抗生素短期干预所致肠道菌群紊乱对大鼠食欲和糖脂代谢的影响
2021-10-09刘秀吕秋兰赵鹏杨晓敏徐大星邢士超
刘秀 吕秋兰 赵鹏 杨晓敏 徐大星 邢士超
[摘要] 目的 探讨抗生素短期干预所致肠道菌群紊乱对大鼠食欲和糖脂代谢的影响。方法 联合使用氨苄西林、新霉素、甲硝唑干预大鼠的肠道菌群,利用高通量测序技术,分析抗生素对大鼠肠道菌群的干预情况。通过连续监测大鼠的进食和进水量、尿量、体质量、血糖和血脂,分析肠道菌群被干预大鼠的食欲及糖脂代谢变化。结果 抗生素干预了大鼠的肠道菌群,而肠道菌群紊乱大鼠的进食和进水量、尿量显著增高(t=3.95~7.54,P<0.05),但其体质量、血糖和血脂无显著变化(t=0.05~1.89,P>0.05)。结论 抗生素短期干预所致肠道菌群紊乱影响了大鼠的食欲,而大鼠的体质量、糖脂代谢未受影响。
[关键词] 胃肠道微生物组;抗菌药;食欲;碳水化合物代谢;脂类代谢;大鼠
[中图分类号] R378.2
[文献标志码] A
[文章编号] 2096-5532(2021)04-0498-05
近年来,越来越多的人出现肥胖和饮食失调症状[1-2],这无疑反映了人们对食欲紊乱的原因认识不足以及对其缺乏有效的治疗[3-4],因此,探究影响宿主食欲调节的因素,是治疗肥胖、厌食症、暴食症和恶病质等食欲紊乱性疾病的重中之重[5-8]。众所周知,人体肠道寄居着数万亿的微生物[9],而这些微生物可以通过与宿主的相互作用和共同的代谢活动影响宿主的生理功能和病理生理,因此肠道微生物在人类的健康和疾病中扮演着重要角色[10-11]。有研究显示,肥胖、厌食症和暴食症病人的肠道微生物群发生改变,提示肠道菌群与宿主的食欲调节及代谢之间有潜在联系[12-14]。本研究通过联合使用抗生素干预大鼠的肠道菌群,进而观察肠道菌群紊乱大鼠的食欲变化以及糖脂代谢情况,探讨肠道菌群对宿主的食欲调节和代谢的影响,以推动针对食欲紊乱性疾病的新型诊断和治疗方法的出现。
1 材料与方法
1.1 材料
雄性6周龄Wistar大鼠20只,购自青岛琴大生物科技有限公司,饲养于青岛大学附属医院SPF级动物房内。动物实验经青岛大学附属医院动物研究伦理委员会批准(AHQU-MAL2017022)。氨苄西林、青霉素、甲硝唑购自生工生物工程(上海)股份有限公司。E.Z.N.A Soil DNA试剂盒(OMEGA),AxyPrep DNA凝胶回收试剂盒(AXYGEN),ABI GeneAmp 9700 型PCR仪,德国罗氏卓越血糖仪,全自动生化分析仪(TOSHIBA,Japan),大鼠代谢笼,大鼠灌胃针。
1.2 动物分组和处理
适应性喂养1周后,将20只大鼠随机分为正常组(C组)和抗生素组(A组),每组10只。参照文献方法并适当改进[15],A组大鼠喂以正常饲料,并称取一定剂量的抗生素(氨苄西林250 mg/kg、新霉素250 mg/kg、甲硝唑50 mg/kg),用无菌水溶解(混合溶液浓度为50 g/L)后每天灌胃1次, 连续灌胃5周;C组大鼠喂以正常饲料,每天以等体积的无菌水灌胃1次,连续灌胃5周。每周六记录1次大鼠体质量,每周日记录1次大鼠的24 h进食量、进水量以及尿量,每周一检测1次大鼠的血糖和血脂。
1.3 大鼠进食量、进水量、尿量检测
每个大鼠代谢笼饲养1只大鼠,将相同质量的大鼠饲料、相同体积的无菌水以及干净的尿杯分配给每只大鼠,24 h后,再次称量每只大鼠的剩余饲料,用量筒或1 000 mL的移液枪分别量取每只大鼠的进水量和尿量,以此分析大鼠的食欲情况。
1.4 大鼠粪便样本的收集
