山新杨LTP家族基因生物信息学及表达模式分析
2022-03-02吴宇航张雅譞曹竞丹石晶静
孙 乾 吴宇航 张雅譞 曹竞丹 石晶静 王 超*
(1. 东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040;2. 东北林业大学林学院,哈尔滨 150040)
高盐土壤会导致植物的生长代谢紊乱,进而也会影响植物的正常生长发育过程。干旱会对植物的生化代谢过程产生影响,进而阻碍植物的正常生长发育。研究林木材性和抗性形成的分子机制,鉴定重要基因,对于林木在抗盐、抗干旱育种方面具有重要意义。
(lipid transfer protein)是研究较少的一类植物基因家族,广泛分布于植物组织和细胞。1975年,基因首次在马铃薯(L.)块茎内被发现,并被证实在植物细胞膜上有可以与磷脂特异性结合并将其转运的蛋白,因此被命名为脂质转运蛋白。它作为一类植物蛋白质,在植物的生长发育过程中发挥着极其重要的调控作用。基因相关研究表明,LTP 蛋白的三级结构中具有一个独立的疏水腔,因此可以推测,通过疏水腔和磷脂等物质的作用,可实现脂质的跨膜运输。有关研究表明基因除了脂质运输功能外,还具有角质膜的生成、生物信号传导的功能。
基因作为一种抗性基因,在植物的生长发育方面具有很重要的作用,因此将具有抗逆功能的基因过量表达可以提高植物抵抗非生物胁迫的能力。邢仪利用马铃薯转基因研究发现该基因具有抗盐、抗旱功能。巴西学者Moraes等利用水稻(L.)定量研究LTP1 类蛋白的11种遗传异构体的耐性(尤其是耐盐性)和抗病基因型;臧庆伟发现为小麦(L.)水分胁迫诱导表达的基因,二者在转基因拟南芥()中的超表达可以提高转基因植株的抗旱性;李晋将转入到TI1068 烟草(L.)品系,发现在高盐和脱水的环境下,转基因烟草的萌发率和根长高于野生型烟草,说明转基因烟草对干旱和高盐 逆 境 具 有 更 大 的 耐 受 力。Jülke 等 发 现 转和基因水稻株系在含有NaCl 的培养基上生长状况更好,说明这两个基因均具有耐盐功能。另外,有相关研究表明基因的表达受多种激素的诱导,程朝泽对基因进行启动子分析发现,基因能够响应盐旱等逆境条件、激素处理诱导。刘勇发现脱落酸(ABA)、乙烯(ET)、茉莉酸甲酯(MeJA)和HO能显著上调基因的表达水平,赤霉素(GA)处理后其表达水平出现下调,且基因DNA 序列的启动子存在干旱、低温等多种(非)生物胁迫响应元件和MeJA等激素响应元件。
