60Co-γ 辐射对扁穗雀麦种子萌发及其幼苗生长的影响
2021-08-03董志晓赵煜晗易莉美
袁 帅,董志晓,赵煜晗,杨 建,易莉美,聂 聪,凌 瑶,马 啸
(四川农业大学草业科技学院草业科学系,四川成都 611130)
辐射诱变具有安全、简单、突变率高等特征,相较于传统的育种方法,辐射诱变是扩大变异和选育优质新品种的行之有效的手段,且采用辐射诱变技术能够得到传统育种方法难以获得的或者自然界中罕见的牧草新基因、新性状,从而解决优异种质资源缺乏的问题[1]。目前,国内外研究主要以X、γ、α、β射线、紫外线和激光等作为辐射源,其中60Co-γ是辐射诱变最常用的方法之一,且已利用该方法育成了一系列高产、多抗的农作物新品种[2]。我国牧草育种工作一般是对野生资源进行大量评价,进而筛选出优异种质来选育品种,因其育种效率较低,所以急迫需要基于现有优良种质或品种进行人工创制优异的育种材料。在辐射诱变育种工作中,选择合适的辐射剂量与辐射源是辐射诱变育种的重要前提。利用0~300 Gy 的60Co-γ 射线对无芒雀麦(Bromus inermis)进行处理,发现随着辐射剂量的增加,种子活力指数、苗高和根长呈先升高后下降的趋势,并在50 Gy 时各项指标测定值最大[3]。利用0~200 Gy 的60Co-γ 射 线 对 草 地 早 熟 禾(Poa pratensis)进行辐射处理,发现低剂量(50 Gy)对草地早熟禾种子萌发和幼苗叶片的超氧化物歧化酶(superoxide dismutase, SOD)活性有促进作用,高剂量则产生抑制作用[4]。利用UV-B射线对甘草(Glycyrrhiza uralensis)进行辐射处理,发现UV-B射线对甘草种子萌发率、发芽势、幼苗株高、叶面积均有抑制作用,对幼苗的丙二醛(malondialdehyde,MDA)和脯氨酸(proline,Pro)含量则产生一定的促进作用[5]。
扁穗雀麦(B.catharticus)系禾本科(Poaceae)雀麦属的一年生或短期多年生草本植物,具有产量高、绿期长、适口性优良及再生能力强等优点[6],原生于南美洲潘帕斯草原,现在主要栽培于全球温带地区[7]。目前,国内外对扁穗雀麦的研究主要集中于盐胁迫[8]、干旱胁迫[9]和低温胁迫[10]等与抗逆相关的研究上,未见扁穗雀麦种子响应60Co-γ 辐射的生理生化研究。扁穗雀麦与辐射育种中较为成功的水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)等植物同属于禾本科,种子结构类似,因此其在辐射诱变育种上具备一定的可行性。鉴于此,探究不同辐射剂量处理对扁穗雀麦种子萌发以及早期幼苗生长的影响,关系到能否成功实现辐射M0代幼苗的建植,可为扁穗雀麦诱变辐射育种提供一定的科学依据及技术参考。
1 材料和方法
1.1 试验材料