在抗生素使用前(0周)和抗生素使用3周后(3周)分别收集大鼠的新鲜粪便。用碘附擦拭大鼠肛门后,将大鼠放于无菌报纸上,用手按摩大鼠屁股,以无菌镊子夹取大鼠肛门处新鲜粪便置于无菌冷冻管中,并立即暂存于液氮罐中,之后转移至-80 ℃冰箱以供肠道菌群测序。
1.5 454焦磷酸测序
1.5.1 粪便DNA提取和PCR扩增 每只大鼠取50~100 mg粪便,使用E.Z.N.A Soil DNA试剂盒抽提粪便的基因组DNA,之后应用10 g/L的琼脂糖凝胶电泳检测抽提的DNA;利用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和533R(5′-TTACCGCGGCTGCTGGCAC-3′)在ABI GeneAmp 9700型PCR仪中扩增16S rDNA的V1~V3高变区,扩增产物用20 g/L的瓊脂糖凝胶电泳,使用AxyPrep DNA凝胶回收试剂盒切胶回收PCR产物。
1.5.2 测序及生物信息学分析 回收的PCR产物用罗氏GS-FLX 454焦磷酸测序系统进行测序,为保证序列的质量,分析前先对原始数据进行过滤处理,得到优化序列。排除标准如下:①引物的错配数大于2的序列;②长度短于250 bp、模糊碱基数大于0、序列平均质量低于 25的序列;③未测通目标区域的序列。其次,将优化的序列按照97%的相似性分归为许多个操作分类单元(OTU),进行生物信息统计分析。将每条优化序列均与SILVA(SSU111版)的小亚基核糖体RNA(SSUrRNA)数据库进行比对,找出其最相近且可信度达80%以上的种属信息。为获得每个OTU的分类学信息,将97%相似水平下每个OTU中的所有序列进行一致性分析,找出同一个OTU中不同序列的最近祖先的种属信息作为该OTU的种属信息。
1.6 血清生化指标的检测
采集静脉血之前,大鼠禁食8~10 h。于每周一早晨,采用剪尾取血法取血,用血糖仪检测大鼠的空腹血糖(BG)浓度;采用内眦取血法采集大鼠的空腹静脉血,以3 000 r/min离心10 min,吸取血清,使用全自动生化分析仪检测血清中三酰甘油(TG)和胆固醇(CHOL)的浓度。
1.7 统计学分析
应用GraphPad Prism软件绘图,SPSS 17.0软件进行统计学处理。计量资料结果以X2±s表示,数据间的比较采用析因设计的方差分析,两组间比较采用t检验,以P<0.05为差异有统计学意义。
2 结 果
2.1 抗生素对大鼠肠道菌群的影响
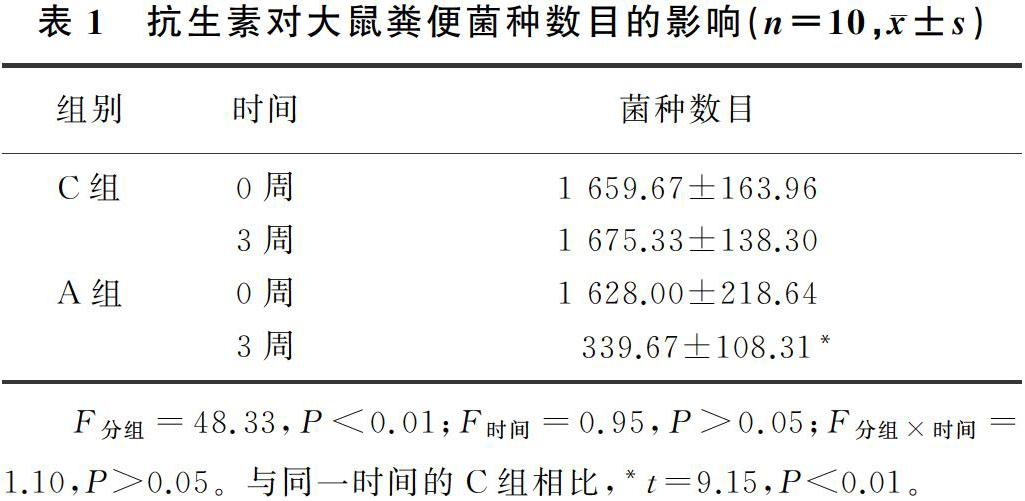
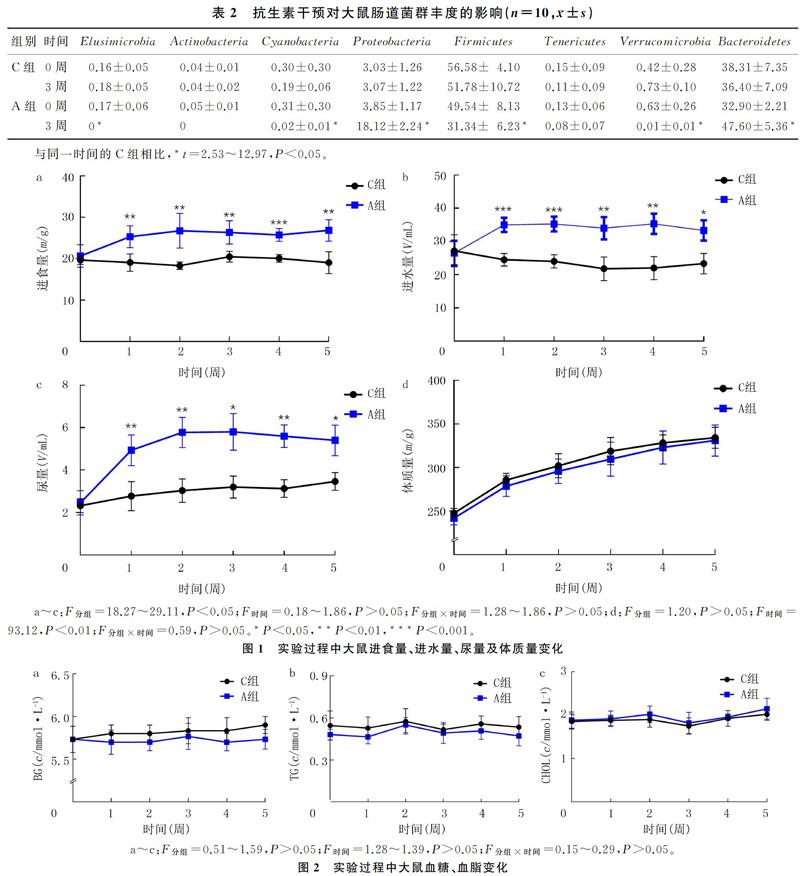
抗生素的使用显著降低了大鼠的肠道菌种数目(t=9.15,P<0.01)。见表1。在门水平分析大鼠的肠道菌群组成,结果显示,抗生素干预了大部分的肠道菌门,即Elusimicrobia、Cyanobacteria、Proteobacteria、Firmicutes、Verrucomicrobia 和Bacteroidetes的相对丰度都发生显著变化(t=2.53~12.97,P<0.05)。见表2。大鼠粪便菌种数目的急剧降低以及肠道菌群相对丰度的显著改变表明,抗生素使大鼠的肠道菌群受到严重干预。
2.2 肠道菌群紊乱大鼠的食欲变化
在整个实验过程中,C组大鼠的进食量、进水量和尿量始终保持稳定。而在抗生素处理1周后,A组大鼠的进食量、进水量和尿量开始显著升高,并一直维持至第5周。抗生素处理后1~5周,A组大鼠的进食量、进水量和尿量始终高于C组(t=3.95~7.54,P<0.05)。提示抗生素短期干预所致的肠道菌群紊乱影响了大鼠的食欲,肠道菌群在大鼠的食欲调节中起着潜在作用。见图1。
