙江省拥有丰富的天然林资源,从20 世纪末至21 世纪初期将天然林转变为人工林或种植园的现象时有发生。天然林转变后的人工林或种植园多采用粗放经营或集约经营等管理措施,这极有可能显著改变土壤的环境因子,进而影响土壤EEAs 的活性。相比天然林,这一地区的人工林或种植园土壤EEAs发生了怎样的变化?起关键驱动作用的土壤环境因子是什么?这些问题还亟待进一步探究。为此,本研究选取浙江省安吉县的天然林土壤及由天然林转变的毛竹林和茶园土壤作为研究对象,分析探讨了土壤EEAs 的变化特征及其与土壤环境因子的关系,以期为进一步明确土壤酶学特征对天然林转变为不同类型人工林或种植园后的响应机制提供理论参考。
1 材料与方法
1.1 研究区概况
研究区域位于浙江省安吉县梅溪镇(30°41'~30°42' N,119°46'~119°47' E),该区域属亚热带海洋性季风气候,四季分明,雨热同期,雨季为每年的5—6月,年平均气温15.1 ℃,年降水量1 286 mm,无霜期228 d,土壤属酸性红壤,原生植被为以苦槠()、木荷()、青冈()等为优势种的天然常绿阔叶林(Evergreen broad-leaved forests,EBFs)。
1.2 土壤样品采集与分析
于2019年4—6月对研究区域进行了多次实地踏勘,在结合当地政府部门提供的相关资料后,选取相同海拔、坡向的EBFs 及其邻近的EBFs 转变约15~20 a 的粗放经营毛竹林(Moso bamboo plantations,MBPs)和转变约20 a 的集约经营(每年10 月平均施用400 kg·hm的复合肥,N∶PO∶KO=15∶15∶15,且茶树修剪枝叶还田)茶园(Tea plantations,TPs)为研究对象,各设置3 块样地,每块样地面积约20 m×20 m。于2019 年7 月在每块样地中根据五点取样法采集0~20 cm 的土壤样品,土样过2 mm 筛后混合成一份样品。将土壤样品置于4 ℃便携式冰箱中迅速带回实验室并分成两份,一份自然风干用于测定土壤基本理化性质,另一份保存在4 ℃冰箱中用于测定土壤生化指标。
