抗菌肽HBβ-C在全雄和杂交黄颡鱼对多子小瓜虫抗性差异中的作用
2021-09-08张义兵
熊 阳 王 帅 丹 成 张义兵 梅 洁
(1. 华中农业大学水产学院, 武汉 430070; 2. 中国科学院水生生物研究所, 武汉 430072)
黄颡鱼(Pelteobagrus fulvidraco)是我国一种重要的淡水经济鱼类, 肉质鲜美, 无肌间刺, 营养价值高, 深受广大消费者的喜爱。在相同的养殖条件下,雄性黄颡鱼在养殖第二年的生长速度是雌性的1.5—2倍, 培育全雄黄颡鱼能够显著提高养殖产量和经济效益[1]。中国科学院水生生物研究所桂建芳课题组和华中农业大学梅洁课题组开发出了性染色体特异的性别标记, 并建立了一条X和Y染色体连锁标记辅助的全雄鱼培育技术路线[2—6], 推动黄颡鱼产业的迅速发展。近年来, 由于YY超雄鱼多代自交退化及在亲本培育过程中遇到一些问题, 制约了全雄黄颡鱼产业的发展[7], 需要进行品种改良或培育新品种。杂交黄颡鱼(黄颡鱼Pelteobagrus fulvidraco♀×瓦氏黄颡鱼Pelteobagrus vachelli♂)形态特征与黄颡鱼相近, 且其生长速度明显优于黄颡鱼。华中农业大学等多家单位和企业进行联合育种研究, 采用传统选育和杂交育种的方法, 培育出了水产新品种杂交黄颡鱼“黄优1号”(GS-02-001-2018)[8]。育种技术的创新和新品种的培育推动了黄颡鱼产业的持续发展, 2019年我国黄颡鱼养殖产量高达5.37×108kg, 成为发展速度最快的特色淡水鱼之一[9]。
近年来, 由多子小瓜虫(Ichthyophthirius multifiliis)引起的小瓜虫病(又称白点病)成为黄颡鱼养殖常见的疾病之一, 给我国黄颡鱼养殖业造成巨大的经济损失。多子小瓜虫没有严格的宿主专一性,能感染绝大部分淡水鱼类, 如斑点叉尾鮰(Ictalurus punctatus)、虹鳟(Oncorhynchus mykiss)、泥鳅(Misgurnus anguillicaudatus)、鲤(Cyprinus carpio)、尼罗罗非鱼(Oreochromis niloticus)、赤眼鳟(Squaliobarbus curriculus)和鳙(Hypophthalmichthys nobilis)等经济鱼类均易受到感染[10—16]。刺激隐核虫(Cryptocaryon irritans; 又称为海水小瓜虫)引起的白点病已成为大范围、经常暴发的传染性疾病, 给海水鱼养殖也造成巨大的经济损失, 大部分海水养殖鱼类如斜带石斑鱼(Epinephelus coioides)、卵圆鲳鲹(Trachinotus blochii)和大黄鱼(Pseudosciaena crocea)等均容易感染刺激隐核虫[17—19]。小瓜虫病的防治目前仍是国际性的难题。