2.3 肠道菌群紊乱大鼠的体质量变化
C组和A组大鼠的体质量均呈现出增高趋势,在整个实验过程中,两组大鼠的体质量差异无统计学意义(t=0.36~1.68,P>0.05)。见图1。
2.4 肠道菌群紊乱大鼠的糖脂代谢变化
C组和A组大鼠的血糖、血脂始终同向波动,并且在整个实验过程中,两组大鼠的血糖、血脂差异均无显著性(t=0.05~1.89,P>0.05)。见图2。
3 讨 论
健康的肠道微生物群对宿主正常的代谢过程和微生态稳态至关重要,而肠道微生物组成的改变,可能通过影响宿主的新陈代谢而导致肥胖、糖尿病等[16-17]。过度使用抗生素,可以对肠道微生物群产生重大影响,导致其组成和功能的改变[18-20],而微生物组成和功能的改变,又可能诱发各种内分泌、代谢性、免疫性疾病[21-23]。本研究参考相关文献的实验方法并加以适当改进[15],联合使用氨苄西林、新霉素和甲硝唑干预大鼠的肠道菌群,通过对大鼠进食量、进水量、尿量、体质量以及血糖、血脂的监测,来分析肠道菌群紊乱大鼠的进食和代谢情况,进而探讨肠道菌群对宿主食欲和代谢的影响。
本研究结果显示,肠道菌群紊乱大鼠的进食量和进水量发生了显著变化,说明肠道菌群可能在宿主的食欲调节中发挥着重要作用。有研究表明,肠道菌群的改变可以导致炎性细胞因子的增加,这些细胞因子又会通过致炎症作用参与代谢疾病(如肥胖和2型糖尿病)和中枢神经系统相关的精神性疾病(如认知障碍、焦虑和抑郁)的发生[17,24],而包括焦虑和抑郁等在内的精神性疾病明显影响着宿主的食欲[25]。因此,肠道菌群或许通过炎性细胞因子或其代谢产物参与调节宿主的食欲,而进一步筛选出与食欲调节有关的肠道菌种,将推动食欲紊乱性疾病的新型诊断与治疗。另外有研究表明,尽管没有任何肠道菌群的无菌小鼠的进食量大于正常小鼠,但其体质量并不会比正常小鼠更大[26]。同样,本研究结果显示,食欲提高大鼠的体质量与正常大鼠相比并无显著变化,这意味着肠道菌群在宿主的能量平衡中起着重要作用。
近年来,大量的研究表明,肠道菌群与宿主糖尿病和脂代谢紊乱性疾病之间有着密切联系[26-27]。然而,本研究并没有观察到肠道菌群紊乱大鼠的异常的血糖和血脂浓度,但这并不能说明肠道菌群与糖脂代谢没有关系,因为有研究发现,紊乱的肠道菌群不能改变正常饮食大鼠的脂代谢,却可以加剧高胆固醇饮食大鼠的血脂积累[15]。这些结果提示,紊乱的肠道菌群尚不能作为单独的影响因素驱动正常宿主的代谢变化。因此,有效的饮食控制和保持肠道微生物的稳态将有望用来预防和治疗糖脂代谢紊乱性疾病。
然而,本次研究仅对大鼠的肠道菌群进行了短期干预,进而对大鼠的食欲、体质量和糖脂代谢情况连续监测了5周,其结果仅代表抗生素短期干预所致肠道菌群紊乱对大鼠食欲和糖脂代谢的影响。在接下来的研究中,我们计划将抗生素组大鼠的粪便菌群移植给无菌大鼠,进而观测无菌大鼠的食欲、糖脂代谢变化或者对比分析两组大鼠的肠道菌群代谢产物[28-30],然后将变化的菌群代谢产物与食欲调节、糖脂代谢相关的信号通路联系起来[31-32],以此驗证肠道菌群在食欲变化、糖脂代谢中的潜在作用。
综上所述,本研究利用抗生素干预了大鼠的肠道菌群,发现肠道菌群紊乱大鼠的进食量、进水量和尿量显著增高,表明肠道菌群可能参与调节宿主食欲,但对正常大鼠的糖脂代谢没有明显影响。这些结果为开发肥胖、厌食症、暴食症和恶病质等食欲紊乱性疾病的新型诊断和治疗方法带来了希望。
[参考文献]
