抗条锈病普通小麦-中间偃麦草7St二体异附加系的分子细胞遗传学鉴定
2021-12-08于军伟王斯文姚广平赵继新陈春环吉万全王长有
于军伟,王斯文,姚广平,赵继新,陈春环,吉万全,王长有
(1.西北农林科技大学农学院,陕西杨凌 712100; 2.农业部作物基因资源与种质创新陕西科学观测试验站,陕西杨凌 712100; 3.三原县农业科学技术中心,陕西三原 713800)
小麦野生近缘植物携带有大量的优异基因,是栽培小麦遗传改良的重要基因资源库[1]。利用远缘杂交技术向栽培小麦品种转移近缘种属的优异基因是创造小麦新的变异类型、扩宽小麦遗传基础、丰富亲本遗传多样性、创新小麦育种资源的重要手段[2]。
中间偃麦草(Thinopyrumintermedium,2n=6x=42,JJJSJSStSt)属于小麦族(Triticeae)偃麦草属(Elytrigia),为多年生异花授粉植物,主要分布于高加索、中亚的东南部[3]。其抗旱性、耐盐性和抗寒性都极强,许多农艺性状表现良好,且几乎不受小麦锈病和白粉病的侵害,对黑穗病、根腐病、茎腐病、叶枯病等病害也表现为高抗。中间偃麦草是偃麦草属中最早与小麦成功杂交的远缘材料,为小麦种质创新和品种改良提供了大量的优异基因,是小麦遗传改良的重要资源库[4]。
目前关于中间偃麦草的基因组组成争议较大,普遍认为中间偃麦草的基因组组成是JJJSJSStSt[5],为部分同源异源六倍体。Kruppa等[6]利用多色GISH(multi-color GISH, mc-GISH)技术,以百萨偃麦草(Thinopyrumbessarabicum)和拟鹅观草(Pseudorogneriastrigosa)的基因组DNA为探针,证明中间偃麦草是由19条J染色体、9条Js染色体和14条St染色体组成。前人研究表明,中间偃麦草J染色体组可能来源于原始的二倍体长穗偃麦草或百萨偃麦草,St染色体组可能起源于拟鹅观草[7]。关于JS基因组的来源,Wang等[8]研究认为,Js基因组供体先与J基因组经过了一次杂交加倍,然后与原始的St基因组供体杂交加倍,形成Js基因组,最后形成了当今的中间偃麦草。崔 雨等[5]提出中间偃麦草的Js染色体组可能与簇毛麦有较近的亲缘关系,并将其表示为Jvs,这为中间偃麦草的起源又带来了新的观点。
小麦条锈病是由条锈菌(Pucciniastriiformisf.sp.tritici(Pst))引起的影响小麦生产的真菌类病害,可使小麦年产量减少20%~30%,发病严重时甚至颗粒无收[9]。小麦条锈病在我国曾数次流行,最近一次于2019年发生在西北地区,并迅速蔓延至全国[10-11]。研究表明,种植抗病品种是最经济、最有效的控制小麦条锈病的策略[12]。前人已经发现了多个小麦条锈病抗性基因,并广泛用于小麦的抗病育种[13]。然而,由于条锈病病原菌毒性小种的变异迅速,大多数品种的抗病性逐渐降低或最终丧失抗病性[14]。因此,培育新的抗病品种对于小麦育种研究是非常迫 切的。