鱼类的皮肤黏液作为抵御寄生虫感染的第一道重要屏障, 含有诸多抑杀病原的活性物质如免疫球蛋白、转铁蛋白、补体、凝集素和抗菌肽等[20]。黄斑蓝子鱼(Siganus oramin)等极少数鱼类具有天然抗刺激隐核虫(Cryptocaryon irritans)的能力[21],其皮肤黏液蛋白对多子小瓜虫和刺激隐核虫均有杀伤效果[22]。鱼类的鳃和皮肤作为分泌皮肤黏液的主要器官, 有学者在其中发现了一个抗菌肽(位于血红蛋白β链HBβ C端的肽段)具有抗多子小瓜虫的效果[23]。最近, 通过蛋白质组技术发现HBβ存在于黄颡鱼皮肤黏液中, 其表达量在鲇爱德华氏菌感染后明显上升[24], 说明HBβ可能具有抗菌功能。据多数养殖户反映, 全雄黄颡鱼和杂交黄颡鱼对多子小瓜虫的敏感程度存在明显差异。为解析全雄黄颡鱼与杂交黄颡鱼对多子小瓜虫抗性差异的分子机制, 本研究系统地比较了全雄黄颡鱼、杂交黄颡鱼和瓦式黄颡鱼对多子小瓜虫的抗性差异, 检测了HBβ基因在全雄黄颡鱼和杂交黄颡鱼免疫相关组织中的表达差异, 研究了人工合成黄颡鱼抗菌肽(HBβ C端的肽段, HBβ-C)体外的杀虫效果, 旨在为黄颡鱼多子小瓜虫病的防控提供技术保障。
1 材料与方法
1.1 实验鱼
全雄黄颡鱼(4.43±0.45) g、杂交黄颡鱼(6.07±0.64) g和瓦式黄颡鱼(8.65±1.18) g在武汉百瑞生物技术有限公司循环水养殖系统暂养2周。在暂养期间, 随机选取全雄黄颡鱼、杂交黄颡鱼和瓦式黄颡鱼各10尾在光学显微镜下对鳃、鳍条和皮肤黏液进行仔细检查, 均未见小瓜虫等寄生虫。
1.2 小瓜虫的培养及其滋养体、包囊体和掠食体的分离
采用张其中等[25]报道的方法分离小瓜虫的滋养体、包囊体和掠食体, 用于体外攻毒试验。将已感染小瓜虫的赤眼鳟与杂交黄颡鱼饲养于同一循环水箱中进行平行感染维持实验所需的小瓜虫。首先用200 mg/L MS-222 (甲基磺酸三胺)将严重感染的杂交黄颡鱼麻醉, 然后用干净的细胞刮刀从鱼皮肤和鳍条上取出包裹着囊泡的滋养体, 反复清洗多次后, 直至去除大部分黏液, 将干净滋养体放置装有养殖系统水的培育皿中用于体外抗虫试验和形成包囊体。滋养体在20℃下约10min停止游动吸附在培育皿底部, 形成一层透明的薄膜形成包囊体,虫体在内旋转运动并以二分裂的形式分裂形成子体, 约20h后成熟并释放出400—600个具有感染性的掠食体, 收集后用于抗虫试验。
1.3 小瓜虫人工感染试验及样品收集
在暂养2周后, 将健康的全雄黄颡鱼、瓦式黄颡鱼和杂交黄颡鱼分别随机分为对照组和感染组置于90 L水箱中, 水温保持在(20±1.0)℃, 溶解氧≥6 mg/L以上。每组3个平行, 每个平行随机放入65尾鱼。此外, 添加一组混合感染组即全雄黄颡鱼和杂交黄颡鱼各32尾, 通过分别剪左右腹鳍进行物理辨别。参照Clark等[26]方法进行小瓜虫人工感染实验, 3尾已被平行感染的杂交黄颡鱼与每个感染组共栖息在同一个养殖缸中, 而对照组则不放入受感染的鱼, 其他养殖环境相同。在试验期间, 及时清理死鱼以免污染水质, 记录其感染情况并统计每个组合死亡率。在感染的第5天, 每个组合分别从3个平行鱼箱中各取5尾鱼, 用200 mg/L MS-222 (甲基磺酸三胺)麻醉后, 分别取部分皮肤和鳃组织固定在4%多聚甲醛24h, 用于免疫组化和病理学分析;而另取部分组织用液氮处理后转移置-80℃冰箱保存, 用于血红蛋白β链基因的组织表达分析。