[1]CANNING P M. Prevalence of overweight and obesity in a provincial population of Canadian preschool children[J]. Canadian Medical Association Journal, 2004,171(3):240-242.
[2]XIAO Y Y, QIAO Y J, PAN L, et al. Trends in the prevalence of overweight and obesity among Chinese preschool children from 2006 to 2014[J]. PLoS One, 2015,10(8):e0134466.
[3]KUMANYIKA S. Overcoming inequities in obesity: what dont we know that we need to know[J]? Health Education & Behavior, 2019,46(5):721-727.
[4]AIGNER M, TREASURE J, KAYE W, et al. World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the pharmacological treatment of eating disorders[J]. The World Journal of Biological Psychiatry, 2011,12(6):400-443.
[5]SMITH S, WOODSIDE D. Characterizing treatment-resistant anorexia nervosa[J]. Frontiers in Psychiatry, 2020,11:542206.
[6]LANTZOUNI E, GRADY R. Eating disorders in children and adolescents: a practical review and update for pediatric gynecologists[J]. Journal of Pediatric and Adolescent Gynecology, 2021. doi:10.1016/j.jpag.2021.01.010.
[7]HAGAN K, WALSH B. State of the art: The therapeutic approaches to bulimia nervosa[J]. Clinical Therapeutics, 2020. doi:10.1016/j.clinthera.2020.10.012.
[8]REAS D, GRILO C. Psychotherapy and medications for eating disorders: Better together[J]? Clinical Therapeutics, 2020. doi:10.1016/j.clinthera.2020.10.006.
[9]OTTMAN N, SMIDT H, DE VOS W M, et al. The function of our microbiota: who is out there and what do they do[J]? Frontiers in Cellular and Infection Microbiology, 2012,2:104.
