核桃/大豆间作对氮素吸收利用及转移的影响
2022-04-12刘小勇韩富军
任 静,刘小勇,刘 芬,彭 海,韩富军
(甘肃省农业科学院 林果花卉研究所,甘肃 兰州 730070)
氮(N)素是生命体的必需元素,是陆地生态系统植物碳同化的主要限制营养元素[1]。在许多农作物系统中,土壤中有限的自然N 素供应限制了作物的生长与产量[2]。为确保产量最大化,作物生产中普遍存在过高的化学N 肥投入和过低的生物多样性等情况,导致了能源成本升高、生态环境破坏及人类健康受损等。因此,在兼顾生产效益和生态建设的目标下,减少N 肥使用量并提高N 素利用效率对现代可持续农业的发展至关重要[3-4]。
间作复合系统是典型的资源高效型种植模式,不仅能通过物种营养生态位与空间生态位的互补来有效利用资源,又能增加物种多样性,改善土壤质量,进而实现高产和优产的目的[5-7]。由于豆科作物具有生物固氮功能,可有效补充土壤中的N 素,其与非豆科作物间作时优势十分突出。有研究结果表明,豆科间作系统中间作群体产量高于N 肥低投入制度下单一作物产量[8]。紫花苜蓿与核桃幼树间作在改善核桃N 素状况的同时,提高了幼树生长量[9]。更为重要的是,间作物种对土壤N素的竞争刺激豆科作物更加依赖共生固氮,从而拓宽N 素营养生态位,进一步提高间作系统的N 素利用效率[10-11]。例如:与豆科间作时,作物群体吸N 量明显高于相对应单作吸N 量的加权平均值[12-13];南酸枣/花生间作系统中,5年生南酸枣竞争利用了9.66%的施于花生的N 肥,促进了花生生物固氮[14];燕麦/花生间作系统提高了成熟期花生的固氮效率,燕麦地上部N 素积累量比单作增加了49%,且间作花生中N 素向燕麦的转移率达21.4%[15]。土壤N 素和豆科作物固定的大气中的N 素是低投入豆科和非豆科间作最主要的N 素来源,豆科作物在参与间套作时均表现出较强的N 素优势。
核桃Juglans regia是重要的干果树种和珍贵用材树种,兼具经济价值和生态价值[16]。在甘肃省,核桃树大多种植于以低氮状态为代表的土壤上,人为氮输入量较高,加之幼苗定植行距宽,导致土地利用效率低和环境成本高等问题的出现。在甘肃省,幼龄核桃园间作大豆是一种处于优势地位的栽培模式,但有关其间作系统对豆科生物固氮养分利用及间作系统内N 素运移情况等的研究鲜见报道。本研究中利用15N-尿素示踪技术,通过种植袋间作试验,探究不施N 肥条件下,核桃幼树对N 素的吸收、利用和间作模式中N 素运转能力,旨在为阐明大豆生物固氮对核桃N 营养的贡献及建立核桃间作大豆模式提供参考。
1 材料与方法
1.1 试验地概况