1.4 黄颡鱼血红蛋白源抗菌肽HBβ-C生物信息学分析及化学合成
前期我们通过蛋白质组技术比较鲇爱德华氏菌感染前后黄颡鱼皮肤黏液蛋白质的表达变化[24]。本实验通过本地blastall程序将黄颡鱼黏液蛋白质组所有肽段序列与抗菌肽数据库(http://aps.unmc.edu/AP/APD3_update2020_release.fasta)进行比较分析确定抗菌肽位置。使用以下软件和工具对血红蛋白源抗菌肽HBβ-C进行分析: Clone Manager软件对黄颡鱼血红蛋白β链基因序列进行分析, 确定其开放阅读框(ORF); DNAMAN 7.0软件进行氨基酸序列分析; BLAST工具(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行核苷酸和氨基酸序列相似性分析。利用HeliQuest工具(https://heliquest.ipmc.cnrs.fr)进行抗菌肽HBβ-C进行氨基酸电荷和疏水性分析。以硬骨鱼Leiostomus xanthurus的血红蛋白β链(蛋白数据检索号1spg.1.B)为模板使用SWISS-MODEL server (http://swissmodel. expasy.org/)进行建模预测抗菌肽结构。
抗菌肽HBβ-C由南京市金斯瑞生物科技股份有限公司采用固相化学合成法合成, 反向液相色谱确定纯度在90%以上, 然后利用质谱确定分子量准确无误。
1.5 血红蛋白β链在黄颡鱼皮肤和鳃组织中表达量分析
用TRIzol试剂(中国大连, TaKaRa)提取每个样品的总RNA; 通过NanoDrop 2000测定RNA浓度和琼脂糖凝胶电泳确认提取的RNA完整后, 等量混合;使用PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa)试剂盒按说明书要求进行反转录得到cDNA。RT-PCR反应体系: SYBR Green Mixture 10 μL,10 μmol/L的正向引物(F: 5′-TTCACGCTTCTTGC TAACT-3′)和反向引物(R: 5′-CAGCCACGACGAC ATTCA-3′)各0.5 μL, 去离子水8.0 μL, cDNA 1.0 μL。反应条件: 95℃预变性30s, 40个循环(95℃ 5s, 60℃30s), 溶解曲线: 65—95℃、0.5℃/次, 5s读板。每个样品设置3个重复。反应结束后分析扩增产物的溶解曲线, 以黄颡鱼β-actin作为内参基因, 分析每个样品中基因表达情况。
Western blot印迹分析: 将黄颡鱼HBβ-C肽段部分序列(PEVHETWQKFLN)进行化学合成后注射到新西兰大白兔获得兔源的HBβ多克隆抗体; 提取鳃组织的总蛋白后再进行15% SDS-PAGE电泳, 用湿转法进行转膜; 在封闭液(5%脱脂奶粉)中封闭1h, 封闭液加入HBβ多克隆抗体(1﹕1000)中在摇床上4℃封闭过夜; 洗膜, 加入HRP-conjugated Goat Anti-Rabbit lgG(H+L) (1﹕1000), 室温摇床孵育1h;洗膜, 用ECL显色系统进行显色, 置红外激光成像系统检测并拍照。Western blot结果中的条带强度使用Image J软件分析。