土壤容重(BD)采用环刀法测定;pH(水土比2.5∶1)利用 METTLER TOLEDOSeven Compact 型 pH 计(METTLER TOLEDO,USA)测定;土壤有机碳(SOC)采用外加热重铬酸钾氧化-容量法测定;全氮(TN)采用半微量凯氏定氮法测定;全磷(TP)采用高氯酸-硫酸-钼锑抗比色法测定;有效磷(AP)采用盐酸-氟化铵-钼锑抗比色法测定;微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸直接提取法测定。土壤EEAs 采用改进后的微孔板荧光法测定。各种酶的名称、缩写、作用及特定底物见表1。分别以4-甲基伞形酮(4-methylumvelliferyl,4-MUB)和L-二羟苯丙氨酸(Levodopa,L-DOPA)作为测定水解酶(其中LAP 用7-Amino-4-methylcoumarin)和氧化酶活性的标准对照物质。取1.5 g 新鲜土壤放入125 mL醋酸钠缓冲液(pH 5.0)中,以磁力搅拌器匀质1 min 后,用八通道移液器分别 3 次吸取 200 µL 土壤浆液注入96 孔板内作为样品组,样品组的第1~3 列额外再分别加入50µL的醋酸钠缓冲液、标准对照物质和特定底物;此外,再分别3 次吸取200µL 醋酸钠缓冲液注入96 孔板内作为对照组,对照组的第1~3列也额外分别加入50µL 的醋酸钠缓冲液、标准对照物质和特定底物。加液完毕后,将96 孔板经25 ℃恒温培养后用 BiotekSynergy H1 型酶标仪(Biotek,USA)读取荧光值或吸光值,其中水解酶在365 nm 激发光和450 nm 发射光下读取,氧化酶在460 nm 吸收光下读取。
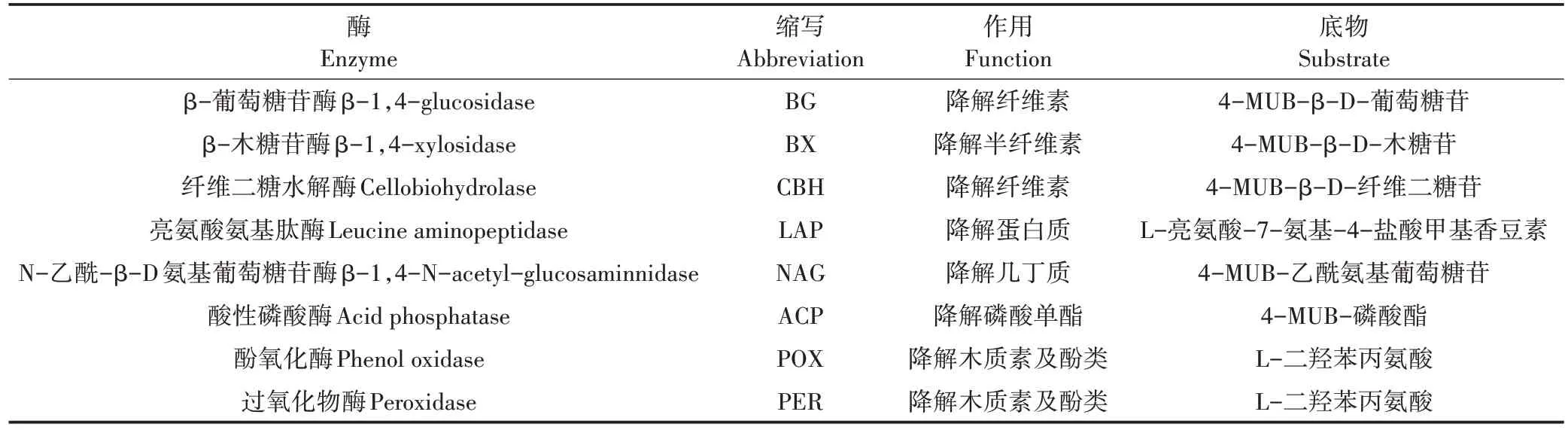
表1 土壤酶的名称、缩写、作用及特定底物Table 1 Names,abbreviations,functions and substrates of the soil extracellular enzymes
1.3 数据处理与分析