2020年4—10月,在甘肃省农业科学院主要果树种质资源圃(103°41′E,36°6′N)进行试验。试验地属温带大陆性气候,海拔1 530 m,年均降水量329 mm,年均气温9.6 ℃,极端最低气温-25 ℃,无霜期196 d。供试土壤为黄绵土,前茬为3年生蒲公英自然生草果园空地,0 ~20 cm 的耕层土壤中有机质、全氮、全磷、全钾的质量分数分别为22.3、1.45、1.18、22.06 g/kg,其速效氮、有效磷、速效钾的质量分数分别为117.71、28.90、334.30 mg/kg,pH 为8.37。将土壤样品除去杂质后备用。
1.2 试验材料
试验用1年生‘元林’核桃嫁接苗为2019年3月25日定植,其砧木为甘肃陇南成县普通晚实厚壳核桃,选择树体生长均匀一致的核桃嫁接苗进行试验,其主干高度为60 cm,冠幅80 cm。大豆品种为2020年4月15日播种的‘陇黄3号’。核桃嫁接苗和大豆均为当地主栽品种。
采用种植袋进行种植,种植袋长宽均为140 cm,每袋装入园土的高度为60 cm,预留20 cm便于浇水。15N-尿素为上海化工研究院生产,15N的丰度为10.10%。
1.3 试验设计
试验中设置3 种种植模式,分别为核桃/大豆间作、核桃单作和大豆单作,具体种植方式如图1所示,每个种植袋设为1个小区,种植面积1.96 m2。核桃/大豆间作模式为距离树干50 cm 的东西两侧,自北向南分别种植1 行大豆,大豆株距20 cm,每袋10 株,核桃投影面积占地42%,大豆投影面积占地58%[17]。核桃单作和间作模式中,核桃嫁接苗均植于种植袋中心。大豆单作模式中,将核桃树替换为1 排自北向南种植的大豆,株行距为20 cm×50 cm,每袋3 行,共15 株。每处理9 次重复,完全随机排列。
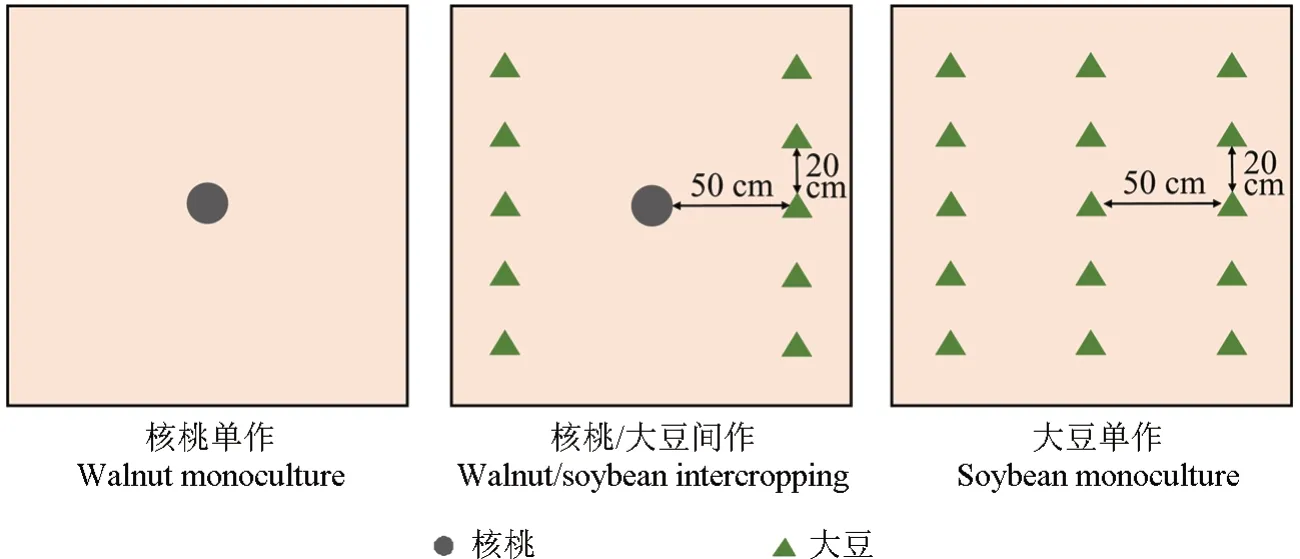
图1 核桃和大豆种植模式单小区示意Fig.1 Single plot diagram of walnut and soybean planting patterns
1.4 试验方法
1.4.1 施 肥
2020年5月10日,开始15N 同位素标记试验,3种种植模式的施肥量相同,为P2O513.2 g、K2O 7.8 g和15N-尿素溶液(0.4 g/L)5 L,均为田间常规用量,同时对3 种种植模式设置不标记试验。在核桃单作模式中,挖开核桃树盘表层土壤(半径35 cm、深度10 cm)进行施肥,然后立即回填表层土壤;在大豆单作模式中,采用条施的方式施肥,即在大豆各行同侧开挖条状沟(宽度5 cm、深度5 cm),均匀施入肥料后,将挖出的表层土壤回填;在核桃/大豆间作模式中,核桃树施肥方法同其单作模式,大豆施肥方法同其单作模式,但条状沟开在与核桃相邻一侧。各处理施入15N-尿素溶液的面积相同,且均不再施入额外的N 肥。
1.4.2 取样和测定
分别在果实硬核期(2020年6月25日)、油脂转化期(2020年8月15日)和核桃落叶前(2020年10月5日),对整株进行破坏性取样。将核桃苗分为主干、枝、叶和根4个部分,将大豆分为豆荚、秸秆和根3个部分。将各部分带回实验室,称取鲜质量,然后于105 ℃条件下杀青30 min,于80 ℃条件下烘至恒质量,冷却后称量各部分的干质量。取出部分干样进行粉碎,过0.25 mm 筛,混匀后装袋备用。所有植株样品全氮含量的测定均采用凯氏定氮法,15N 丰度使用IsoPrime100 稳定同位素质谱仪(英国IsoPrime 公司)进行测定。