1.6 HBβ-C体外抗小瓜虫试验
将化学合成的HBβ-C粉剂溶于灭菌双蒸水中使其终浓度为1000 μg/mL, 用双蒸水稀释为500、250、125、50、25、15、5、2和1 μg/mL。体外试验在96孔细胞培养板中进行, 每孔加入70 μL不同浓度抗菌肽和10个滋养体, 每种浓度3个平行孔。试验设置未加抗菌肽的作为阴性对照组。每0.5h镜检1次, 4h后统计各孔死亡情况。包囊体和掠食体的药物处理同上。不同生活史小瓜虫死亡判断标准为虫体纤毛不再运动, 膜破裂, 胞内物质外流。
1.7 组织病理学分析
组织样品做石蜡包埋后进行连续切片, 分别进行苏木精-伊红(HE)染色和免疫组化分析。
免疫组化分析: 组织石蜡切片置于65℃烘箱中烘片2h进行脱蜡, PBS洗涤(洗涤3次, 每次5min, 下同)后置于EDTA缓冲液中微波修复, 自然冷却后PBS洗涤; 切片置于3%过氧化氢溶液中室温下避光孵育10min, 阻断内原酶反应; PBS洗涤, 甩干后用5% BSA封闭20min, 封闭非特异性反应; 去除BSA液, 每张切片加入HBβ多克隆抗体(1﹕200)覆盖组织,4℃过夜; PBS洗涤, 每张切片加入Cy3-conjugated Goat Anti-Rabbit lgG(H+L) (1﹕1000), 37℃孵育50min; PBS洗涤, 每张切片加50—100 μL DAPI染液, 室温避光孵育5min; 染色后将切片放入PBS洗涤, 滴加适量的抗荧光淬灭剂于组织上, 盖玻片封片, 荧光显微镜下观察。
1.8 数据分析
使用Excel 2010和SPSS 11.0软件统计分析数据, 采用单因素方差分析(One-Way ANOVA)检验小瓜虫感染组与对照组样品间血红蛋β链基因表达量差异。实验数据为平均值±标准差(SD)的方式表示,P<0.05表示差异性显著(*),P<0.01表示差异性极显著(**)。利用GraphPad Prism 6.0软件做基因表达和Kaplan-Meier生存曲线图, 用Mantel-Cox检验分析曲线之间的差异。
2 结果
2.1 全雄黄颡鱼、杂交黄颡鱼和瓦氏黄颡鱼对多子小瓜虫的抗性差异分析
为了评估全雄黄颡鱼、杂交黄颡鱼和瓦式黄颡鱼对多子小瓜虫的敏感性, 我们进行了感染试验。在3种鱼进行感染实验后, 每日死亡状况如表 1所示。杂交黄颡鱼和瓦式黄颡鱼极易感染多子小瓜虫, 在感染5—6d即可观察到清晰的小白点寄生在皮肤和鳍条上, 光学显微镜下也可观察鳃部被感染, 感染率高达100%。在试验期间, 全雄黄颡鱼未发现临床症状, 通过光学显微镜观察未检测到多子小瓜虫存在于全雄黄颡鱼鳃、皮肤和鳍条组织。在相同感染条件下, 瓦式黄颡鱼和杂交黄颡鱼的死亡率高达100%显著高于全雄黄颡鱼(P<0.0001), 而所有对照组鱼均存活且未被感染(图 1)。在混合感染组中, 杂交黄颡鱼因感染多子小瓜虫大批量死亡,而全雄黄颡鱼依旧镜检未检测到多子小瓜虫存在(图 2), 少量几条全雄黄颡鱼可能由于鳍条破损细菌感染而死亡。