‘川西’扁穗雀麦(2020年农业部审定品种)种子萌发率约为93%,由四川农业大学牧草课题组提供,于2016年收种保存在四川雅安−20℃冰箱中。种子在试验前用2%的萎锈灵处理24 h,以预防黑穗病,再冲洗干净后晒干待用。
1.2 试验设计
1.2.1辐射处理
将扁穗雀麦种子于2020年9月送至四川省农科院生物技术核技术研究所进行60Co-γ 辐射处理,辐射率1.84 Gy·min−1,辐射剂量依次为0(对照)、50、100、150、200、250和300 Gy,共7个辐射处理,每个处理100 g 种子。
1.2.2幼苗培养
选取不同辐射剂量处理下的种子各50粒,浸泡于35℃蒸馏水中2 h,每个处理10个重复,置于铺有两层滤纸的培养皿中,加入适量蒸馏水(以没过种子为宜),并放入植物生长室进行培养,培养条件为日温25℃ (16 h)、夜温16℃(8 h)。在第4天发芽高峰期记录发芽势,在第7天萌发率不再变化时记录萌发率。挑选出部分发芽种子移栽至加有石英砂和营养液的方形育苗盘中,继续放于植物生长室,条件同种子萌发一致,培养14 d 后进行幼苗和根系形态及生理相关指标的测定。由于后期观察到高辐射剂量处理后幼苗的畸变率非常高,难以建成完整的幼苗结构,所以幼苗的根系形态、幼苗的形态和生理指标均选用0~200 Gy 辐射处理下的幼苗进行测定。
1.3 指标测定及方法
1.3.1种子萌发指标
种子萌发指标包括发芽势、萌发率,其计算方法为发芽势=(种子第4天时的发芽数/供试种子总数)×100%,萌发率=(种子第7天时的发芽数/供试种子总数)×100%。
1.3.2半致死剂量
根据线性回归方程式y=a+bx确定半致死剂量。式中:y代表不同辐射处理导致的幼苗死亡率,x代表不同辐射剂量[11]。
1.3.3根系形态参数测定
用数字化扫描仪(WinRHIZO_Pro2016)扫描各处理下的幼苗根系,再利用配套软件WinRHIZO_Pro V2007d(Regent Instrument Inc.,Canada)获取幼苗根长(cm)、表面积(cm2)、平均直径(mm)和体积(cm3)等指标,每个处理10个重复。
1.3.4幼苗形态指标测定
将培养后处于三叶期的幼苗进行取样,每个辐射剂量处理选取10株,利用游标卡尺测定叶宽(展开第2片真叶)和茎粗,直尺测定苗高和叶长(展开第1片真叶),并计算各个辐射剂量处理的叶面积(叶面积校正系数0.73×叶长×叶宽)[12]。
1.3.5生理指标测定
取各辐射剂量处理的幼苗叶片,迅速放入液氮中,随后存贮于−80℃超低温冰箱中。依照《植物生理学实验》[13]对MDA、Pro和叶绿素(chlorophyll, Chl)含量以及过氧化氢酶(catalase,CAT)和SOD 活性进行检测,每个处理取3个重复样本。
1.4 数据处理