试验数据利用Excel 2016 进行预处理,包括计算平均值和标准误差(SE)等,文中所有天然林、茶园和毛竹林数据皆为3 个样地的平均值。在检验数据的正态性和方差齐性后,采用SPSS 18.0 软件的单因素方差分析(One-way ANOVA)及最小显著差异法(LSD)确定3种土地利用类型间测定指标的显著差异性(<0.05 为差异显著),采用Spearman 相关系数对土壤环境因子和土壤EEAs 进行相关性分析。采用R语言 vegan 包、rdacca.hp 包和 ggplot2 包进行冗余分析(RDA)并绘图。
2 结果与分析
2.1 研究区土壤基础理化性质及微生物学性质
天然常绿阔叶林转变为茶园和毛竹林后,土壤BD未发生显著改变(表2)。与天然林土壤相比,毛竹林土壤pH 显著升高(<0.05),而茶园土壤pH 略有降低,但未达到显著水平。天然林转变为毛竹林后SOC含量略有升高但未达到显著水平,而转变为茶园后SOC 含量显著降低了31.6%(<0.05)。土壤TN 和TP含量在3 种土地利用类型间无显著差异。天然林转变为茶园和毛竹林后,土壤C/N 有所下降,但未呈现出显著差异;毛竹林土壤C/P未出现显著变化,而茶园土壤C/P 显著降低了41.2%(<0.05);毛竹林和茶园土壤N/P 略有升高,但均未达到显著水平。天然常绿阔叶林转变为毛竹林后,土壤MBC 和MBN 含量分别显著增加了118.2%和90.6%(<0.05),转变为茶园后,MBC 和MBN 含量却分别显著降低了54.9%和51.2%(图1,<0.05),而MBC/MBN 在3种土地利用类型间无显著差异。
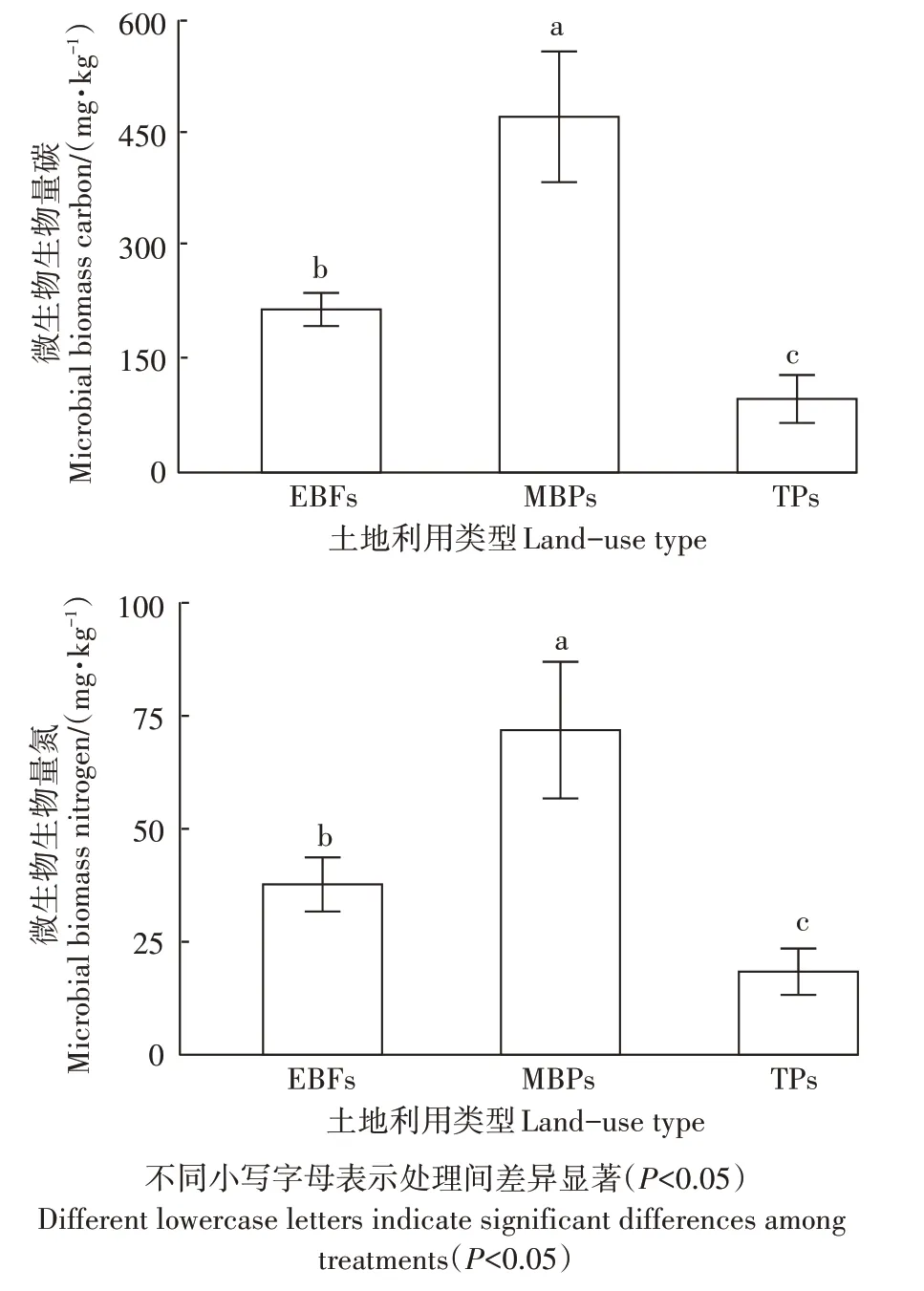
图1 研究区土壤微生物生物量碳、氮含量Figure 1 Soil microbial biomass carbon and nitrogen concentrations in study area
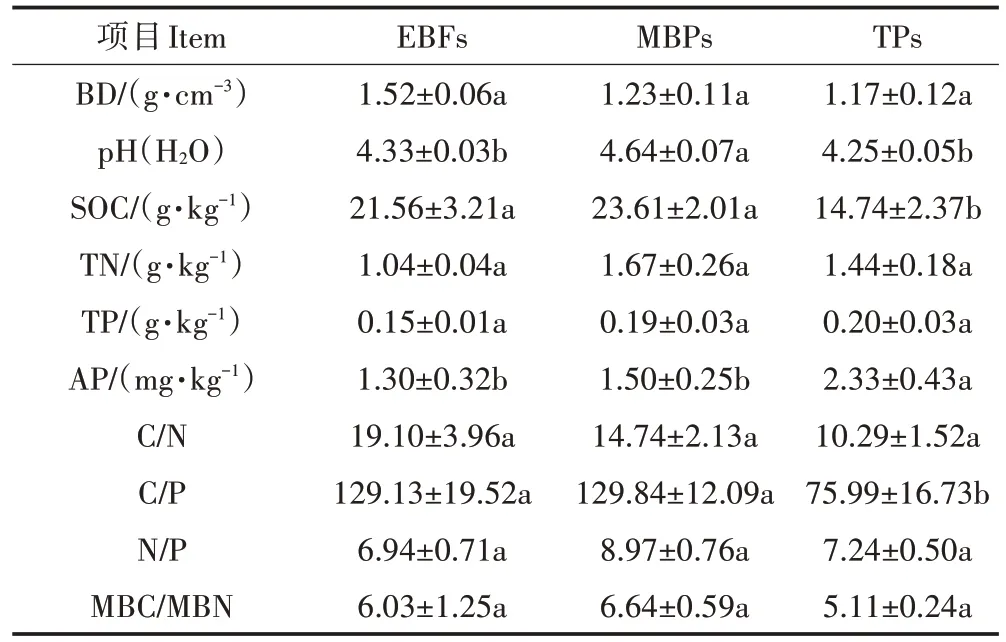
表2 研究区土壤理化性质Table 2 Soil physicochemical properties in study area
2.2 研究区土壤EEAs的特征