图1 全雄黄颡鱼、杂交黄颡鱼和瓦式黄颡鱼感染多子小瓜虫的存活曲线图Fig. 1 The survival curves of all-male yellow catfish, hybrid yellow catfish and darkbarbel catfish after infected with Ichthyophthirius multifiliis
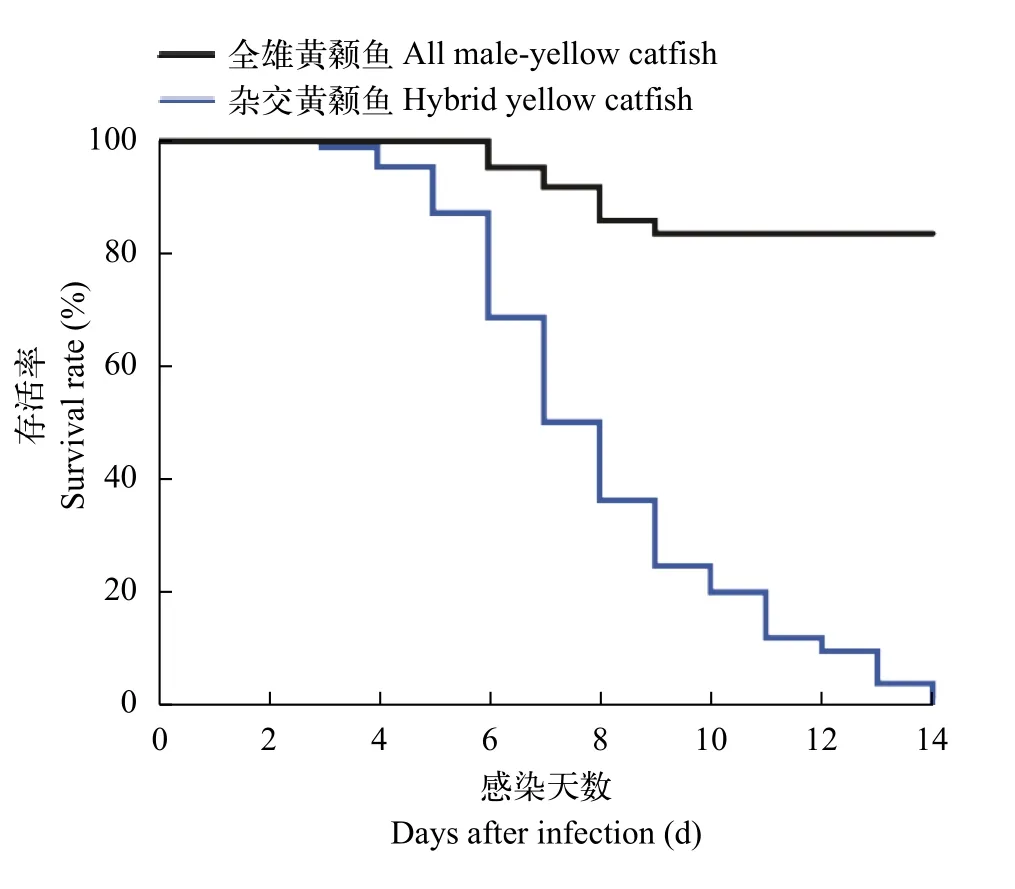
图2 全雄黄颡鱼和杂交黄颡鱼混合感染多子小瓜虫的存活曲线图Fig. 2 The survival curves of all-male yellow catfish and hybrid yellow catfish after infected with Ichthyophthirius multifiliis. Allmale yellow catfish and hybrid yellow catfish were mixed together
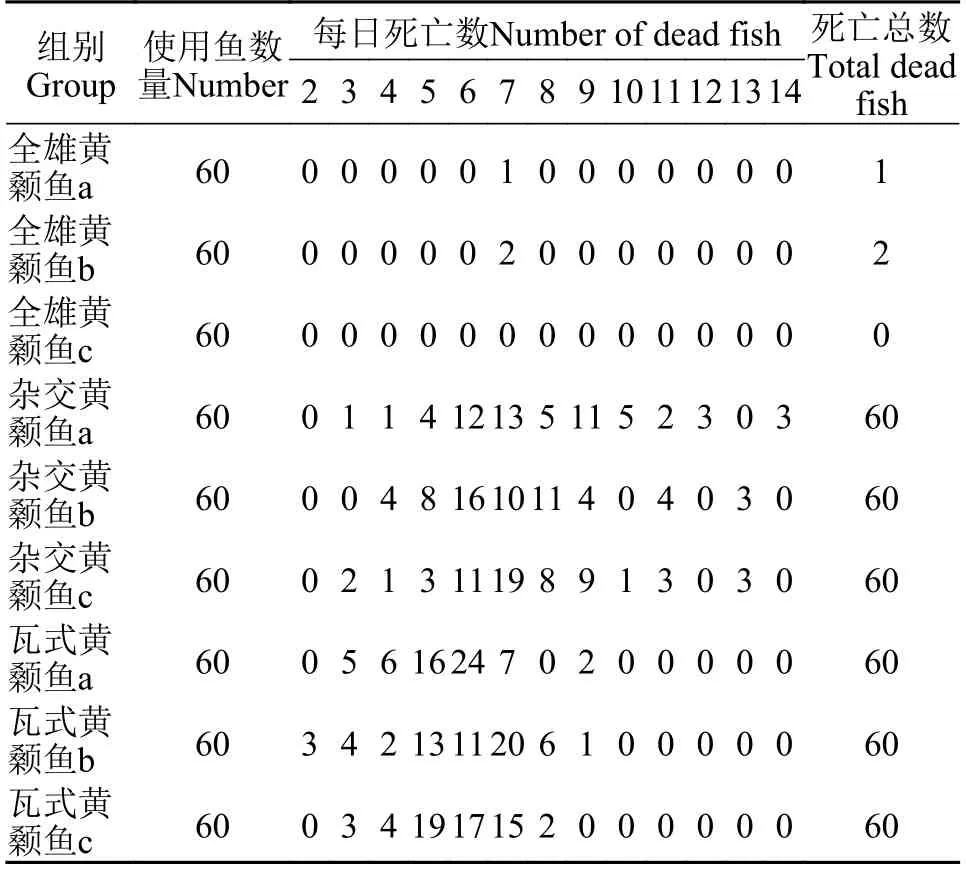
表1 黄颡鱼感染多子小瓜虫每日死亡状况(尾)Tab. 1 The daily dead status of yellow catfish after infected with Ichthyophthirius multifiliis
2.2 黄颡鱼血红蛋白β链基因(HBβ)及其C端抗菌肽HBβ-C生物信息学分析
基于生物信息学分析, 黄颡鱼HBβ的cDNA全长为447 bp, 共编码148个氨基酸(aa; 图 3)。通过与抗菌肽数据库(http://aps.unmc.edu/AP/APD3_update2020_release.fasta)进行比较分析发现黄颡鱼抗菌肽HBβ-C与斑点叉尾鮰抗菌肽AP01619匹配度高达84.84%, 位于黄颡鱼血红蛋白β链基因的C端33 aa (图 3下划线)。抗菌肽HBβ-C的螺旋轮分布如图 4A所示, 其总电荷为+1.5, 总疏水率为51.8%。抗菌肽HBβ-C的模型结构图如图 4B所示, 由序列10PEVHETWQKFLNVVVAAL28形成α螺旋结构。

图3 黄颡鱼HBβ基因cDNA全长及其预测的氨基酸序列Fig. 3 The cDNA sequence and deduced amino acid sequences of HBβ in yellow catfish
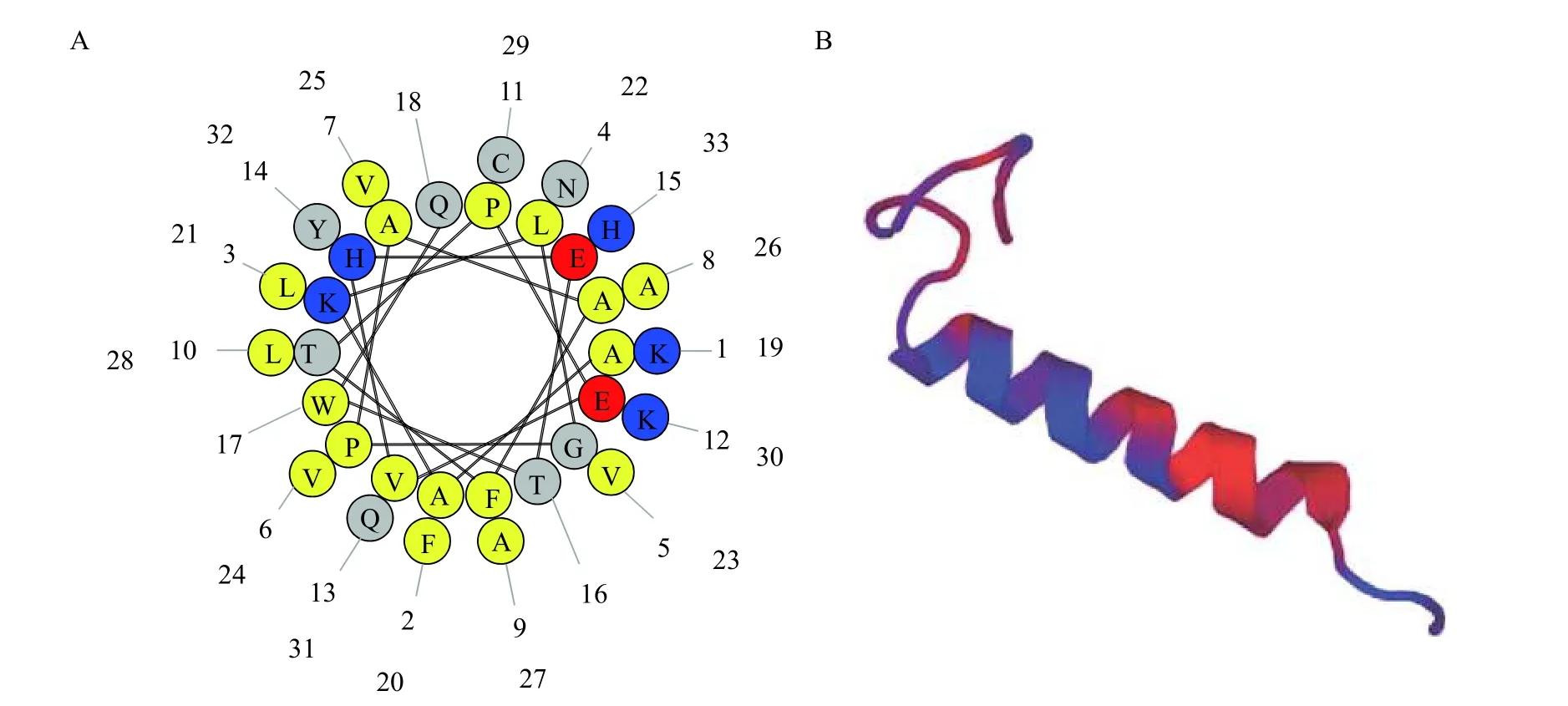
图4 抗菌肽HBβ-C的螺旋轮分布和结构模型图Fig. 4 The helical wheel distribution and structure model of the antimicrobial peptide HBβ-C