试验中幼苗及根系形态指标的数据采用SPSS 27.0进行单因素方差分析(One-way ANOVA),利用Duncan 新复极差法进行多重比较(P<0.05),运用Excel 2019及GraphPad Prism 8进行绘图。
为挑选出长势较好且形态指标较对照变异均较大的辐射剂量,本研究运用隶属函数法对各处理的各项指标进行综合分析比较,计算公式[14]:正向隶属函数值y=(xa−xmin)/(xmax−xmin),负向隶属函数值y=1−(xa−xmin)/(xmax−xmin)。式中:xa代表某一指标的值,xmax和xmin分别代表同一指标中的最大值与最小值。
2 结果与分析
2.1 60Co-γ 辐射对种子萌发指标的影响
随着辐射剂量的上升,扁穗雀麦种子发芽势呈先升高后降低的趋势(图1)。在50 Gy 时种子发芽势达到最大值,为32%,且与对照相比差异显著(P<0.05)。随着辐射剂量从50 Gy 升至300 Gy,种子发芽势持续下降。当辐射剂量为150~300 Gy 时种子发芽势均显著低于对照(P<0.05),辐射剂量超过200 Gy时种子发芽势约下降至对照的10%。除50 Gy 处理下种子萌发率略高于对照外(P>0.05),其余各辐射处理下均低于对照。随着辐射剂量从50 Gy 升至300 Gy,种子萌发率持续下降,且在辐射剂量为200~300 Gy 时种子萌发率显著低于对照(P<0.05),与对照相比分别降低了22.98%、55.42%和66.27%(图1)。上述结果表明,在低辐射(50 Gy)处理下,辐射对种子萌发率和发芽势均有促进效应,而在高辐射(100~300 Gy)处理下,则对种子萌发具有不同程度的抑制作用,且辐射剂量在300 Gy 时抑制作用最强。

图1 60Co-γ 辐射对扁穗雀麦种子萌发指标的影响Figure 1 Effects of 60Co-γ radiation on Bromus catharticus seed germination indices
2.2 辐射对幼苗植株和根系形态指标的影响
2.2.1辐射对幼苗形态的影响
随着辐射剂量从50 Gy 升至200 Gy,幼苗高度和叶长持续下降(表1)。除50 Gy 处理下的幼苗苗高与50和100 Gy 处理下的幼苗叶长略高于对照外(P>0.05),其余各辐射处理下幼苗苗高和叶长均低于对照。当辐射剂量达到150和200 Gy 时,苗高和叶长均显著低于对照(P<0.05)。除150 Gy 处理下幼苗叶面积低于对照外,其余处理的叶宽、茎粗和叶面积均高于对照。当辐射剂量为100 Gy 时幼苗叶面积达到最大,200 Gy 时幼苗叶宽和茎粗达到最大,且叶宽及茎粗与对照组相比分别显著增加了31.70%和42.27%(P<0.05)。总体来看,高辐射剂量会促使幼苗低矮化、叶片短粗化。

表1 60Co-γ 辐射对扁穗雀麦幼苗形态的影响Table 1 Effects of 60Co-γ radiation on the morphology of Bromus catharticus seedlings