1.5 数据计算和分析
1.5.1 植株样品中N 素来自肥料的比例[17]
R1=(Ps1-0.366 3%)×100%/(Pf-0.366 3%);
R2=(Ps2-0.366 3%)×100%/(Pf-0.366 3%)。
式中:R1为核桃各器官样品中N 素来自肥料的比例;Ps1为核桃各器官样品中15N 的丰度;Pf为肥料中15N 的丰度;0.366 3%为N 素的自然丰度;R2为大豆各器官样品中N 素来自肥料的比例;Ps2为大豆各器官样品中15N 的丰度。
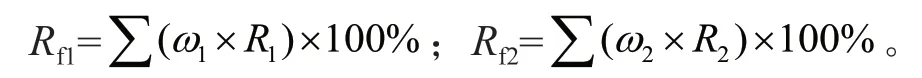
式中:Rf1为核桃植株样品中N 素来自肥料的比例;ω1为核桃植株各器官干质量占全株总干质量的比例;Rf2为大豆植株样品中N 素来自肥料的比例;ω2为大豆植株各器官干质量占全株总干质量的比例。
Rf′=(Rf1×x+Rf2×y)×100%。
式中:Rf′为单作或间作体系样品中N 素来自肥料比例的加权平均值;在单作体系中,x=42%,y=58%;在间作体系中,x=1,y=1。
1.5.2 植株N 素的分配率和利用率[17]
m1=md1×RN1×R1×1 000;
m2=md2×RN2×R2×1 000。
式中:m1为核桃各器官15N 的吸收量,mg;md1为核桃各器官总干质量,g;RN1为核桃各器官N 含量占全株N 含量的比例;m2为大豆各器官15N 的吸收量,mg;md2为大豆各器官总干质量,g;RN2为大豆各器官N 含量占全株N 含量的比例。
Rd1=(m1/ms1)×100%;Rd2=(m2/ms2)×100%。
式中:Rd1为核桃各器官15N 的分配率;ms1为核桃植株对15N 的吸收总量,mg;Rd2为大豆各器官15N的分配率;ms2为大豆植株对15N 的吸收总量,mg。

式中:Ru1为核桃植株样品中N 素的利用率;ma为15N 的施用量,g;Ru2为大豆植株样品中N 素的利用率。
Ru′=(Ru1×x+Ru2×y)×100%。
式中:Ru′为单作或间作体系样品中N 素利用率的加权平均值;在单作体系中,x=42%,y=58%;在间作体系中,x=1,y=1。
1.5.3 大豆固氮效率[18]
Pe=Ps-PN。
式中:Pe为植株15N 的超量原子百分数;Ps为标记样品中15N 的丰度;PN为未标记样品中15N 的丰度。
E=(1-Pe2/Pe1)×100%。
式中:E为单作或间作体系中大豆的固氮效率;Pe1为单作或间作核桃样品中15N 的超量原子百分数;Pe2为单作或间作大豆样品中15N 的超量原子百分数。
1.5.4 氮转移率和核桃中氮素来自大豆固氮转移的比例
