大鼠牙囊细胞成骨分化过程中Notch信号表达情况*
2022-02-12邝惠芳
罗 文,邝惠芳,王 婧
(1.海南医学院第一附属医院口腔科,海口 570102;2.海南医学院口腔医学院,海口 571199)
来源于牙体组织的牙源性干细胞/祖细胞用于牙周疾病模型中具有良好的可行性和可预期性[1]。牙囊细胞(dental follicle cells,DFC)是牙周组织的前体细胞,其中包括牙槽骨成骨细胞、成牙骨质细胞和牙周成纤维细胞[2]。DFC成骨分化的不同阶段和相关基因表达与众多信号通路相关,如Notch、转化生长因子-β(TGF-β)/骨形态发生蛋白(BMP)和WNT通路[3]。Notch通路通过Notch受体与膜结合型配体,如Jagged-1结合介导相邻细胞间的信息交流。位于细胞表面的受体Notch和配体(如Jagged-1)结合后被γ分泌酶分解。胞内活化域(NICD)释放并易位到细胞核中与RBP-JK结合,从而调控细胞增殖与分化[4]。本研究探讨在大鼠DFC成骨分化时Notch信号的表达情况变化,现报道如下。
1 材料与方法
1.1 材料
SD大鼠购自海南医学院药物安评中心;α-MEM培养基、胎牛血清、胰蛋白酶均购自美国Hyclone公司;青霉素、链霉素、抗坏血酸、Ⅰ型胶原酶、Triton、牛血清清蛋白(BSA)、茜素红、二脒基苯基吲哚(DAPI)均购自美国Sigma公司;小鼠抗大鼠波形蛋白(Vimentin)单克隆抗体购自美国 Thermo公司,小鼠抗大鼠细胞角蛋白-14(CK-14)单克隆抗体购自英国Abcam公司;山羊抗小鼠异硫氰酸荧光素(FITC)-免疫球蛋白G(IgG)购自武汉博士德公司;Trizol试剂、引物Notch 1和β-actin均购自上海生工公司;M-MuLV逆转录酶试剂盒购自美国Fermentas公司;定量逆转录聚合酶链反应(qRT-PCR)试剂盒SYBR Green Ⅰ kit购自瑞士罗氏公司;OLYMPUS IX71O倒置相差荧光显微镜与OLYMPUS CKX41倒置显微镜均购自日本HITACHI公司;LightCycler PCR 2.0系统购自瑞士罗氏公司。
1.2 方法
1.2.1大鼠DFC原代培养
取出生后6~7 d的SD大鼠的乳鼠,无菌条件下将下颌骨切除放置于磷酸盐缓冲溶液(phosphate buffered solution,PBS)中。体视显微镜下用显微镊分离下颌第一磨牙的牙胚,机械分离牙囊组织,放置于含1 %双抗的α-MEM培养液中,用眼科剪彻底剪碎。37 ℃、放置在60 U/mL Ⅰ型胶原酶中消化60 min。加入4 mL细胞培养液(α-MEM+10%胎牛血清+100 U/mL青霉素+100 μg/mL链霉素+100 mg/mL抗坏血酸),并将大鼠DFC转移至T-25细胞培养瓶中,放置在37 ℃、5%二氧化碳培养箱中12 h让细胞贴壁。细胞融合后用0.25%胰酶消化传代。取第3代大鼠DFC作为实验细胞。实验过程中所有动物实验操作符合海南医学院伦理及操作要求。
1.2.2大鼠DFC来源的鉴定(Vimentin、CK-14细胞免疫荧光染色)
1.2.2.1细胞爬片的制作
取第3代对数生长期大鼠DFC消化制备成细胞悬液1×105/mL;,加入6孔细胞培养板中1×105个/孔,制作成爬片。
1.2.2.2细胞固定
待爬片上的细胞长至50%~60%,用4%多聚甲醛固定10 min。用1%Triton溶液配制的3%BSA封闭液中封闭作用30 min(37 ℃湿盒中)。
1.2.2.3孵育一抗
用1% BSA溶液按抗体说明书所示比例稀释抗体,Vimentin(1∶250)、 CK-14(1∶100),37 ℃湿盒中孵育2 h。