山新杨(var.)是由山杨()和新疆杨(var.)杂交选育而成的杨树品种,具有生长迅速、抗逆性强、树干挺直、树型美观等特点,品质优良,很受人们重视,具有广阔的发展前景。本研究以家族基因为研究对象,进行蛋白性质分析、多序列比对分析、进化树分析及qRT-PCR 分析,初步了解家族基因在不同组织及非生物胁迫条件下的表达模式,为研究山新杨木质部发育及抗非生物胁迫性状形成的分子基础研究及分子育种提供理论数据和研究材料。
1 材料与方法
1.1 实验材料
本研究所使用的植物材料选自于东北林业大学林木遗传育种国家重点实验室保存的山新杨组培苗,培养温度为25 ℃,光周期为16 h 光照/8 h 黑暗,光照强度为400 µmol·m·s。山新杨组培苗培养4 周后将其移至营养土中,置于温室人工培养,人工培养的营养土壤中(黑土)∶(蛭石)∶(珍珠岩)=5∶3∶2,生长条件为,光周期为光照:16 h/黑暗:8 h,光照强度为400µmol·m·s,温度为(22±2)℃,相对湿度为65%~75%。生长8 周左右,在200 mmol·LNaCl,20% PEG6000 胁迫处理0、6、9、12、24、48 h 后进行取材,每个处理重复3 次,每次重复含3 株,以水处理为对照。5 年生野生山新杨用于进行应拉木诱导处理,将处理组的树干全部朝同一方向弯曲,将树干顶端部分用绳子和胶带固定于相邻树上,使树干部分大致倾斜45°,应拉木处理为5 棵1 组,未处理对照组为5 棵1 组,设置3 次生物学重复,待2 周之后取材。倾斜上部木质部为应拉木(TW),下部为对应木(OW),以直立木(NW)为对照,刮取正在发育的木质部。所取材料用液氮处理,-80 ℃保存备用。
1.2 实验方法
1.2.1家族基因生物信息学分析
利用在线软件ExPASy 提供的ProtScale(http://web.expasy.org/cgi-bin/protscale/)对家族基因进行亲水性预测。利用在线软件SOP‐MA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)预测分析山新杨LTP 家族蛋白二级结构。利用软件SWISSMODEL(https://www.sw issmodel.expasy.org/interac‐tive/DTDUCp/models/)预测PdbLTP 家族蛋白的三级结构。用BioEdit 软件进行多序列比对,将结果导出后,利用MEGA 6.0 软件进行分析,选择NJ 方式构建进化树。
1.2.2 RNA提取及cDNA合成