Rt=Pe1×mN1×100%/(Pe2×mN2+Pe1×mN1)。
式中:Rt为氮转移率;mN1为核桃样品中N 素的吸收量,mg;mN2为大豆样品中N 素的吸收量,mg。
Pt=(1-Rf1 间/Rf1 单)100%。
式中:Pt为核桃中氮素来自大豆固氮转移的比例;Rf1 间为间作核桃样品中N 素来自肥料的比例;Rf1 单为单作核桃样品中N 素来自肥料的比例。
1.5.5 数据分析
使用SPSS 22.0 软件进行统计和方差分析(ANOVA),使用Microsoft Excel 2016 软件进行图表绘制。
2 结果与分析
2.1 间作对植株干物质量的影响
单作和间作体系中不同生育期核桃和大豆植株的干物质量如图2所示。由图2 可见,不同种植模式下,核桃和大豆的地上部、地下部干物质量在整个生育期的变化规律一致,均呈上升趋势,10月达到最大值,且单作和间作核桃地上部、地下部干物质量均于8月后迅速增加。与6月相比,10月单作和间作核桃地上部干物质量每株分别增加了283.35%、282.16%,地下部干物质量每株分别增加了169.43%、157.63%;与6月相比,10月间作和单作大豆地上部和地下部干物质量每株增长量分别为91.21%、123.00%、71.85%和11.56%。在各生育期,间作与单作模式间,核桃的地上部、地下部干物质量的差异均不显著(P>0.05),大豆的地上部、地下部干物质量的差异均显著(P<0.05),且单作模式下的干物质量高于间作模式。
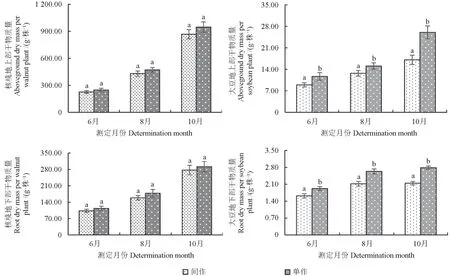
图2 单作和间作体系中核桃和大豆植株的干物质量Fig.2 Dry matter mass of walnut and soybean plants in monoculture and intercropping systems
在间作模式中,除10月间作总干物质量与单作核桃差异不显著(P>0.05),间作总干物质量均显著高于单作(表1)(P<0.05)。因而可知,间作模式总干物质量较单作具有明显的总体优势。

表1 单作或间作体系中核桃和大豆的总干物质量†Table 1 Total dry matter mass of plants at different growth stages g/袋
2.2 间作对植株中N 素来自肥料比例的影响
不同种植模式下核桃和大豆中N 素来自肥料的比例见表2。由表2 可知,生育期(即测定月份)极显著影响核桃各器官中N 素来自肥料的比例(P<0.01);种植模式和生育期对核桃中N 素来自肥料的比例均表现出极显著影响(P<0.01),并随着生育期推进,间作和单作模式下核桃中N 素来自肥料的比例逐渐降低,6—10月分别减少了0.57%和0.58%,且单作模式的各生育期核桃中N 素来自肥料的比例显著高于间作模式(P<0.05)。