ES-24是普通小麦品种阿勃缺体系与普通小麦-中间偃麦草部分双二倍体远中4杂交和连续自交产生的后代,田间对条锈病表现为高抗。本研究利用细胞学、原位杂交、分子标记等技术,结合形态学和条锈病抗性调查,对ES-24进行综合鉴定,以期为ES-24在小麦遗传改良中的应用提供依据。
1 材料与方法
1.1 供试材料
供试材料包括中间偃麦草(2n=6x=42, JJJSJSStSt)、中国春(2n=6x=42,AABBDD)、百萨偃麦草(2n=14,JJ 或EbEb)、拟鹅观草(2n=14,StSt)、小麦-中间偃麦草二体异附加系ES-24、八倍体亲本远中4和阿勃缺体系,以及条锈病感病对照品种辉县红,均由西北农林科技大学农学院小麦远缘杂交及染色体工程实验室提供。供试条锈菌生理小种CYR-32和CYR-33混合小种由西北农林科技大学植物保护学院提供。
1.2 试验方法
1.2.1 细胞学鉴定
分别于3月下旬和4月中旬,在适宜的温度条件下,在田间取材料的根尖和幼穗进行细胞学鉴定。根尖和幼穗的固定参照Yang等[15]的方法完成。利用Olympus BX-43显微镜(日本)进行镜检并采集照片,观察并统计其体细胞有丝分裂中期Ⅰ染色体的数目、花粉母细胞减数分裂中期Ⅰ染色体的数目及配对情况、减数分裂后期Ⅰ同源染色体的分离情况。
1.2.2 原位杂交分析
根尖处理:将种子在室温下置于湿润滤纸上,腹沟朝下均匀摆放,置于23℃培养箱萌发2~3 d,待幼根长至2~3 cm时,将根尖分生区切下,采用Wang等[16]的方法处理。选取分裂相良好的染色体采用滴片法制片。
原位杂交分析:用CTAB法提取试验材料幼嫩叶片全基因组DNA。参照Yang等[15]的方法,以中国春的全基因组DNA为封阻,中间偃麦草的全基因组DNA为探针,对ES-24进行基因组原位杂交(genomic in situ hybridization, GISH)鉴定。参照Han等[17]的方法,用中间偃麦草的全基因组DNA以及寡核苷酸探针Oligo-pSc119.2(绿色)和Oligo-pTa535(红色)进行荧光原位杂交(fluorescence in situ hybridization, FISH)鉴定。参照Kruppa等[6]的方法,用Fluorescein-12-dUTP标记百萨偃麦草全基因组DNA,用Texas red-5-dUTP标记拟鹅观草全基因组DNA,以百萨偃麦草全基因组(J)DNA(绿)和拟鹅观草的全基因组(St)DNA(红)为探针,进行mc-GISH鉴定,用Olympus BX-53荧光显微镜观察并用Adobe Photoshop CC2018处理图像。
1.2.3 分子标记分析