2.3 黄颡鱼HBβ在鳃和皮肤等免疫组织中mRNA和蛋白表达量分析
我们分别以全雄黄颡鱼和杂交黄颡鱼作为抗感染和易感个体, 选取多子小瓜虫的主要感染部位即皮肤和鳃组织检测HBβ的mRNA表达水平。定量结果如图 5A和5B所示, 在对照组中, 杂交黄颡鱼的鳃和皮肤组织中HBβ的mRNA表达量均高于全雄黄颡鱼; 在攻毒试验第5天,HBβ的mRNA表达量在杂交黄颡鱼鳃和皮肤组织中分别上升了3.7倍和1.5倍,而在全雄黄颡鱼鳃和皮肤组织分别上升了23.8倍和26.3倍, 此时全雄黄颡鱼的鳃和皮肤组织中HBβ的mRNA表达量显著高于杂交黄颡鱼。在未感染多子小瓜虫的鳃组织中, 全雄黄颡鱼HBβ蛋白表达量比杂交黄颡鱼高2.2倍(图 5C和5D), 表明全雄黄颡鱼HBβmRNA在鳃和皮肤等免疫组织中蛋白翻译水平明显高于杂交黄颡鱼; 在应对小瓜虫感染的过程中, 全雄黄颡鱼的mRNA转录水平快速提升, 远远高于杂交黄颡鱼, 可能与全雄黄颡鱼抗多子小瓜虫的特性相关。

图5 HBβ在鳃和皮肤组织中的表达分析Fig. 5 The relative expression of HBβ in gill and skin
2.4 抗菌肽HBβ-C对多子小瓜虫的体外杀伤影响
多子小瓜虫的生活史分为滋养体、包囊体和掠食体三个阶段。当使用50 μg/mL的HBβ-C处理后, 滋养体从黏液囊中逃出后在水中剧烈运动, 然后25s左右裂解, 胞质外流而死(图6A和6B); 仅需要15 μg/mL的HBβ-C能在3min内杀死所有滋养体(表 2)。由于包囊体具有较厚的包囊膜(图 6C), 抗菌肽HBβ-C较难渗透; 当使用500 μg/mL的HBβ-C处理后, 前40min包囊体的包囊膜迅速皱缩, 里面的子体急剧运动(图 6D), 之后包囊体中的子体逐渐逃脱游离在水中, 约1h后胞质外流最后死亡(图 6E和6F)。对于刚释放的掠食体来说, HBβ-C需要使用较高浓度(125 μg/mL)及花费更长时间(约3h)才能将其完全杀死(图 6G和6H)。所有对照组的3个阶段的多子小瓜虫完好无损, 各个阶段正常分裂发育。
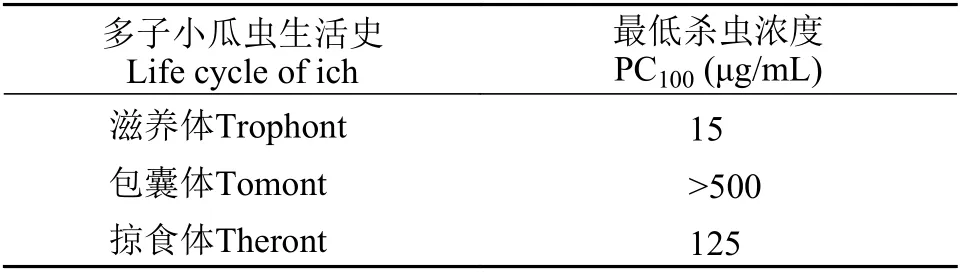
表2 抗菌肽HBβ-C对不同生活史的多子小瓜虫体外影响Tab. 2 The effect of antimicrobial peptide HBβ-C on the different life cycles of Ichthyophthirius multifiliis in vitro

图6 抗菌肽HBβ-C对不同生活史的多子小瓜虫的影响Fig. 6 The effect of antimicrobial peptide HBβ-C on Ichthyophthirius multifiliis different life cycles
2.5 HBβ在杂交黄颡鱼感染小瓜虫过程中的表达定位分析