2.2.2辐射对根系形态的影响
随着辐射剂量从100 Gy 升至200 Gy,根系长度均显著下降(P< 0.05),与对照相比分别降低了30.44%、38.97%和69.29%(表2)。除50 Gy 处理下幼苗根长略高于对照外(P>0.05),其余各辐射处理下的根长均低于对照。根系表面积和体积的变化均与辐射剂量的变化呈反比。当辐射剂量为200 Gy 时,根系表面积和体积均达到最小值,但根系平均直径在各辐射处理下与对照之间无显著差异(P> 0.05)。

表2 60Co-γ 辐射对扁穗雀麦根系形态的影响Table 2 Effectsof 60Co-γ radiation on root morphology of Bromuscatharticus
2.3 辐射对幼苗生理指标的影响
对不同辐射处理下扁穗雀麦幼苗叶片MDA 和Pro含量的分析比较发现,二者随着辐射剂量的上升均表现出先上升后降低的趋势(图2)。辐射剂量从50 Gy 上升至200 Gy 时,MDA 含量相较于对照分别增加了12.55%、45.54%、102.73%和8.67%,Pro含量分别增加了84.64%、230.74%、166.06%和0.88%。当辐射条件分别为100和150 Gy 时叶片Pro和MDA含量显著高于对照(P<0.05),且值为最大。同时,幼苗抗氧化酶SOD及CAT 活性随着辐射剂量的上升均表现出先上升后降低的趋势(图2)。在50 Gy时SOD活性达到最强,较对照显著上升了14.79%(P<0.05)。在50~200 Gy 辐射剂量下,相较于对照而言,CAT 活性均有显著提升(P< 0.05),且在150 Gy时,CAT活性最强。
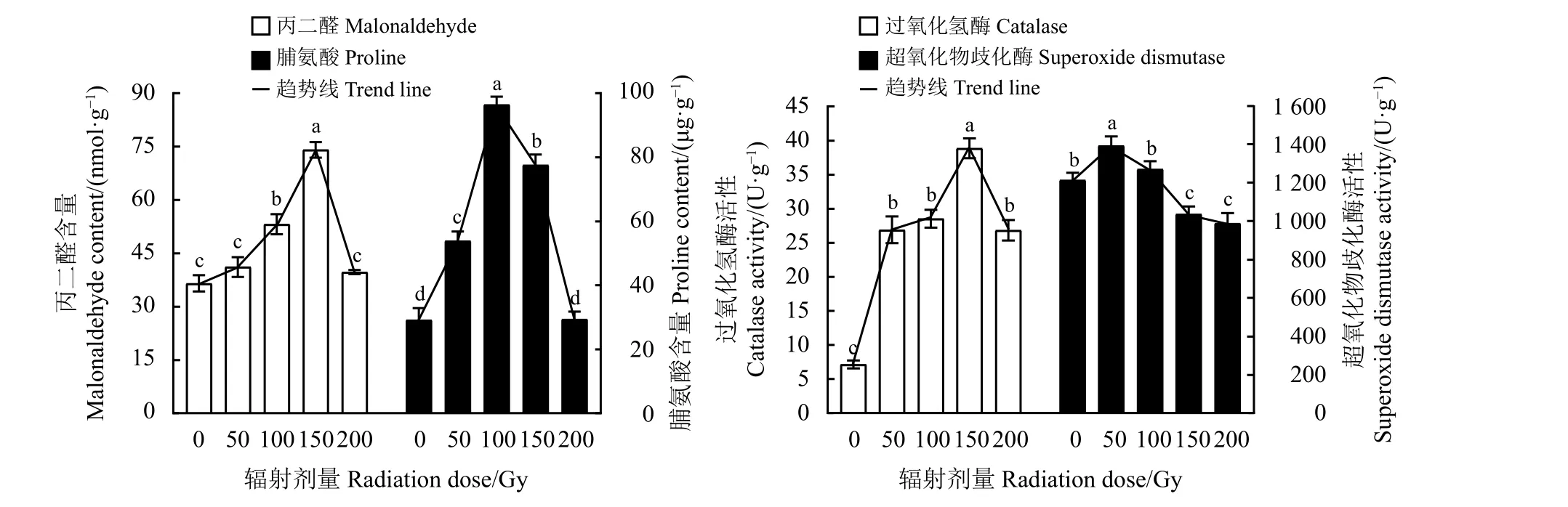
图2 60Co-γ 辐射对扁穗雀麦生理指标的影响Figure2 Effects of 60Co-γ radiation on physiological indicesof Bromuscatharticus
2.4 辐射对幼苗叶片叶绿素含量的影响
随着辐射剂量的升高,Chl a、Chl b和总Chl 含量均呈先上升后下降的趋势(图3)。当辐射剂量为50 Gy 时,Chl a、Chl b 和总Chl 含量均达到最大值,且显著高于对照(P<0.05);随着辐射剂量从50 Gy升至200 Gy,Chl a、Chl b和总Chl含量均持续降低。在高辐射剂量(150和200 Gy)下,Chl a、Chl b和总Chl 含量相较于对照均显著下降(P< 0.05)。

图3 60Co-γ 辐射对扁穗雀麦幼苗叶片叶绿素含量的影响Figure 3 Effect of 60Co-γ radiation on leaf chlorophyll content in seedlings of Bromus catharticus