除种植模式对大豆秸秆中N 素来自肥料的比例有显著影响外(P<0.05),种植模式、生育期及其交互效应均对大豆各器官和整株中N素来自肥料的比例存在极显著影响(P<0.01),且整个生育期间作模式的大豆中N 素来自肥料的比例显著高于单作模式(P<0.05)。种植模式、生育期及其交互效应对间作或单作体系中N素来自肥料比例的加权平均值产生了极显著影响(P<0.01);6—10月,间作和单作模式下大豆中N 素来自肥料的比例均大于核桃。同时,间作体系中N 素来自肥料比例的加权平均值显著大于单作体系(P<0.05)。
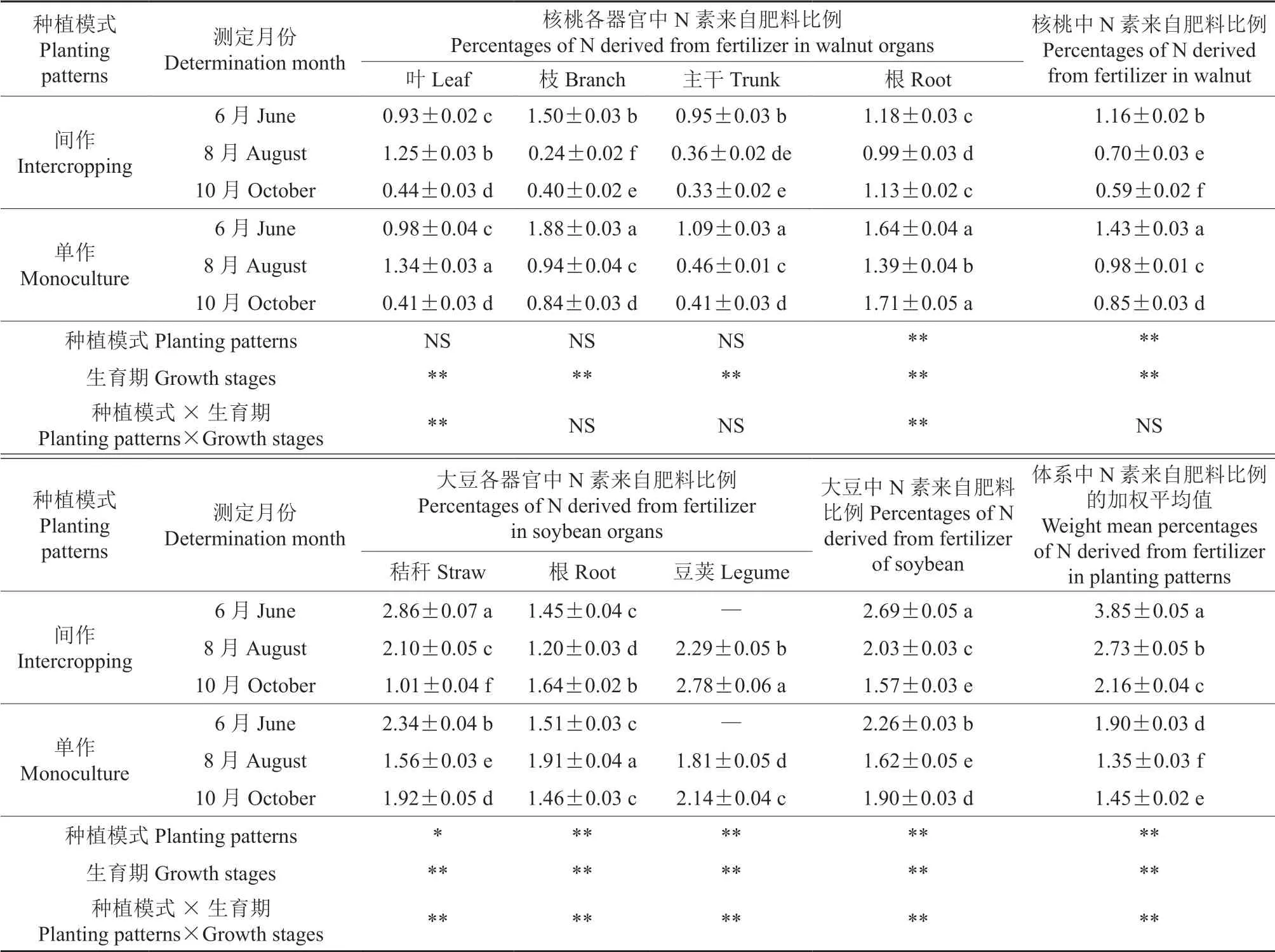
表2 不同种植模式下核桃和大豆中N 素来自肥料比例†Table 2 Percentages of N derived from fertilizer in walnuts and soybeans under different planting patterns %
在整个生育期内,种植模式对核桃和大豆各器官中N素来自肥料比例的变化趋势的影响较大。2 种种植模式下,核桃叶、主干和根中N 素来自肥料比例的变化趋势基本相同,分别为先升后降、不断降低及先降后升3 种趋势;2 种种植模式下,核桃枝中N 素来自肥料比例的变化趋势不同,间作模式下8月出现谷值,单作模式下则持续降低。相同生育期中,除10月核桃叶外,单作模式下核桃各器官中N 素来自肥料的比例均显著高于间作模式(P<0.05)。间作和单作模式下大豆豆荚中N 素来自肥料的比例不断上升,采收时较8月分别增长了21.40%和18.23%;间作模式下大豆秸秆和根中N 素来自肥料比例的变化趋势与单作模式下的变化趋势均不同。
2.3 间作对植株各器官15N 分配率及N 肥利用率的影响
不同种植模式下核桃和大豆各器官的15N 分配率和利用率见表3。由表3 可知,种植模式、生育期及其交互效应显著或极显著地影响了核桃和大豆的各器官15N 分配率(P<0.05 或P<0.01)。随着生育期推进,间作和单作模式下核桃叶、枝的15N 分配率均于8月出现峰值,间作模式下核桃叶15N 分配率的峰值较单作模式下显著提高了17.75%(P<0.05),枝15N 分配率的峰值较单作模式下显著降低了8.41%(P<0.05);核桃根的15N 分配率恰恰相反,8月出现全年最低值,与单作相比,间作模式下核桃根的15N 分配率显著提高了5.24%。间作模式下核桃主干15N 分配率的变化趋势与根相同,单作模式下核桃主干15N 分配率呈持续下降趋势。6月和8月核桃各器官的15N 分配率均表现为地上部(叶、枝、主干)大于地下部(根),10月则相反。与核桃相比,间作和单作模式下大豆各器官的15N 分配率表现出相同的变化趋势;10月,间作和单作模式下秸秆的15N 分配率达到最小值,分别较6月降低了84.46%和67.20%;10月,间作和单作模式下豆荚的15N 分配率达到最大值,较8月分别增加了68.47%和56.59%,且间作模式下豆荚的15N分配率显著高于单作模式(P<0.05)。