天然林和两种人工林的土壤EEAs 结果如表3 所示。天然林转变的毛竹林中,土壤LAP、POX 活性分别显著增加了385%和84.4%(<0.05),土壤ACP 活性显著降低了58.9%(<0.05),其余EEAs 均未发生显著变化;在由天然林转变的茶园中,除土壤BX、LAP 及PER 活性未出现显著变化外,BG、CBH、NAG和ACP的活性均显著降低(<0.05),相比天然林分别降低了29.9%、51.8%、63.9%和36.6%,而POX 活性显著升高了82.9%(<0.05)。
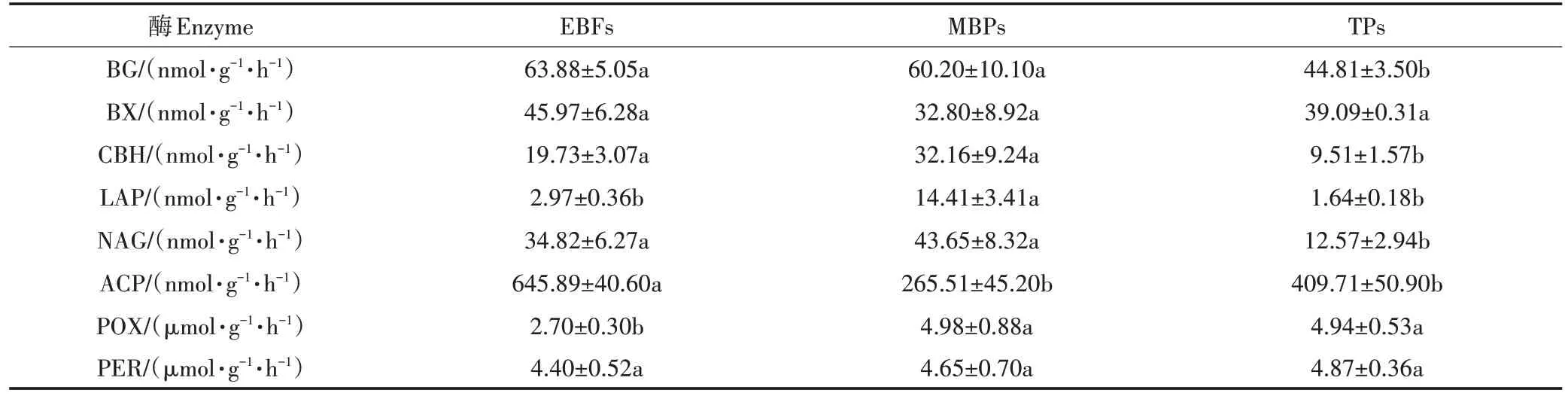
表3 不同土地利用类型土壤胞外酶活性(均值±标准误)Table 3 Soil extracellular enzymes activity in three land-use types(Mean±SE)
2.3 土壤环境因子与土壤EEAs的关系
Spearman 相关性分析结果表明(表4),土壤pH与LAP活性呈极显著正相关(<0.01);土壤SOC含量与BG、CBH、NAG 和LAP 活性均呈极显著正相关(<0.01);土壤 AP 含量与 LAP 活性、C/N 与 POX 活性均呈显著负相关(<0.05);土壤 C/P 与 CBH 和 NAG 活性呈显著正相关(<0.05),与LAP活性呈极显著正相关(<0.01);土壤N/P 与LAP 活性呈显著正相关(<0.05),但与 PER 活性呈显著负相关(<0.05);土壤MBC 和 MBN 均与 BG、CBH、NAG 和 LAP 活性呈极显著正相关(<0.01)。
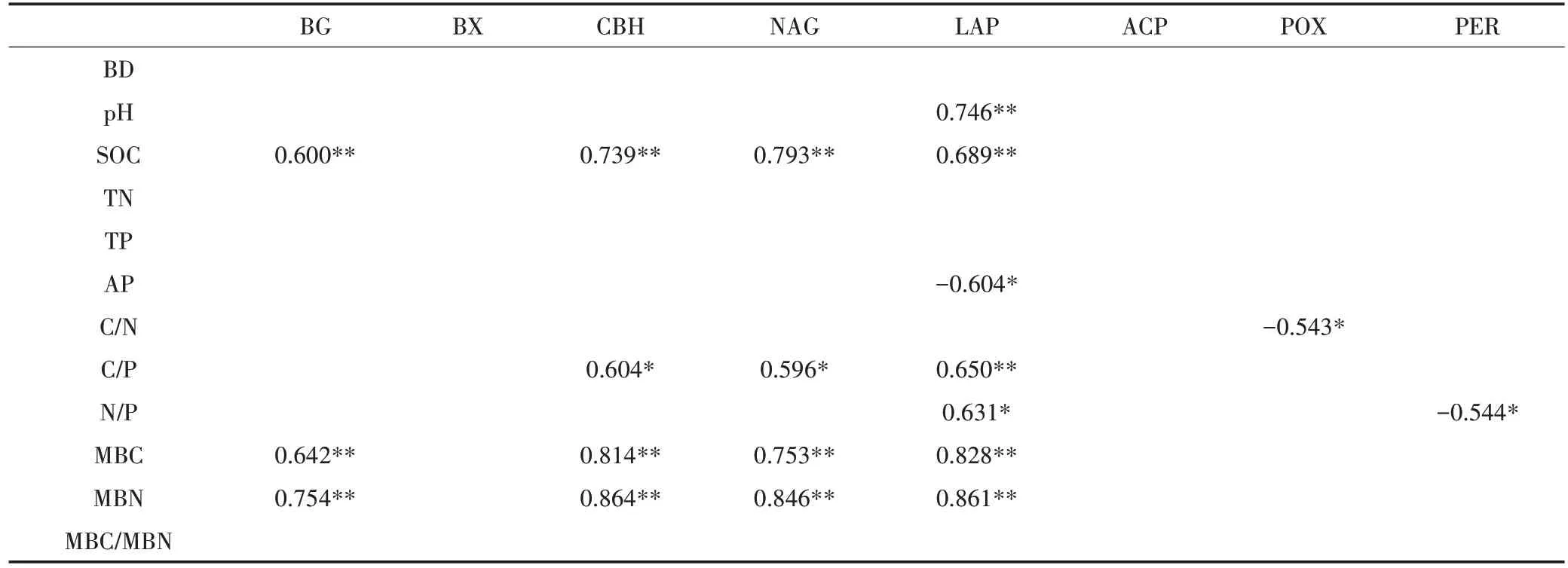
表4 土壤胞外酶活性和土壤环境因子的关系Table 4 Correlations between soil extracellular enzyme activities and the environment factors
为了进一步筛选出影响土壤总体EEAs的关键环境因子,将土壤环境因子BD、pH、SOC、TN、TP、AP、C/N、C/P、N/P、MBC、MBN 和 MBC/MBN 作为解释变量,将土壤EEAs 作为响应变量进行冗余分析。结果发现,RDA 的前两轴分别解释了土壤EEAs 变化的88.03%(RDA1)和6.53%(RDA2),其中土壤pH 是影响土壤总体EEAs的关键驱动因子(图2)。