1.2.2.4孵育二抗
用1%BSA溶液按1∶200比例稀释FITC结合的羊抗鼠IgG二抗,37 ℃湿盒中孵育1 h。
1.2.2.5细胞核染色
每张爬片滴加200 μL的DAPI(1 μg/mL)溶液浸染2 min,双蒸水洗2次,每次3 min。中性树胶封片。
1.2.3成骨分化诱导
对照组继续用基础培养液培养,成骨诱导组加入地塞米松为基础的成骨诱导液继续培养21 d,3 d换1次诱导液,镜下观察出现不规则结节即可。弃成骨诱导液,PBS洗3遍,4%多聚甲醛固定30 min,茜素红染色15 min对分化细胞进行染色,显微镜下采图。
1.2.4qRT-PCR
使用Trizol试剂分别分离对照组和成骨诱导组总RNA,采用紫外分光光度计测定RNA的量和纯度。使用M-MuLV逆转录酶试剂盒进行cDNA合成。qRT-PCR用SYBR Green Ⅰ kit和LightCycler PCR 2.0系统及LightCycler4.05软件计算域值循环数。引物的设计和合成由上海生工公司完成,序列见表1。以β-actin为内参照,qRT-PCR检测Notch1基因表达。反应结束后由软件分析扩增曲线和产物溶解曲线,按相对定量法自动计算目的基因mRNA拷贝数。
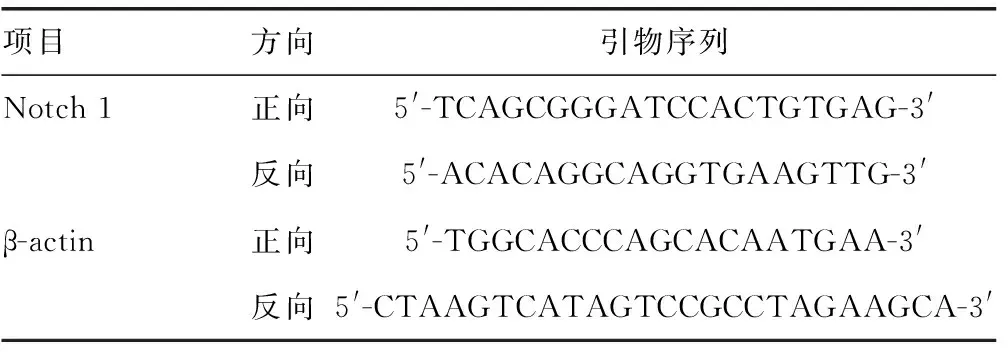
表1 Notch 1与β-actin引物序列
1.3 统计学处理
采用SPSS 13.0统计软件进行数据分析,计数资料以例数或率表示,组间比较采用χ2检验。以P<0.05为差异有统计学意义。
2 结 果
2.1 大鼠DFC的分离、鉴定及成骨分化潜能
大鼠DFC的原代培养3 d左右细胞从组织块周围爬出,见图1A。DFC大部分呈纺锤状、长梭形,少部分为多角形,其来源于外胚层间充质,具有典型的间充质成纤维细胞形态,见图1B。CK-14染色为阴性,见图1C。Vimentin几乎表达于所有大鼠DFC,染色阳性,见图1D。表明获取的DFC是间充质干细胞,而无上皮细胞。混杂在大鼠DFC原代中的上皮细胞传至第3代时大鼠DFC已基本被纯化。用含有地塞米松的成骨诱导液诱导大鼠DFC 28 d后用茜素红染色液染色可见许多红色矿化小结节生成,见图1E。

A:组织块原代培养DFC;B:第3代DFC;C:CK-14抗体免疫荧光检测;D:Vimentin抗体免疫荧光检测;E:第3代DFC成骨诱导后茜素红染色。图1 大鼠DFC的分离、鉴定及成骨分化潜能
2.2 经成骨诱导后DFC的Notch 1 信号表达情况
与对照组比较,成骨诱导28 d后DFC的Notch 1基因表达下降,差异有统计学意义(P<0.05),见图2。

*:P<0.05,与对照组比较。图2 经成骨诱导后DFC的Notch 1 信号表达情况
3 讨 论
DFC作为骨组织工程的种子细胞之一,具有取材方便、多向分化能力强等优点,在骨修复研究中具有良好的应用前景[5]。牙囊是包绕于牙齿发育期成釉器和牙乳头周围的疏松结缔组织,来源于外胚层间充质,在牙齿萌出和牙周组织形成中具有重要作用。DFC是成骨细胞和成牙骨质细胞的前体细胞,DFC中有碱性磷酸酶、Ⅰ型胶原、骨钙蛋白、骨涎蛋白等成骨细胞标志物表达,有牙骨质附着蛋白等成牙骨质细胞标志物表达,经诱导分化可形成牙槽骨和牙骨质[6]。DFC与上皮根鞘细胞和牙髓细胞混合,可以诱导DFC成骨及成牙骨质方向分化,形成骨样组织和成牙骨质组织[7]。本研究成功分离、培养、鉴定了DFC。DFC的鉴定包括细胞形态鉴定和细胞表面特异性标志物检测。DFC大多为呈梭形的成纤维细胞样,呈纺锤状结构,漩涡状排列,贴壁生长。DFC来源于外胚间充质,可检测到间充质干细胞表面标志物——Vimentin染色阳性,同时上皮细胞表面标志物——CK-14染色阴性。利用茜素红染色液对经成骨诱导的DFC进行染色可见若干红色矿化小结节生成,表明DFC在合适的成骨诱导条件下可以向成骨分化,与相关研究结果一致[8]。