引物及样品的测序均委托吉林省库美生物科技有限公司(www.comatebio.com)进行。主要试剂包 括200 mmol·LNaCl(上 海 生 工);20%PEG6000(Biosharp);通用植物总RNA 提取试剂盒(Bioteke Corporation);反转录试剂盒(TaKaRa);qRT-PCR反应试剂盒(全式金)。
使用CTAB 法提取山新杨中的总RNA,利用反转录试剂盒(TaKaRa,日本)进行cDNA 的合成。反应体系为20 µL∶2 × TS Reaction Mix 10 µL,TransScript ⑧RT/RI Enzyme Mix 1 µL,Anchored Oligo(dT)18 Primer(0.5 µg·µL)1 µL,Total RNA 50 ng~5 µg,gDNA Remove 1 µL,补充RNase-Free Water 至20 µL。反应条件∶在PCR 仪(Bi-ometra)中,42 ℃条件下,反应30 min;85 ℃条件下,反应5 s。实验结束后,将cDNA于-20 ℃冰箱保存备用。
1.2.3 qRT-PCR分析
根据基因组中鉴定的山新杨家族CDS 序列选取200 bp 左右的序列长度设计特异性定量引物(如表1)选择-和作为内参基因。qRT-PCR 反应体系为特异性定量F,R primer各1 µL、模板2 µL、去离子水7 µL 和2×Power SYBR Green PCR master mix 10 µL(Toyobo Co.,Osaka,Japan)总体系为20µL,3 次技术重复,反应程序为94 ℃预变性3 min,94 ℃变性12 s、58 ℃退火30 s、72 ℃延伸40 s、78.4 ℃读板1 s,共45 个循环,72 ℃延伸7 min。在Bio-Rad(Hercules.CA.USA.)上完成该反应。读数分析其数据,运用2方法进行相对表达量分析。
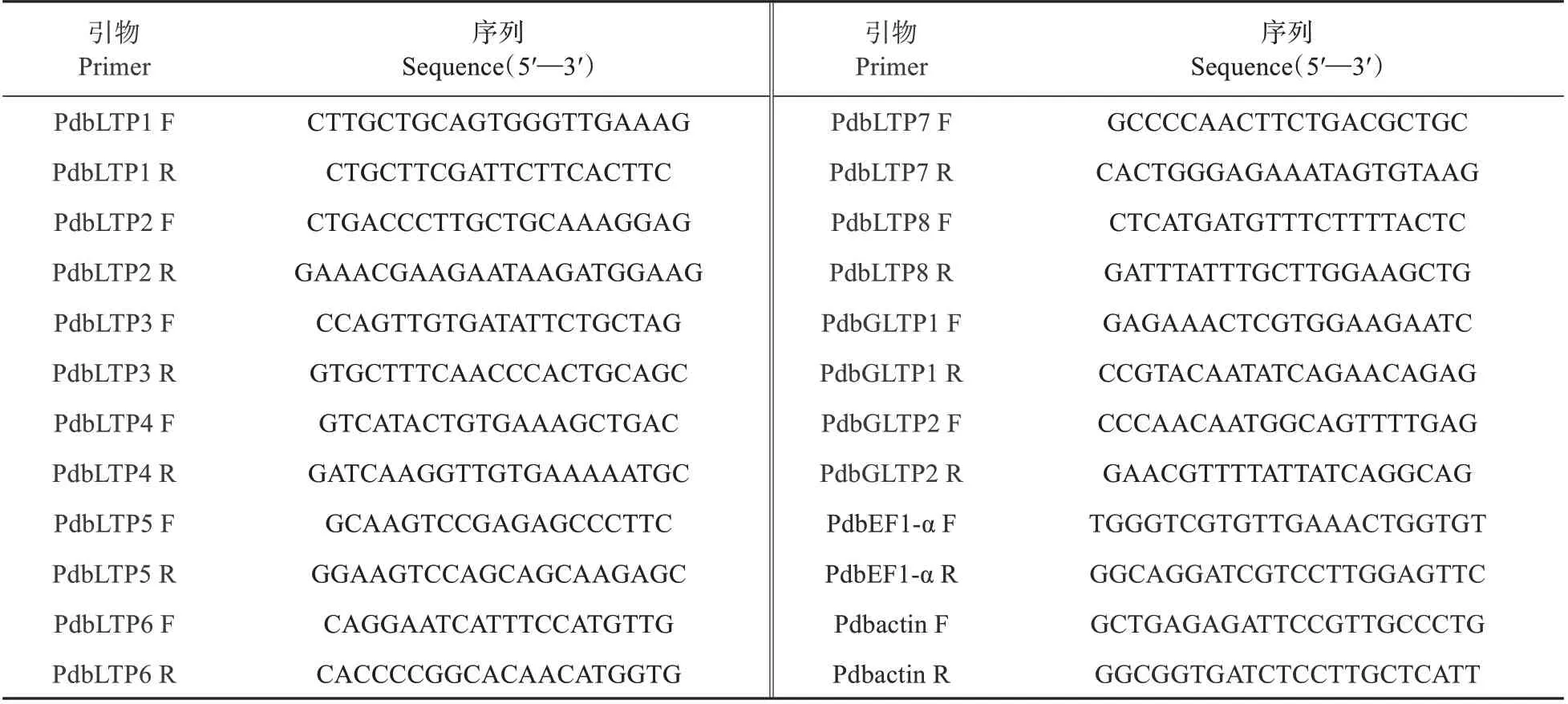
表1 山新杨LTP家族基因定量引物序列Table 1 Quantitative primer sequences for LTP family genes of Populus sibiricum
1.2.4 数据处理
数据统计与分析采用Excel 2016(Microsoft Office)与SPSS 22(IBM)软件进行,利用检验进行方差分析中显著性检验。将家族基因表达模式分析结果分别进行两两比较,不同材料及处理分别进行独立样本检验,选择Analyze 模式进行分析,<0.05表明结果显著,<0.01表明结果极显著。
2 结果与分析
2.1 PdbLTP基因生物信息学分析