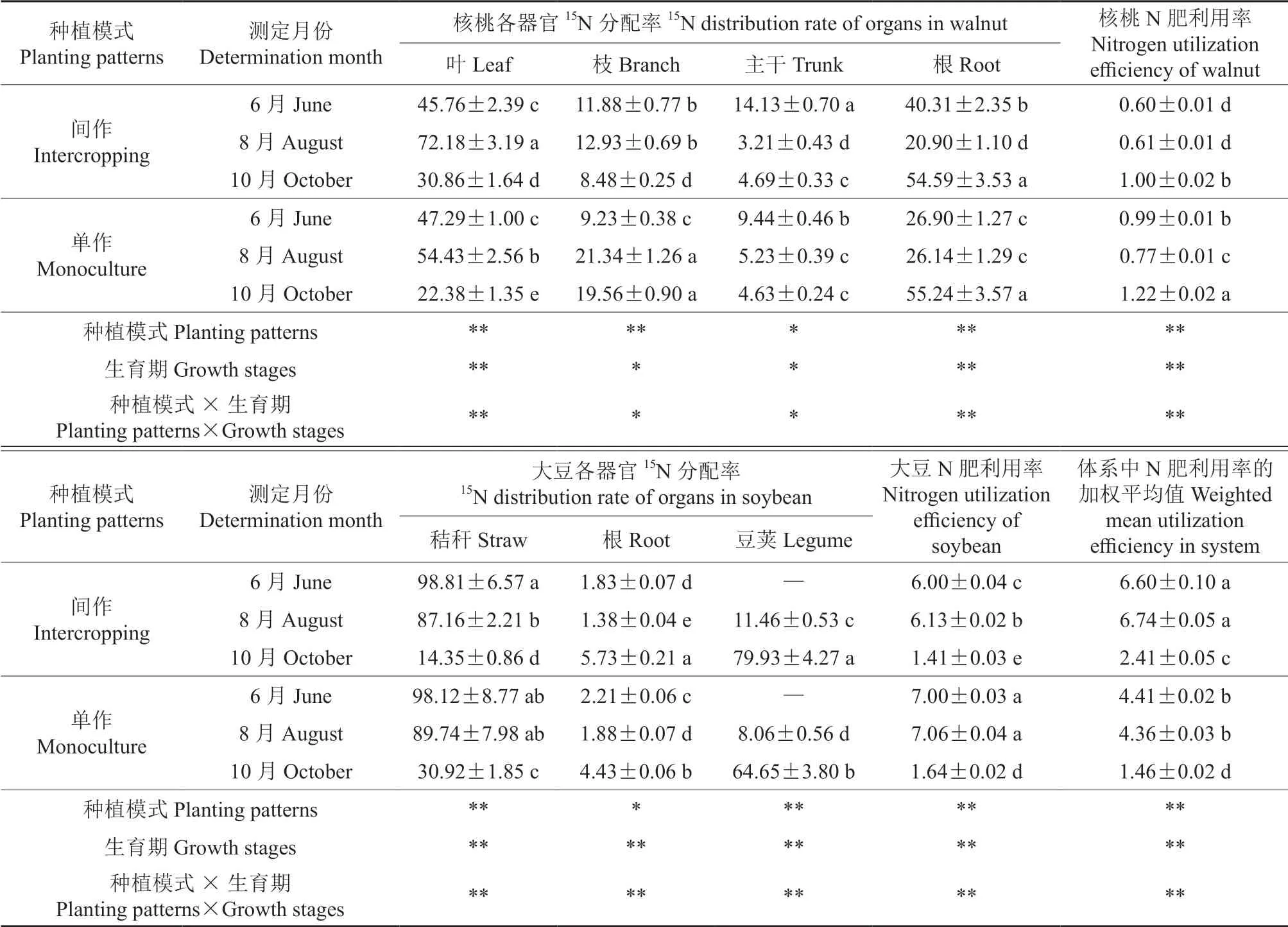
表3 不同种植模式下核桃和大豆各器官的15N 分配率和利用率†Table 3 The value of 15N distribution rate and nitrogen use efficiency of walnuts and soybeans under different planting patterns %
种植模式、生育期及其交互效应极显著影响了核桃N 肥利用率、大豆N 肥利用率、间作和单作体系N 肥利用率的加权平均值(P<0.01)。10月,间作和单作模式下核桃N 肥利用率均达到最大值,与单作模式相比,间作模式下核桃N 肥利用率显著降低了0.22%(P<0.05),大豆N 肥利用率、间作和单作体系N 肥利用率的加权平均值此时均为最低。在整个生育期内,单作模式下核桃和大豆N 肥利用率大于间作模式,大豆的N肥利用率大于核桃,间作体系的N 肥利用率加权平均值显著高于单作体系(P<0.05)。
2.4 间作对大豆固氮效率及氮由大豆向核桃转移的影响
不同种植模式下大豆的固氮效率如图3所示。由图3 可见,在整个生育期,不同种植模式下大豆生物固氮效率的变化趋势基本一致,固氮效率均不断升高。6月和8月,种植模式对大豆的固氮效率均有显著影响(P<0.05)。6月,单作模式下大豆的固氮效率高于间作;8月,间作模式下大豆的固氮效率高于单作。10月,大豆的固氮效率达到最大值,间作模式下为63.74%,单作模式下为61.26%,但差异不显著(P>0.05);10月,间作和单作模式下大豆的固氮效率分别较6月增加了36.31%和26.11%。
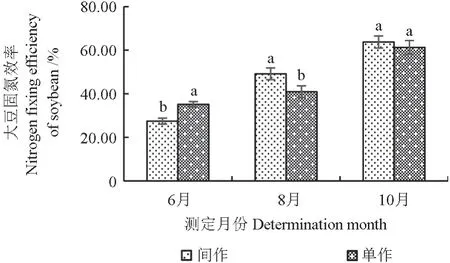
图3 不同种植模式下大豆的固氮效率Fig.3 Biological nitrogen fixation efficiency of soybeans in different planting patterns
间作体系中核桃和大豆N 素含量见表4。由表4 可知,在整个生育期,间作模式下大豆N 素含量均高于核桃,且差异显著(P<0.05),大豆N 素含量为0.16 ~0.33 g/株,核桃N 素含量为1.26 ~1.81 g/株。结果表明,核桃与大豆的N 素含量间存在浓度差,大豆固定的N 素可向间作核桃转移[19]。

表4 间作体系中核桃和大豆N 素含量Table 4 Nitrogen content of walnuts and soybeans in intercropping system g/株
核桃中N 素来自大豆固氮转移的比例如表5所示。由表5 可知,随生育期的推进,N 素转移率和核桃中N 素来自大豆固氮转移的比例均呈上升趋势。8月与10月的N 素转移率及核桃中N 素来自大豆固氮转移的比例的差异均不显著(P>0.05),6月的这2个指标值均显著低于8月和10月(P<0.05)。10月N 素转移率和核桃中N素来自大豆固氮转移的比例分别是6月的2.05、1.59 倍。