利用分布于小麦1~7部分同源群上的90个EST标记和135个PLUG标记,鉴定ES-24所携带的中间偃麦草染色体片段的同源群归属,追踪普通小麦背景中中间偃麦草的遗传物质。EST引物信息参照网站Http://wheat.pw.usda.gov/SNP/new/pcr_primers.shtml,PLUG引物信息参照Ishikawa等[18]发表的引物。引物均由北京奥科鼎盛生物技术有限公司合成。PCR和电泳分析按照Zhu等[19]的方法进行。
1.2.4 条锈病抗性鉴定和农艺性状调查
成株期条锈病抗性鉴定在小麦育种圃进行。鉴定材料于2019年10月中上旬播种,每个材料播种2行,行长1 m,行距0.25 m,株距10 cm,同时种植感病对照辉县红。在2020年3月中旬傍晚,用清水给感病对照辉县红轻喷细雾后,采用撒粉接种法接种条锈病菌生理小种CYR-32和CYR-33的混合小种,然后用薄膜覆盖,翌日清晨去掉薄膜。待辉县红充分发病后,调查发病情况,按照参考文献[20]报道的方法记载反应型,2~3 d后再复查1次。
于2020年6月收获植株后,调查并记录ES-24及亲本的株高、穗长、分蘖数、小穗数、小穗粒数、芒性等性状。
2 结果与分析
2.1 细胞学鉴定结果
根尖分生区体细胞染色体数目观察结果(图1A)显示,ES-24体细胞在有丝分裂中期含有44条染色体。花粉母细胞镜检结果(图1B)显示,减数分裂中期Ⅰ染色体有22对二价体,未观察到多价体的形成,配对情况良好,其构型为2n=44=22Ⅱ。在减数分裂后期Ⅰ,同源染色体均等分离,未检测到滞后染色体(图1C)。以上结果说明,ES-24具有稳定的细胞遗传学机制。初步推测,ES-24携带了一对中间偃麦草的染色体。

A:有丝分裂中期;B:减数分裂中期I;C:减数分裂后期I。A:Mitotic metaphase; B:Meiotic metaphase I; C:Meiotic anaphase I.图1 ES-24的细胞学观察结果Fig.1 Cytological observation of ES-24
2.2 原位杂交鉴定结果
以中间偃麦草的全基因组DNA作为探针,以中国春的全基因组DNA作为封阻,对ES-24根尖染色体进行GISH鉴定,DAPI(蓝色)复染。结果(图2A)显示,ES-24有两条染色体被杂交上绿色信号,而其余42条染色体均呈现为蓝色,表明ES-24是携带42条普通小麦染色体和2条中间偃麦草染色体的二体附加系。
用百萨偃麦草的全基因组DNA和拟鹅观草的全基因组DNA为探针,中国春的全基因组DNA为封阻,进行mc-GISH分析,结果(图2B)显示,ES-24的一对染色体产生了明亮的红色信号。说明ES-24携带了一对中间偃麦草的St染色体。