2.1.1基因序列信息
山新杨基因的核苷酸数量多数集中在294~396 bp,氨基酸数集中在97~131 aa;而基因的序列长度较基因长,两个基因分别为482和720 bp(见表2)。

表2 PdbLTPs基因序列统计表Table 2 PdbLTPs genes sequence statistics table
2.1.2 PdbLTPs蛋白亲疏水性
由图1 可知,亲水性分析结果显示山新杨LTP蛋白既有亲水性区域也有疏水性区域,图中正值的高峰部分即是疏水性区域,负值的高峰部分即是亲水性区域。可以明显看出,疏水性区域多于亲水性区域,说明PdbLTPs 蛋白家族均具有疏水性,推测其为跨膜蛋白。
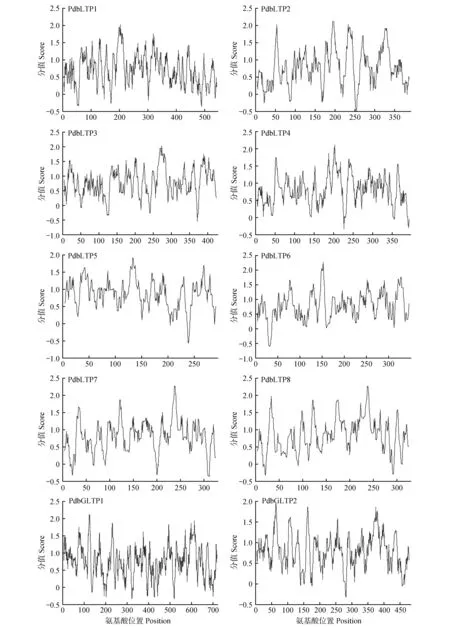
图1 PdbLTPs蛋白亲水性分析Fig.1 Hydrophilic analysis of PdbLTP proteins
2.1.3 PdbLTPs蛋白高级结构分析
PdbLTP 家族蛋白质二级结构预测分析发现,PdbLTP1 各二级结构的峰值均在60~80 nm,Pd‐bLTP2 各二级结构的峰值区域处于40~100 nm,PdbLTP3 的二级结构中延伸连峰值最大,且峰值集中在20 nm,PdbLTP4 二级结构峰值集中在40~80 nm,相比之下PdbLTP5、PdbLTP6、PdbLTP7、Pd‐bLTP8 蛋白的二级结构都有多次不同大小的峰值(见图2);为进一步进行分析,进行了蛋白质三级结构预测,预测结果显示,山新杨LTP 家族蛋白均将进行不同程度的折叠,其中,PdbLTP1、Pd‐bLTP6、PdbLTP8 含有-螺旋和无规则卷曲和-转角,PdbLTP4、PdbLTP5、PdbLTP7、GLTP1含有-螺旋和无规则卷曲和-转角和-折叠,PdbLTP2含有-螺旋和无规则卷曲和-折叠,PdbLTP3含有-螺旋和-转角,PdbGLTP2 含有-螺旋和无规则卷曲(见图3)。PdbLTP 家族蛋白由螺旋、折叠、转角和无规则卷曲相互盘绕、折叠三维构象,而Pdb‐GLTP 蛋白则是由螺旋、折叠、转角盘绕、折叠三维构象。

图2 PdbLTPs蛋白二级结构预测分析Fig.2 Secondary structure prediction of PdbLTP Proteins

图3 PdbLTPs蛋白三级结构预测分析Fig.3 Tertiary structure prediction of PdbLTP proteins
2.1.4 PdbLTPs蛋白多序列比对及系统进化分析