Notch信号通路广泛存在于脊椎动物和非脊椎动物,在进化上高度保守,通过相邻细胞之间的相互作用调节细胞、组织、器官的分化和发育[9]。相邻细胞可以通过Notch受体与配体的结合传递Notch信号。Notch信号是相邻细胞之间通讯进而调控细胞发育分化的重要通路[10-11]。Notch 1作为Notch信号通路的主要受体,与其配体——Jagged 1及下游靶基因——Hes 1均为调控细胞分化的关键信号通路因子。在胶原诱导性关节炎(CIA)大鼠模型中,CIA组大鼠关节Notch 1呈强阳性表达,同时Jagged 1、Hes 1表达也显著上升,高于对照组和中药(傅山风湿外治方)控制组,差异均有统计学意义(P<0.05),说明Notch 1/Jagged 1信号通路在诱发CIA的过程中得到激活,参与了CIA的发生、发展。通过药物控制能显著降低Notch 1、Jagged 1、Hes 1的表达,抑制CIA大鼠中Notch 1/Jagged 1信号通路的活化[12]。表明Notch 1与配体Jagged 1结合后能影响其下游靶基因Hes 1的转录,Notch 1的表达水平能影响Notch 1/Jagged 1信号通路的激活或抑制,从而调控多种炎症因子的表达,如白细胞介素-6、γ干扰素等,发挥炎症调控作用[13]。
Notch 1 是DFC维持未分化状态的标记物之一,成骨分化过程中Notch信号在DFC中被激活[14]。Notch信号通路被激活不仅影响DFC成骨分化,而且影响BMP/DLX3信号通路(成骨分化相关通路)的激活,Notch信号通路可能通过负反馈回路调节DFC的成骨分化[15]。有研究发现,抑制Notch信号通路后可抑制骨肉瘤细胞MG63成骨分化,联合调控Notch信号通路与分化抑制因子1靶基因的表达可遏制骨肉瘤异常成骨分化,诱导其向正常成骨分化,促使异常中断的成骨分化重新发生[16],说明Notch信号在骨肉瘤细胞MG63成骨分化中具有重要作用。有研究将人牙周膜干细胞经成骨诱导分化后发现,整合素金属蛋白酶10(ADAM10)过表达组矿化结节形成量、成骨相关基因ALP、Runx2、OCN mRNA表达水平及Notch信号通路相关基因——Notch1、NICD、Hes mRNA和蛋白表达水平均显著降低,差异均有统计学意义(P<0.05),表明过表达ADAM10可促进人牙周膜干细胞增殖并抑制其成骨分化,其机制可能与Notch信号通路的激活有关[17]。在骨质疏松症大鼠动物模型中,经成骨诱导后骨质疏松症脂肪干细胞形成的矿化结节数量、Runx2、Notch1表达水平均比正常脂肪干细胞低,说明骨质疏松症脂肪干细胞成骨能力比正常脂肪干细胞弱,且Notch信号通路受到抑制[18]。本研究结果显示,与对照组比较,DFC经28 d成骨诱导后Notch 1基因表达下降,表明在成骨诱导环境下Notch信号通过下调影响DFC向成骨方向分化,与相关研究结果一致。然而Notch信号通路的各级信号因子如何相互作用精确调控DFC的成骨分化仍需进一步研究。
综上所述,DFC表达间充质干细胞表面标志物和Notch信号,具有间充质干细胞的特性。在成骨分化过程中Notch信号表达量下调,通过负反馈回路调节DFC的成骨分化。