A:以中间偃麦草全基因组DNA(绿色)为探针与ES-24根尖染色体进行的基因组原位杂交;B:以百萨偃麦草基因组DNA(绿)与拟鹅观草基因组DNA(红)为探针进行的多色GISH;C:以Oligo-pTa535(红)、Oligo-pSc119.2(绿)为探针与ES-24根尖染色体进行的荧光原位杂交。白色箭头所指为中间偃麦草染色体。A:GISH of ES-24 root tip chromosomes using whole genome DNA (green) of Th.intermedium as probe; B:mc-GISH with genomic DNA of Th.bessarabicum(green) and Ps.strigosa(red) as probes; C:FISH of ES-24 root tip chromosomes using Oligo-pTa535 (red) and oligo-pSc119.2 (green) probes.White arrows indicate the chromosomes from Th.intermedium.图2 ES-24的GISH、FISH和mc-GISH原位杂交鉴定Fig.2 GISH,FISH and mc-GISH identifications of ES-24
通过以Oligo-pTa535(红)、Oligo-pSc119.2(绿)为探针对ES-24进行FISH鉴定,ES-24含有42条与中国春FISH核型一致的染色体,一对染色体在其长臂和短臂末端产生Oligo-pTa535红色信号,其信号不同于其他小麦的FISH核型(图2C)。综合来看,ES-24附加了一对中间偃麦草的St染色体。
2.3 分子标记分析结果
利用分布在小麦1~7同源群上的90个EST标记和135个PLUG标记,对ES-24及其亲本(阿勃、远中4和中间偃麦草)进行分子标记分析,最终筛选出3个EST特异标记(BE591737、BG274576和BE637663)和2个PLUG特异标记(TNAC1826和TNAC1821)(表1)。以上标记在ES-24、远中4和中间偃麦草中均扩增出明显的特异性条带(图3),且都位于小麦的第七部分同源群染色体上。结合上述原位杂交结果,表明ES-24携带了一对中间偃麦草的7St染色体。
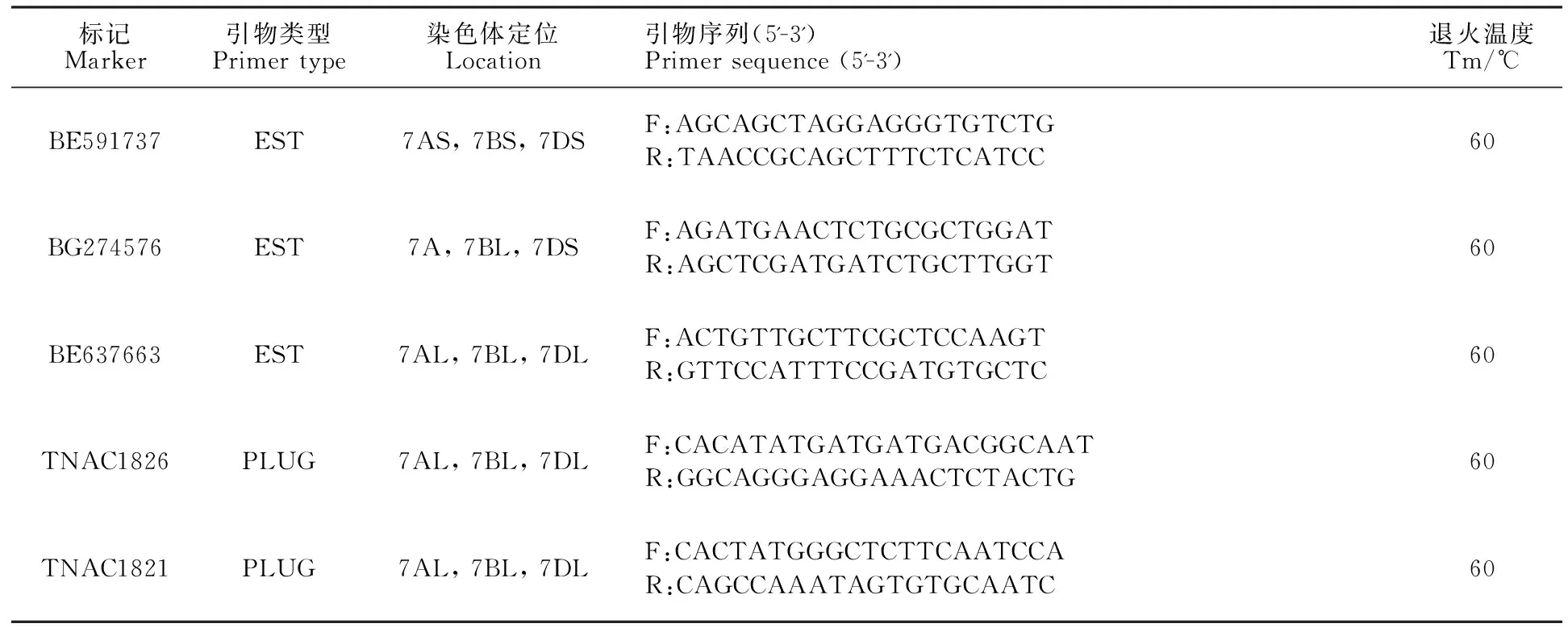
表1 ES-24所携带中间偃麦草7St连锁的EST及PLUG多态性标记Table 1 EST and PLUG polymorphic markers linked to 7St from Th.intermedium carried by ES-24
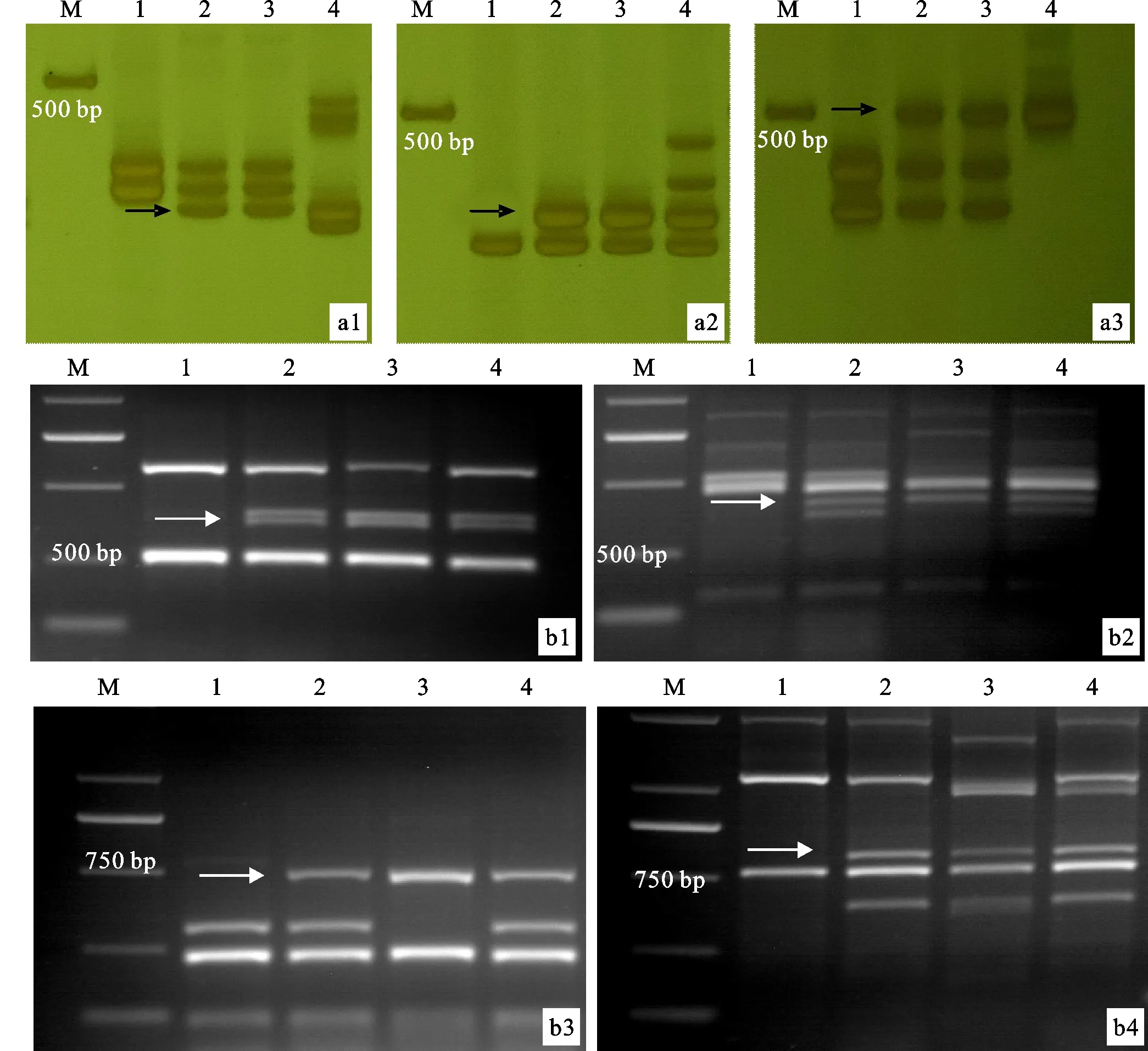
M:DL2000;1:阿勃;2:ES-24;3:远中4;4:中间偃麦草;a1:BE591737;a2:BG274576;a3:BE637663;b1:TNAC1826-Hae III;b2:TNAC1826-Taq I;b3:TNAC1821-Hae III;b4:TNAC1821-Taq I。箭头所指为特异性条带。