多序列比对结果显示PdbLTP 蛋白在保守位置具有8 个Cys 残基,是LTP 家族蛋白重要结构(见图4)。进化树分析显示,PdbLTP1、PdbLTP2、PdbLTP3、PdbLTP4 与响应盐胁迫的BpLTP4 基因蛋白在同一分支。因此,推测这些基因也具有耐盐功能。PdbLTP5 与AtLTP3 和AtLTP4 同源性较高,PdbLTP6、PdbLTP7、PdbLTP8 与TaLTP 也具有同源性(见图5)。研究显示和具有耐盐能力,因此推测基因具有相似功能。

图4 PdbLTPs蛋白多序列比对分析Fig.4 Sequence alignment analysis of PdbLTP proteins

图5 PdbLTPs蛋白进化树分析Fig.5 Phylogenetic tree analysis of PdbLTP proteins
2.2 PdbLTPs基因表达模式分析
2.2.1 重力处理表达模式分析
在人工弯曲模拟重力刺激处理后,qRT-PCR分析结果显示、、、在应拉木中的表达量最高,、在对应木中的表达量最高,这些山新杨基因响应人工弯曲和重力刺激。和对人工弯曲和重力刺激处理响应不明显(见图6)。

图6 重力处理下PdbLTPs基因表达模式分析*P<0.05,**P<0.01,下同Fig.6 Expression pattern analysis of PdbLTP genes under gravity processing*P<0.05,**P<0.01,the same as below
2.2.2 组织特异性表达模式分析
组织特异性表达分析结果显示,、、和均在茎中的表达量最高;在根中的表达量最高;在叶中的表达量最高;在各组织中表达差异不大(见图7)。

图7 PdbLTPs基因组织特异性表达模式分析Fig.7 PdbLTPs tissue specific expression pattern analysis
2.3 NaCl胁迫下PdbLTPs基因表达模式分析
NaCl 胁迫处理下基因表达量分析结果显示,基因受盐胁迫不同程度的诱导,且响应盐胁迫的时间不相同。其中基因表达量在NaCl 胁迫9 h 后高度上调表达,基因的表达量随胁迫时间的增长而不断上调在NaCl 胁迫9 h 达到峰值。、、表 达 量 在NaCl 胁 迫6 h 时 达 到 峰 值,、基因表达量在NaCl 胁迫3 h 时表达上调,随后其表达量显著下调表达。基因在NaCl 胁迫3 h 时下调,随着胁迫时间变长,其表达量上调并在NaCl胁迫24 h时表达量达到峰值(见图8)。说明基因响应盐胁迫,响应模式不同。