[10]HILLS R D, PONTEFRACT B A, MISHCON H R, et al.Gut microbiome: profound implications for diet and disease[J]. Nutrients, 2019,11(7):1613.
[11]PINDJAKOVA J, SARTINI C, LO RE O, et al. Gut dysbiosis and adaptive immune response in diet-induced obesity vs. systemic inflammation[J]. Frontiers in Microbiology, 2017,8:1157.
[12]TURNBAUGH P J, LEY R E, MAHOWALD M A, et al. An obesity-associated gut microbiome with increased capacity for energy harvest[J]. Nature, 2006,444(7122):1027-1031.
[13]ARMOUGOM F, HENRY M, VIALETTES B, et al. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients[J]. PloS one, 2009,4(9):e7125.
[14]HIMMERICH H, BENTLEY J, KAN C, et al. Genetic risk factors for eating disorders: an update and insights into pathophysiology[J]. Therapeutic Advances in Psychopharmacology, 2019,9:1-20.
[15]HU X, WANG T, LIANG S, et al. Antibiotic-induced imba-lances in gut microbiota aggravates cholesterol accumulation and liver injuries in rats fed a high-cholesterol diet[J]. Applied Microbiology and Biotechnology, 2015,99(21):9111-9122.
[16]COX L M, YAMANISHI S, SOHN J, et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences[J]. Cell, 2014,158(4):705-721.
[17]PAIVA I H R, DUARTE-SILVA E, PEIXOTO C A. The role of prebiotics in cognition, anxiety, and depression[J]. European Neuropsychopharmacology: the Journal of the European College of Neuropsychopharmacology, 2020,34:1-18.
[18]姚增武,仲蓓,王东升,等. 广谱抗生素对大鼠肠道菌群多样性影响的16S rDNA基因测序研究[J]. 齐鲁医学杂志, 2014,29(1):32-34.
[19]ZHAO L, HUANG Y F, LU L, et al. Saturated long-chain fatty acid-producing bacteria contribute to enhanced colonic motility in rats[J]. Microbiome, 2018,6(1):107.
[20]王虑,刘绍泽,王胜云,等. 广谱抗生素对肠道菌群紊乱诱导的侵袭性真菌感染的影响[J]. 创伤外科杂志, 2010,12(4):24-27.
[21]CRUSELL M K W, HANSEN T H, NIELSEN T, et al. Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum[J]. Microbiome, 2018,6(1):89.
[22]BEHROUZI A, ASHRAFIAN F, MAZAHERI H, et al. MiRNAs in outer membrane vesicles of gut microbiota: Friend or foe[J]? Microbial Pathogenesis, 2020. doi:10.1016/j.micpath.2020.104200.
[23]XU M Q, CEN M S, SHEN Y Q, et al. Deoxycholic acid-induced gut dysbiosis disrupts bile acid enterohepatic circulation and promotes intestinal inflammation[J]. Digestive Diseases and Sciences, 2020. doi:10.1007/s10620-020-06208-3.
[24]DINAN T G, CRYAN J F. Mood by microbe: towards clinical translation[J]. Genome Medicine, 2016,8(1):36.
[25]SCHELLEKENS H, FINGER B C, DINAN T G, et al. Ghrelin signalling and obesity: at the interface of stress, mood and food reward[J]. Pharmacology & Therapeutics, 2012,135(3):316-326.
[26]BACKHED F, DING H, WANG T, et al. The gut microbiota as an environmental factor that regulates fat storage[J]. PNAS, 2004,101(44):15718-15723.
[27]LUCA M, DI MAURO M, DI MAURO M, et al. Gut microbiota in Alzheimers disease, depression, and type 2 diabetes mellitus: the role of oxidative stress[J]. Oxidative Medicine and Cellular Longevity, 2019(7,article e138):1-10.
[28]YANO J, YU K, DONALDSON G, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis[J]. Cell, 2015,161(2):264-276.
[29]闫瑶瑶. 高尿酸血症相关肠道菌群研究[D]. 青岛: 中国海洋大学, 2015.
[30]XI Y, YAN J, LI M, et al. Gut microbiota dysbiosis increases the risk of visceral gout in goslings through translocation of gut-derived lipopolysaccharide[J]. Poultry Science, 2019,98(11): 5361-5373.
[31]ZHAO J, ZHANG X, LIU H, et al. Dietary protein and gut microbiota composition and function[J]. Current Protein & Peptide Science, 2018,20(2):145-154.
[32]ZHANG L, DAVIES S. Microbial metabolism of dietary components to bioactive metabolites: opportunities for new therapeutic interventions[J]. Genome Medicine, 2016,8(1):46.
(本文編辑 马伟平)