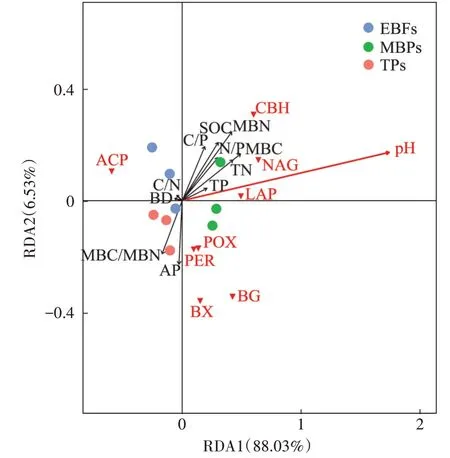
图2 环境因子与土壤EEAs的冗余分析(RDA)Figure 2 Redundancy analysis(RDA)of relationship between soil extracellular enzyme activities with soil environment factors
3 讨论
3.1 土壤EEAs的变化特征
土壤碳转化过程中有机质的降解主要由土壤生物分泌的胞外酶催化实现。本研究发现天然林转变为毛竹林和茶园后,土壤活性碳及惰性碳获取酶的活性分别产生了不同的响应。天然林转变为粗放经营的毛竹林后,除土壤POX 活性显著增加外,土壤活性碳获取酶BG、BX、CBH 以及惰性碳获取酶PER 的活性均未发生显著变化,这与LIN 等在相邻区域发现天然林转变为集约经营毛竹人工林后土壤BG、CBH活性均显著降低的结果不同。造成这种差异的可能原因是集约经营毛竹林氮肥的施用会抑制土壤微生物活性,而本研究中粗放经营毛竹林土壤EEAs 并不会受到施肥的抑制。与毛竹人工林不同,天然林转变为集约经营的茶园后,伴随SOC 和MBC 含量的降低,土壤BG 和CBH 活性均显著下降,这可能是因为茶园长期施肥刺激了有机碳的矿化分解,进而使活性有机碳含量降低,由于底物量的减少,微生物分泌活性碳获取酶的量也随之减少,此外,茶园土壤生态系统特殊的物质循环模式造成了土壤酸化和铝离子积累,这也会使土壤微生物丰度和活性下降,从而导致EEAs 的降低。土壤POX 与木质素降解有关,其能将土壤腐殖质中的芳香族化合物氧化成醌,再与土壤中的蛋白质、氨基酸、糖类等反应生成有机质。在本研究中,天然林转变为毛竹林和茶园后,土壤POX 活性均显著升高,这可能是由于天然林与毛竹林、茶园相比拥有更多的凋落物,充足的养分供应使微生物为了降低制造酶的成本而减少了相关酶的生产,但在凋落物较少的毛竹林和茶园土壤中,微生物为了保障养分的供给不得不生产更多的POX 以从顽固性碳库中获取能量。