M:DL2000; 1:Abbondanza; 2:ES-24; 3:Yuanzhong 4; 4:Th.intermedium; a1:BE591737; a2:BG274576; a3:BE637663; b1:TNAC1826-Hae III; b2:TNAC1826-Taq I; b3:TNAC1821-Hae III; b4:TNAC1821-Taq I. Arrows indicate specific bands.图3 用5个多态性标记对ES-24及其亲本的扩增结果Fig.3 Amplification results of ES-24 and its parents using five polymorphic markers
2.4 农艺性状调查和条锈病抗性鉴定结果
从株高、分蘖数、穗长、小穗数、小穗粒数、芒性等农艺性状对ES-24及亲本阿勃和远中4进行成株期综合评价,结果(图4)显示,ES-24的株高、穗长和分蘖数明显低于双亲,小穗数、小穗粒数和芒性与其母本阿勃相近,且其籽粒较为饱满。表明ES-24具有矮秆、籽粒饱满等优良农艺性状,可以作为小麦育种的中间材料。

A:植株;B:穗子;C:小穗;D:籽粒;1:远中4;2:ES-24;3:阿勃。A:Plant; B:Spike; C:Spikelet; D:Grain; 1:Yuanzhong 4; 2; ES-24; 3:Abbondanza.图4 ES-24与其亲本的农艺性状对比Fig.4 Comparison of agronomic traits among ES-24 and its parents
成株期条锈病鉴定结果(图5)显示,感病对照地方品种辉县红表现为高感(IT=4),亲本阿勃表现为中感(IT=3),远中4号和中间偃麦草表现为免疫(IT=0),ES-24表现为高抗(IT=1)。表明ES-24成株期对于条锈菌生理小种CYR32和CYR33混合小种表现为高抗,根据其亲本的抗病性鉴定以及系谱分析,推测ES-24成株期对条锈病的抗性可能来源于其携带的中间偃麦草7St染色体,且中间偃麦草的7St染色体携带有抗条锈病基因。

a:中间偃麦草;b:远中4;c:ES-24;d:阿勃;e:辉县红。a:Th.intermedium ; b:Yuanzhong 4; c:ES-24; d:Abbondanza; e:Huixianhong.图5 ES-24 及其亲本的成株期条锈病抗性表现Fig.5 Stripe rust resistance at adult stage of ES-24 and its parents
3 讨 论
GISH和FISH分析是追踪小麦近缘种属与普通小麦远缘杂交后代外来染色体或片段的准确而强大的工具[21-22]。本研究利用FISH、GISH和mc-GISH,准确而高效地鉴定出ES-24附加了一对中间偃麦草7St染色体。用Oligo-pTa535和Oligo-pSc119.2探针,通过比对ES-24与亲本及中国春标准FISH核型,明确了中间偃麦草7St染色体的FISH核型,为今后中间偃麦草染色体FISH核型的揭示奠定了基础。相比于传统的细胞遗传学技术,分子标记技术则可以更加方便而准确地鉴定出小麦中的外源染色体或片段的同源群归属[23]。本研究通过EST和PLUG分子标记鉴定,发现小麦第七同源群上的5个标记能在ES-24和中间偃麦草中扩增出明显的条带,进一步说明了ES-24的外源染色体为中间偃麦草的7St染色体。