为了研究HBβ在多子小瓜虫感染过程中的组织分布, 我们使用连续切片分别进行HBβ抗体荧光免疫组化和苏木精—伊红(HE)染色分析。荧光免疫组化结果显示, HBβ抗体在杂交黄颡鱼鳃组织中的红细胞中有较强的表达信号(图 7A和7B, 箭头所示)。此外, HBβ抗体能特异性结合到多子小瓜虫表面, 表现出很强的荧光信号(图 7C和7D, 箭头所示)。在皮肤组织中, 多子小瓜虫寄生位置较深紧靠着基底膜, HBβ抗体也特异性附着多子小瓜虫上(图7E和7F, 箭头所示)。
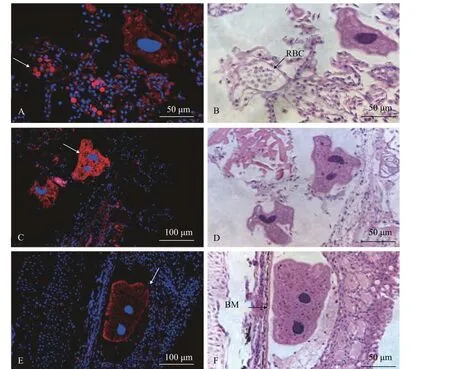
图7 HBβ免疫荧光定位及组织切片分析Fig. 7 Detection of HBβ localization by immunohistochemistry and analysis of histological section
3 讨论
多子小瓜虫病是水产养殖的主要病害之一, 国内外科研工作者早期对小瓜虫的防治药物进行了大量的筛选, 已筛选的有效治疗药物如孔雀石绿、硝酸亚汞和醋酸亚汞等, 均由于毒性强、致癌和严重污染环境而被禁用。近年来中草药上筛选的辣椒、生姜、五倍子等和化合物高锰酸钾、福尔马林、硫酸铜等有一定疗效, 但大多数药物仅对掠食体阶段有一定效果, 对滋养体阶段效果并不显著。小瓜虫病防治的最大难题就是一般药物不能有效渗透到小瓜虫的寄生部位杀死滋养体[27]。
血红蛋白广泛存在于脊椎动物和无脊椎动物的红细胞中, 是由2个α亚基和2个β亚基形成一个四聚体结构具有携氧功能的重要呼吸蛋白[28]。近年来, 研究者们发现血红蛋白除了具有携氧功能外,其亚基片段可形成各种活性肽, 具有止痛、促生长激素释放、阿片样活性、促肾上腺皮质激素释放和血管紧张素转换酶抑制活性等功能[29—33]。血红蛋白源的肽段可形成多种抗菌肽, 其抗菌特性在哺乳类、爬行类、鱼类和贝类中均有发现[34,35]。诸多研究发现血红蛋白α链35—56位氨基酸和位于血红蛋白β链的111—146位氨基酸片段存在抗微生物活性[23,36—41]。Ullal等[23]在斑点叉尾鮰的鳃和皮肤组织中发现位于血红蛋白β链的116—148位氨基酸的短肽, 其仅对小瓜虫滋养体阶段有较好的杀伤效果, 对掠食体和包囊体却没有明显效果。
我们前期在黄颡鱼的皮肤黏液蛋白质组中也曾发现血红蛋白β链的碳端序列HBβ-C, 其表达量在鲇爱德华氏菌感染后明显上升[24]。本研究合成的黄颡鱼HBβ-C抗菌肽对滋养体阶段效果最佳, 仅需要15 μg/mL的HBβ-C抗菌肽能在3min内杀死所有滋养体; 黄颡鱼HBβ-C处理滋养体后短时间内会被诱导分泌出黏液囊并急剧运动, 而这种黏液囊与正常滋养体脱离宿主后往包囊体分化时形成的黏液囊肿非常相似, 而同样对多子小瓜虫具有杀伤作用的抗菌肽Piscidin 1处理后却并未发现滋养体分泌黏液囊[42,43]。从诱导滋养体运动加剧和分泌黏液囊的作用效果来看, 抗菌肽HBβ-C可能诱导滋养体离开宿主并加速分化, 具体还需通过活体试验来进一步证实。此外, 黄颡鱼抗菌肽HBβ-C对掠食体也有杀伤效果, 且高浓度处理包囊体后能导致包囊膜皱缩, 导致体内子体旋转加剧甚至从包囊中逃逸出来最终死亡。