土壤氮素作为土壤肥力中比较活跃的元素,在生物地球化学循环中具有举足轻重的地位,植物在生长过程中需从土壤获取大量的氮素养分,而这些氮素营养的获取也与土壤的酶促作用有关。本研究中,天然林转变的粗放经营毛竹人工林相比天然林土壤,LAP活性显著增加,NAG 活性略有增加但未达到显著水平;而天然林转变的集约经营茶园相比天然林,土壤NAG 活性和LAP 活性分别降低了63.9%和44.8%,根据资源配置理论可解释为茶园土壤施肥增加了氮素等养分的输入,充足的氮素使微生物或植物根系等不需要分泌更多的氮素获取酶,因此降低了茶园土壤中NAG 和LAP 的活性。此外,氮获取酶活性的降低也可能预示着茶园土壤微生物依靠有机氮矿化来获取氮素养分的活动减少,氮肥成为茶园土壤微生物的主要氮素养分来源。
土壤ACP是主要的磷素获取酶,在本研究涉及到的6 种水解酶中,其活性最高,这在一定程度上反映出研究区域可能存在较强的磷限制。天然林转变为毛竹林和茶园后土壤ACP 的活性都出现了显著降低,这可能与亚热带森林酸性土壤磷固定效应强,存在磷限制有关。天然林土壤微生物在磷素供给不充足的情况下会增加土壤磷获取酶的生产,以满足其对磷素养分的需求,而施肥使茶园土壤磷素供给充足(茶园土壤AP 含量显著高于天然林土壤),土壤微生物无需分泌更多的磷获取酶,从而造成了与天然林相比,毛竹林和茶园土壤ACP 活性显著降低,且土壤有机磷的矿化过程可能也不活跃。
3.2 土壤EEAs变化的关键驱动因子分析
土壤EEAs 特征受环境因子的驱动。BG 和CBH调节土壤纤维素的降解,尤其是BG 参与纤维素解聚的最后一步,其被认为是最重要的碳获取酶。本研究中土壤SOC 与BG 活性显著相关,这与前人研究结果一致,同时也说明了研究区内纤维素在土壤有机质输入中有较大的贡献。NAG 与LAP 是重要的氮获取酶,其分别影响甲壳素和蛋白质的降解,土壤NAG 活性会因为SOC 含量及土壤微生物量的降低而受到抑制,在本研究中,NAG 与 SOC、MBC 和 MBN有显著的相关性;此外,土壤LAP 活性与pH、SOC、AP、C/P、N/P、MBC 和 MBN 均具有显著的相关性,这说明LAP 活性相较其他土壤胞外酶更容易受到环境变化的影响。土壤C/N 与EEAs 关系密切,土壤EEAs受制于养分有效性及其化学计量比,本研究发现土壤C/N是POX活性的主要驱动因子(表4),且ZHANG等的研究也进一步支持了这一结果,即较低的C/N会使土壤氧化酶的活性增加。