表5 核桃/大豆间作体系中N 素的转移比例Table 5 Percentage of walnut nitrogen transferred from soybeans %
3 结论与讨论
本研究结果表明,除10月外,间作模式下植株的总干物质量高于单作模式。随生育期推进,间作体系的N 素来自肥料比例的加权平均值、N肥利用率的加权平均值均大于单作体系。种植模式可以显著影响生育前中期的大豆生物固氮效率。间作模式中,核桃中N 素来自大豆固氮转移的比例不断增加。经综合分析可知,核桃和大豆间作有利于N 素的吸收利用,间作模式中N 素发生了由大豆向核桃的转移,间作模式表现出明显的总体优势。
3.1 间作对N 素吸收利用的影响
植株器官从肥料中吸收分配到的15N 量对该器官全氮量的贡献率(即器官中N 素来自肥料的比例)反映了植株器官对肥料15N 的吸收征调能力[20]。在本研究中,与单作模式相比,间作核桃中N 素来自肥料的比例在各生育期均显著降低(P<0.05),间作大豆中N 素来自肥料的比例和间作体系中N素来自肥料的比例显著升高。由此可知,与单作相比,间作模式中大豆对15N 吸收征调能力与间作体系中N 素来自肥料的比例的关系更为密切。相关研究结果表明,农林复合系统中林木与作物对N素的吸收利用因具有时间生态位上的分离而使间作体系中N素来自肥料的比例高于单作体系[17,21]。本试验中6月大豆中N 素来自肥料的比例在整个生育期中最高,并且高于核桃中N 素来自肥料的比例。6月之前,树体贮藏的N 素是核桃N 素需求的主要来源,对外源N 素的需求较少,此时正是大豆幼苗期,大豆需要吸收较多的土壤N 素来完成其营养生长和降低“N 阻遏”效应,当核桃进入吸收土壤N 素的高峰期时,间作模式下部分土壤N 素已被消耗,导致其土壤N 素含量低于单作模式,产生N 素竞争作用,进而能更大程度地利用土壤中所剩余N 素,最终使间作体系中N 素来自肥料的比例高于单作体系[22-23]。
本研究结果表明,种植模式、生育期及其交互效应均可显著或极显著地影响核桃各器官的15N分配率(P<0.05 或P<0.01),且10月之前的15N 分配率表现为地上部(叶、枝、主干)高于地下部(根),10月则相反。因为,6—8月是核桃的生长旺盛期,叶片进行光合作用需要大量N 素,使其对N 素的吸收征调能力增强,15N 分配率较高[24];10月核桃进入落叶期,地上部的N 素养分回流至地下部,被贮藏起来以备来年树体前期生长所用,这与王万才[25]的研究结果基本一致。已有研究结果证实,农林复合系统的N 素利用率高于单作[26],且林木对N 素的利用竞争弱于作物[27]。核桃为速生树种,当树体贮藏养分再利用结束后,大量土壤N 素被吸收以供其快速生长所需,但此时大豆对N 素的需求峰值已过,从而产生时间生态位互补[28]。再者,核桃和大豆的根系在空间上形成垂直分层,核桃为深根性树种,大豆根系处于土壤浅层,核桃根系能捕获未被大豆根系吸收而淋溶至土壤深层的养分,降低了土壤N 素淋溶率,在空间生态位上实现养分互补,进而提高间作模式下N 素的利用率[29]。