以上表明黄颡鱼的HBβ-C抗小瓜虫能力高于斑点叉尾鮰的HBβ-C, 抗小瓜虫能力的差异可能是由于HBβ-C在2个物种中的序列差异造成的[23]。一般抗菌肽发挥功能涉及3个重要特征:α螺旋结构、抗菌肽整体带正电荷和较高含量的疏水性氨基酸。大量研究表明, α螺旋结构抗菌肽的阳离子电荷和疏水性氨基酸对抗菌作用至关重要[44—46]。黄颡鱼抗菌肽Hbβ-C大部分氨基酸呈α螺旋结构, 整体带+1.5电荷, 疏水率高达51.8%, 而在斑点叉尾鮰发现的抗小瓜虫肽的整体0电荷, 疏水率为45%。
斑点叉尾鮰易患鮰爱德华氏菌引起的鲶鱼肠道败血症(Enteric septicemia of catfish, ESC), 而蓝鲶(Ictalurus furcatus)对鮰爱德华氏菌具有较强的抗性, 其杂交种表现出中等抗性水平[47]。本研究发现全雄黄颡鱼对由多子小瓜虫引起的白点病具有抗性, 瓦式黄颡鱼易感染多子小瓜虫而快速死亡;但在养殖过程中也偶有报道全雄和普通黄颡鱼感染小瓜虫的案例, 这可能与养殖条件和鱼体本身的健康状态有关系。我们分析杂交黄颡鱼和全雄黄颡鱼在感染多子小瓜虫前后HBβ的表达发现, 虽然杂交黄颡鱼的鳃和皮肤组织中HBβ的mRNA表达量均高于全雄黄颡鱼, 然而HBβ蛋白在全雄黄颡鱼鳃组织中要明显高于杂交黄颡鱼, 这可能与翻译效率有关[48]。在感染小瓜虫后, 全雄黄颡鱼和杂交黄颡鱼的鳃和皮肤组织中HBβ的mRNA均显著上升, 且全雄黄颡鱼上升倍率更高。研究者发现感染多子小瓜虫产生免疫抗性的鲤中, 掠食体就可以侵入皮肤, 但在感染后2h 80%的多子小瓜虫消失了, 表明鱼体本身可针对寄生虫迅速产生反应抵抗入侵[49]。在多子小瓜虫感染后, 全雄黄颡鱼能快速合成大量HBβmRNA并高效率翻译成蛋白质, 产生的高水平HBβ-C抗菌肽可能有效抵御多子小瓜虫。此外, 黄颡鱼在感染多子小瓜虫后, 抗菌肽HBβ-C会富集在多子小瓜虫表面。多子小瓜虫的抑动抗原(Immobilization antigens)可诱导宿主在血清和黏液中产生抗体, 在体外会附着在不同生活史阶段的多子小瓜虫上[50]。Xu等[51]发现免疫球蛋白T和免疫球蛋白M广泛存在于虹鳟(Oncorhynchus mykiss)皮肤中,当多子小瓜虫感染时立即附着在其表面。抗菌肽导致膜破裂主要通过与寄生虫表面相互作用[52]。杀虫类抗菌肽发挥作用主要通过作用于细胞膜, 扰乱电化学梯度, 从而引起寄生虫渗透性休克, 这一过程主要涉及膜电位的迅速崩溃和细胞内ATP下降; 膜通透性增加; 形态改变如气泡或破裂[53]。
综上所述, 目前绝大多数药物仅对多子小瓜虫的掠食体阶段有效, 而对寄生在鱼体表的滋养体阶段很难发挥作用; 黄颡鱼抗菌肽HBβ-C对寄生在鱼体的滋养体阶段有较好的杀伤效果, 未来可探索利用生物工程技术对抗菌肽HBβ-C进行规模化生产,应用到水产养殖业中。结果证明, 全雄黄颡鱼对小瓜虫病具有抗性, 而杂交黄颡鱼易感。全雄黄颡鱼和杂交黄颡鱼的抗病性能存在差异, 在养殖过程中,可以根据养殖条件和养殖环境合理选择黄颡鱼的养殖品种。