2.5 半致死剂量的确定
利用线性回归方程式,绘制标准曲线,可得标准曲线方程为y= 0.199 3x+ 5.392 9,R2= 0.857 1(图4)。把y=50代入方程式,求得x=223.82,即扁穗雀麦种子受60Co-γ 辐射的半致死剂量为223.82 Gy,约为224 Gy。当y≠50时,需要以实际对照种子的萌发率来计算。

图4 辐射剂量与死亡率的一元线性回归分析Figure 4 Univariate linear regression between radiation dose and percentage mortality
2.6 隶属函数综合分析
各辐射剂量处理下扁穗雀麦各指标的变化表现出一定程度上的相关性和差异性,且采用一种指标并不能全面地解释辐射对扁穗雀麦种子萌发和幼苗生长的影响,所以需要对多指标进行综合评价,以筛选出最适辐射剂量。根据各处理下测定的各项指标数值,包括2个萌发指标、5个幼苗形态指标、4 个根系形态指标和7个生理形态指标,计算其隶属函数值(表3),值越大,说明诱变后种子和幼苗的生长势与活力越强。结果表明:不同辐射处理下平均隶属函数值排序表现为50 Gy>100 Gy> 0 Gy>150 Gy>200 Gy>250 Gy>300 Gy,即50和100 Gy
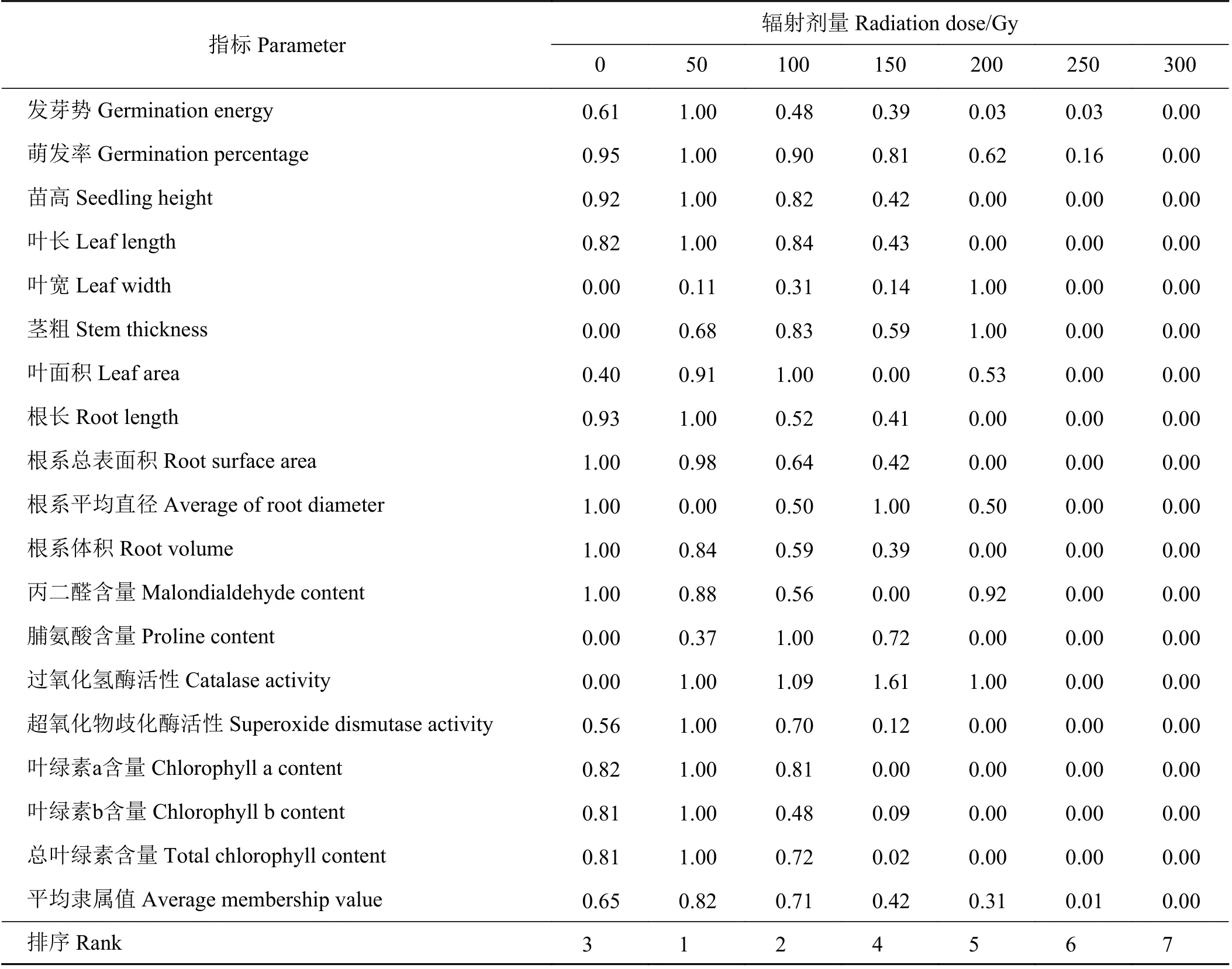
表3 基于多指标利用隶属函数分析评价扁穗雀麦种子辐射后生长效应Table3 Membership function analysisbased on multipleindicatorsfor theeffectsof radiation on Bromus catharticus
辐射剂量最适用于扁穗雀麦的辐射诱变育种,且后代存活率和生长势较好。
3 讨论与结论
本研究表明,60Co-γ 射线辐射对扁穗雀麦种子萌发及幼苗生长有显著的影响。低辐射剂量(50 Gy)对扁穗雀麦种子萌发具有促进作用,此时种子的萌发指标及幼苗的苗高、叶长、叶宽、茎粗、叶面积和根系的长度、表面积均有提高;而高辐射剂量(100~300 Gy)对扁穗雀麦种子萌发有一定的抑制作用,且幼苗叶片出现黄叶和紫叶现象。在苜蓿(Medicago sativa)[15]和桂花(Osmanthus fragrans)[16]等植物的研究结果中也有类似情况。低辐射处理有助于种子的生长发育,这可能是因为种子内部有关酶的活性或生物自由基在合适的辐射剂量条件下发生改变,以加快种胚组织细胞的分裂与分化[17];而高辐射处理可能会破坏种胚组织或改变种子内多种生理生化作用,从而降低了种子活力。