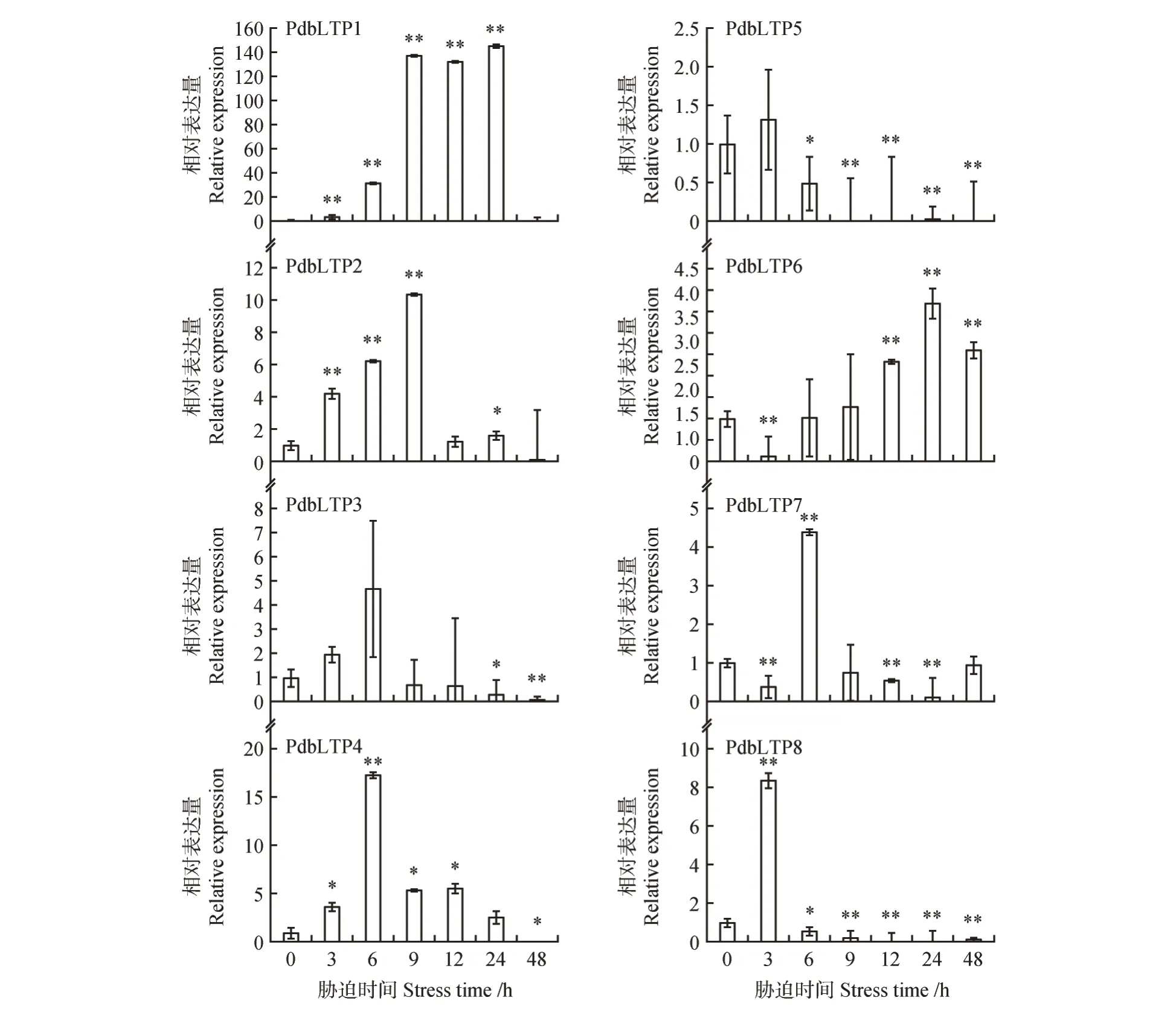
图8 NaCl胁迫处理下PdbLTPs基因表达模式分析Fig.8 Expression pattern analysis of PdbLTP genes under NaCl stress
2.4 PEG 胁迫处理下PdbLTP 家族基因表达模式分析
PEG 胁迫处理下家族基因表达量分析结果显示:家族基因的表达对PEG 胁迫处理均有不同程度的应答。其中,和基因具有相似的表达模式,其表达量随着胁迫时间变长而不断上调,在PEG 胁迫9 h时其表达量达到峰值。基因表达量在PEG 胁迫24 h时表达量最高,、基因表达量随着PEG 胁迫时间变长而下调。的表达量在PEG 胁 迫 处 理24 h 时 最 高,和基因具有相似的表达模式,在PEG 胁迫处理9 h 时出现特异表达高峰。的表达变化不显著。结果说明,家族基因可以不同程度响应干旱胁迫(见图9)。

图9 PEG胁迫处理下PdbLTPs基因表达模式分析Fig.9 Expression pattern analysis of PdbLTP genes under PEG stress
3 讨论
高等植物脂质转运蛋白LTP 是一种分子量为3~9 kDa 的小分子碱性蛋白,占可溶性蛋白的4%左右。这种在植物的细胞膜上可以与脂质特异性结合的蛋白在植物中一直发挥着不可替代的调控作用,相关研究已经表明,在LTP 蛋白的三级结构中有一个独立的疏水腔,可能用以实现脂质的跨膜运输。有研究显示LTP 参与疏水性相互作用,对细胞壁扩张产生重要的影响。本研究对PdbLTP 的亲水性预测分析结果也显示该蛋白为疏水性蛋白。其高级结构存在疏水腔结构,说明山新杨LTP 蛋白具有LTP 家族蛋白的结构特征,在参与疏水性相互作用,利用疏水腔进行脂质跨膜运输中具有一定功能。进化树分析显示,基 因 分 别 与、、及耐盐基因具有较高同源性,研究显示白桦()基因和小麦基因具有耐盐性,因此推测基因具有耐盐功能。
为了进一步分析山新杨基因可能参与的功能,对其进行了不同组织的响应重力刺激,响应盐和旱胁迫的表达模式分析。曾有学者研究过水曲柳(Rupr.)的基因,发现基因在木质部的表达量最高时对应拉处理有响应。对毛果杨(Torr.&Gray)应拉木中特异表达的基因进行进一步验证,发现其在一定程度上参与木质部发育调控。之前有研究者对光皮桦(H.Winkl.)基因进行分析时发现,当基因在茎表达量较低时,应拉木相关检测数值显著低于直立木。本实验对基因进行重力处理表达模式分析,结果显示、、、在应拉木中的表达量最高。