据报道,染色体组之间的交换重组可能与远缘杂交后代染色体组的稳定性和适应性有关[24]。小麦属的亚端粒区域为染色体末端附近的可变区,它们可能通过基因组的不断变化和不同染色体末端之间的均质化来稳定基因组结构。但有关插入外源染色体以及对染色体组的进化机制尚未被证明[22-25]。本研究通过mc-GISH验证,发现ES-24携带一对St染色体,但在St染色体末端的亚端粒区,我们也检测到来自J染色体明亮的绿色信号,表明在进化过程中这对外源染色体与J染色体发生了小片段的易位。此易位可能发生在创制其父本远中4的远缘杂交过程中,也有可能发生在远中4与阿勃杂交的过程中,这需要对其父本的染色体进行全面检测。
条锈病是对小麦最具毁灭性的病害之一。本研究发现,ES-24成株期对于条锈菌生理小种CYR32和CYR33混合小种表现为高抗,推测中间偃麦草的7St染色体携带有抗条锈病基因,但需要进一步的研究验证。
普通小麦与其野生近缘种属远缘杂交所得的二体异附加系,不仅可将所需有益基因转移到普通栽培小麦,而且还可用于研究基因的表达、物种的进化、染色体同源关系、基因定位等,同时也是创制外源染色体代换系和小片段易位系的重要中间材料[26-27]。中间偃麦草作为小麦抗病育种的理想中间资源,前人对其进行了大量研究。Friebe等[28-30]发现,普通小麦-中间偃麦草7Ai#2(7D)和7Ai#2(7A)代换系7Ai#2染色体长臂的末端携带有叶锈病抗性基因Lr38,7Ai#1染色体携带有大麦黄矮病毒抗性基因Bdv2;另外还发现,在7E(J或Js)·7D代换系和7E附加系中携带有大麦黄矮病毒抗性基因Bdv3。Liu等[31]在T7BS·7S#3L易位系(KS12WGGRC59)中发现了一个新的基因wsm3,对小麦条纹花叶病毒具有抗性。本研究发现,ES-24成株期对条锈菌生理小种CYR32和CYR33混合小种表现为高抗,推测中间偃麦草的7St染色体携带有抗条锈病基因。本研究还发现,ES-24具有矮秆、籽粒饱满等优良性状,且其小穗数、穗粒数和芒性都与其亲本阿勃相似。所以ES-24可作为育种中间材料,将中间偃麦草7St染色体上的优良基因导入到普通小麦背景中,但7St染色体上是否具有矮秆基因还需要进一步的验证。由于异附加系的局限性,通过60Coγ辐射,能快速、有效地引起染色体之间的变异[32],进而创制染色体的代换系或易位系。因此,利用以上方法对ES-24进行处理,可以创制高抗条锈病的代换系或小片段易位系材料,并通过连续的回交和自交,可获得高抗条锈病等优良性状的新种质。