MDA 含量的高低可以作为植株个体受外界胁迫大小的衡量标准。当植株受到外界胁迫时,植株体内会出现膜脂过氧化作用,产生导致植株某些部位受损的膜质过氧化产物MDA,从而改变植株脂质过氧化的速度和强弱,其含量与植物受损程度呈正相关关系[18]。本研究中,随着辐射剂量的上升,MDA含量表现出先升高后降低的趋势,各处理的MDA含量均高于对照,在150 Gy 处理下MDA 含量达到最大值,说明不同剂量60Co-γ 射线处理均对扁穗雀麦幼苗造成了膜质过氧化损伤,致使机体活性氧与自由基积累增加,从而导致膜结构的完整性被破坏,且在150 Gy 辐射剂量下破坏程度最大,类似结果在白刺花(Sophora davidii)[19]和高羊茅(Festuca elata)[20]等饲用植物中也有报道。在200 Gy 下,MDA 含量有所降低,这也许是由于高辐射剂量(200 Gy)加强了植物体内某些修复酶活性,膜脂过氧化受到抑制,从而降低了植物受损的程度。Pro作为渗透调节物质,能够增加细胞液浓度、降低渗透势,从而增加植物对水分的吸收,同时能够提高细胞膜的稳定性,从而在植物受到辐射或其他逆境胁迫时发挥重要作用[21]。本研究中,随着辐射剂量的上升,Pro含量表现出先上升后下降的趋势,表明中低辐射剂量下脯氨酸的积累对幼苗有一定的保护作用,而高剂量辐射下由于生理系统被强烈破坏导致脯氨酸含量下降。在绣球(Hydrangea macrophylla)[22]和无芒雀麦[23]的研究结果中也有类似情况。SOD作为一种保护酶,可以减少或消除超氧自由基对细胞膜的损害,其活力大小可以作为衡量植物抗氧化能力强弱的指标,属于植物体内保护酶的组成成分之一[24]。CAT 属于一种清除性酶类,具有抑制植物受H2O2毒害的作用,因为其可以促进H2O2分解为水和氧离子[25]。本研究中,随着辐射剂量的上升,SOD与CAT 活性均表现出先升高后降低的趋势,类似结果在露地菊(Chrysanthemum morifolium)[26]和草地早熟禾[4]中也有报道。本研究中,当辐射剂量分别为50MDA 含量的高低可以作为植株个体受外界胁迫大小的衡量标准。当植株受到外界胁迫时,植株体内会出现膜脂过氧化作用,产生导致植株某些部位受损的膜质过氧化产物MDA,从而改变植株脂质过氧化的速度和强弱,其含量与植物受损程度呈正相关关系[18]。本研究中,随着辐射剂量的上升,MDA含量表现出先升高后降低的趋势,各处理的MDA含量均高于对照,在150 Gy 处理下MDA 含量达到最大值,说明不同剂量60Co-γ 射线处理均对扁穗雀麦幼苗造成了膜质过氧化损伤,致使机体活性氧与自由基积累增加,从而导致膜结构的完整性被破坏,且在150 Gy 辐射剂量下破坏程度最大,类似结果在白刺花(Sophora davidii)[19]和高羊茅(Festuca elata)[20]等饲用植物中也有报道。在200 Gy 下,MDA 含量有所降低,这也许是由于高辐射剂量(200 Gy)加强了植物体内某些修复酶活性,膜脂过氧化受到抑制,从而降低了植物受损的程度。