本研究还通过将所有土壤环境因子作为解释变量,所有EEAs 作为响应变量进行RDA,以筛选出土地利用变化后造成土壤总体EEAs变异的关键环境驱动因子。RDA 结果表明(图2),土壤pH 是造成总体EEAs 变异的唯一显著影响因子。值得注意的是,与RDA 结果不同,Spearman 相关性分析显示pH 仅与LAP 活性显著相关,这是由于RDA 与经典相关性分析的原理和目的均不相同,从RDA 可得知土地利用变化后土壤pH 对整体EEAs 变异的解释度,而从Spearman 相关性分析得到的却是pH 与单个土壤EEAs 的相关性,所以两种分析可能会出现不同的结果。此外,以往研究也发现,土壤pH 是影响EEAs 的重要因素,例如ZHANG 等采用RDA 发现中国亚热带地区林地转变为农田后,土壤pH 显著解释了6 种碳获取酶的总变异;TURNER等采集了29个英国低地牧场土壤样品,发现土壤pH 可能通过影响土壤中占优势的生物类群来对磷酸酶活性造成影响。土壤pH 与胞外酶在土壤溶液中的稳定性紧密相关,其可通过改变土壤微生物群落结构和酶的空间结构而改变土壤胞外酶的催化活性。土壤pH 对EEAs 的影响机制较为复杂,本研究中土壤EEAs总变异对pH的响应机制还需在未来采用扩增子测序、功能基因定量或代谢组学等方法做更深入地探究。
4 结论
(1)相比天然林,粗放经营的毛竹人工林土壤活性碳获取酶、NAG 和PER 活性均无显著差异;此外,毛竹林土壤的LAP和POX活性显著高于天然林,ACP活性显著低于天然林。
(2)相比天然林,集约经营茶园土壤BX、LAP 和PER 活性均无显著差异;此外,茶园土壤的POX 活性显著高于天然林,BG、CBH、NAG 和ACP 活性显著低于天然林。
(3)冗余分析发现,粗放经营毛竹林、集约经营茶园和天然林间土壤总体EEAs 存在明显差异,在影响土壤总体EEAs的一系列环境因子中,土壤pH的贡献最大。因此,在土地利用变化背景下,土壤pH可在一定程度上解释亚热带地区由天然林转变的粗放经营毛竹人工林和集约经营茶园中土壤总体EEAs的变异规律。