对组织特异性表达进行分析,结 果 表 明、、、均在茎中的表达量最高,在根中的表达量最高,和在叶中的表达量最高。在重力刺激下,表达量变化不明显的与,在茎中的表达量也较低,这符合上述研究的表达规律,而、、则很可能影响茎与木质部的发育过程。有人曾经就沙柳()的基因组织特异性表达结果分析,发现基因表达量在某些部位高表达时的确与发育有一定联系。
刘群等对腊梅()叶片中3 个基因应答非生物胁迫的情况进行了qRT-PCR 分析和比较,发现这些基因受干旱与Na‐Cl 处理的影响程度不同,李倩发现过表达的转基因拟南芥耐盐性有明显提高,表明小麦基因与植物的高盐胁迫耐性相关。PEG 胁迫对同一植物的不同器官影响不同,很多学者发现,一些植物对PEG 胁迫做出一定程度上的响应,而这往往与其所含有的基因蛋白有关,比如小麦干旱响应蛋白WZY2 与WZY3-1,和大豆()中的GsPIP1-4 蛋白对干旱条件的响应,还有杨树(spp.)的多聚泛素蛋白对干旱的响应。本研究利用qRT-PCR 分析基因在盐胁迫和干旱胁迫下的表达模式,发现虽然不同基因在响应两种胁迫的表达量和表达时间均有不同程度的差异,但最终都表明基因的表达响应盐胁迫和干旱胁迫。这进一步说明基因家族中的多个基因都参与到植物的耐盐和耐旱调控中。
结合聚类分析的结果可以看出,系统进化上聚为一组的基因具有相似的表达模式,如、、聚为一组,它们在受到人工弯曲和重力刺激时响应表达,在木质部发育相关组织高度表达,在盐胁迫和干旱胁迫下受诱导。、、聚为一组,他们在应力木中变化不显著,在木质部发育相关组织中表达量较低,在盐胁迫条件下,和在处理6 h出现特异表达高峰。说明聚为一组的基因具有相似的表达模式,乃至可能具有相似功能。、、和木质部发育相关,且在环境刺激(盐、旱胁迫)中起到一定的调节作用。、、则是另外响应模式的盐响应基因。
本研究对山新杨家族基因进行生物信息学分析及其表达模式分析,分析家族基因中不同基因的不同序列特征及每一种成员潜在的功能,增加对这一类广泛存在的植物基因家族的了解,为林木优质、抗逆育种等研究提供理论依据。
4 结论
通过对山新杨基因的生物信息学研究以及对其表达模式分析,发现PdbLTPs 蛋白具有疏水性,不存在跨膜区域。系统进化分析结果显示PdbLTPs 蛋白与已知抗逆相关的LTP 进化关系较近。在山新杨茎中表达量较高,且受人工弯曲和重力刺激诱导,在应拉木中高度表达,这些基因与木质部发育相关。大部分基因对NaCl和PEG 处理下的逆境有不同程度的响应,表明基因很可能作为重要因子参与植物抗逆响应的过程,提高植物抗逆的能力。