Pro作为渗透调节物质,能够增加细胞液浓度、降低渗透势,从而增加植物对水分的吸收,同时能够提高细胞膜的稳定性,从而在植物受到辐射或其他逆境胁迫时发挥重要作用[21]。本研究中,随着辐射剂量的上升,Pro含量表现出先上升后下降的趋势,表明中低辐射剂量下脯氨酸的积累对幼苗有一定的保护作用,而高剂量辐射下由于生理系统被强烈破坏导致脯氨酸含量下降。在绣球(Hydrangea macrophylla)[22]和无芒雀麦[23]的研究结果中也有类似情况。SOD作为一种保护酶,可以减少或消除超氧自由基对细胞膜的损害,其活力大小可以作为衡量植物抗氧化能力强弱的指标,属于植物体内保护酶的组成成分之一[24]。CAT 属于一种清除性酶类,具有抑制植物受H2O2毒害的作用,因为其可以促进H2O2分解为水和氧离子[25]。本研究中,随着辐射剂量的上升,SOD与CAT 活性均表现出先升高后降低的趋势,类似结果在露地菊(Chrysanthemum morifolium)[26]和草地早熟禾[4]中也有报道。本研究中,当辐射剂量分别为50和150 Gy 时,幼苗SOD和CAT活性最强,随后有所下降,这可能是由于低剂量未导致CAT 与SOD酶活性系统受到损伤,可以消除机体内过多积累的氧化性物质,有助于植株体内一些修复酶活性的提升,从而能够抵御各种胁迫环境;而当辐射剂量过高时,幼苗体内累积过多的过氧化物及自由基,造成一系列膜透性损伤,导致SOD与CAT 活性下降。
Chl 是与光合效应有关的色素之一,是植株进行能量及物质代谢的基本物质,同时可以反映在不利环境下植株的受损程度[27]。本研究中,扁穗雀麦种子受到60Co-γ 射线辐射后,叶片的Chl a、Chl b和总Chl 含量先随辐射剂量的增加而增加,当辐射剂量上升到50 Gy 时,Chl a、Chl b和总Chl 含量均达到最大值,此时可能是由于扁穗雀麦叶片为了提高自身的光合速率而增加叶绿素含量,以抵御逆境胁迫的危害,这有助于叶片的光合效应,对植株生长产生一定的促进作用。而在辐射剂量为100~200 Gy下,其含量均低于对照,可能是由于60Co-γ 射线辐射扁穗雀麦种子破坏了叶绿体的结构,使叶绿素含量降低,从而影响植株的生长。这与使用60Co-γ 射线对紫薇(Lagerstroemia indica)[28]辐射后使叶片叶绿素含量呈先升高后下降趋势的研究结果一致。而在对露地菊[26]的研究中发现,其变化趋势为先下降后上升,这与本研究结果不同,可能是由于各个植株对60Co-γ 射线的敏感度不一致所致。
本研究利用60Co-γ 射线对扁穗雀麦种子进行辐照处理,当辐射剂量为50和100 Gy 时,在种子萌发、幼苗生长、减轻膜质过氧化程度、维持细胞渗透势以及加快消除自由基等方面表现最优。经推算,60Co-γ 辐射扁穗雀麦种子的半致死剂量为224 Gy。上述结果对于创制扁穗雀麦新种质提供了一定的技术支撑和理论参考。