3.2 间作对生物固氮及N 素转移的影响
在间作系统中引入豆科作物,常常是因为其具有生物固氮能力。本研究结果表明,核桃落叶前期,间作对大豆生物固氮效率有显著影响(P<0.05)。前人的研究结果也表明,与豆科间作时,非豆科作物对土壤N 素的大量吸收可激发豆科作物的生物固氮潜力,增加豆科作物的固氮比例[30]。本研究中,6月大豆正处于苗期,因根系发育不完全,结瘤量和固氮作用极不稳定,使得单作大豆的固氮效率可能高于间作大豆[31]。当大豆完成苗期生长,进入生殖期后,其根系发育完全,并随着种间根系相互作用时长的增加,大豆结瘤量增加,加之间作模式中大豆与核桃的N 素竞争导致土壤中可利用N 素数量降低,迫使大豆通过生物固氮来获得更多大气中的N 素,以此满足自身生长[32]。因而,6月后间作大豆的固氮效率高于单作大豆,表明核桃与大豆间作有利于大豆生物固氮效率的提高。然而,有研究结果表明,间作虽然提高了豆科作物的固氮效率,却降低了其干物质量[33]。这与本试验结果类似,可能与试验过程中未施N肥有关,植物生长后期共用N 素匮乏,物种间的竞争作用加强,直接影响了间作物种干物质量的积累,使得间作体系中地上部和地下部的干物质量均较单作时低,其中大豆尤为显著(P<0.05),这可能也与核桃阻挡了自然光,导致大豆光能截获量降低相关。但间作体系中的总干物质量高于单作体系,间作物种通过根间相互作用,实现N素营养互补,以此弥补不施N 肥导致的核桃干物质量降低。另外,通过根间相互作用,间作能消减因土壤低N 含量引起的大豆“N 阻遏”并刺激大豆固氮能力的增强,达到为核桃提供N 素的目的[34]。
N 素在地下部由豆科固氮作物向非豆科作物转移是十分重要的。褚贵新等[18]报道了水稻与花生间作模式中N 素的转移情况,表明因为间作模式中根系相互竞争能力、接触程度及土壤N 素含量等因素的不同,间作物种间N 素转移方式存在差异,N 素转移量受到影响。Fujita 等[35]的研究结果也表明,间作模式中2个物种根系相互作用能促进N 素转移。在本研究中,随着生育期推进,N 素转移率和核桃中N 素来自大豆固氮转移的比例逐渐增加,这可能是因为核桃和大豆根系不断生长、相互交织,形成了较多接触面,从而使核桃能够从大豆获得更多通过生物固定的大气N素。目前,认为农林间作模式中固氮植物的N 素转移有2 种方式,即非固氮植物根系吸收固氮植物释放于土壤中N 素的间接方式,和通过连接间作物种根系的真菌菌丝进行N 素转移的直接方式[36]。本研究中仅对核桃N 素来自大豆固氮转移的比例进行了分析,但系统物种根系间的相互作用尚未明确,后续将对核桃和大豆间作系统地下部N 素转移方式进行深入研究。
