洱海湿地昆虫群落网络结构与自适应性研究
2021-06-30郝鸿俊祝远波罗桂兰
郝鸿俊,王 熙,张 梅,刘 璇,祝远波,罗桂兰*
(1.大理大学工程学院,云南大理 671003;2.大理大学数学与计算机学院,云南大理 671003)
昆虫是数量最多的动物,占所有动物总数的73%〔1〕。昆虫能在自然界营养循环等生态系统服务区域中发挥重要作用〔2〕,并可以作为监测湿地生态环境和生物多样性的指示性物种〔3〕。湿地昆虫种类繁多,群落结构也很复杂。韩争伟等〔4〕对太湖湿地昆虫群落结构及多样性进行研究,利用多样性特征指数等参数找到了影响太湖湿地昆虫群落的重要因子为湿地植被类型和人工干扰。何云川等〔5〕对银川4种湿地生境的陆生昆虫群落多样性与稳定性进行研究,对4种生境群落特征指数聚类分析,结果表明昆虫群落多样性随季节时序变化。可见昆虫对环境的变化十分敏感,环境发生变化将会导致昆虫的数量和群落结构变化〔6〕。
洱海是云南省第二大淡水湖,北起洱源,长约42 km,周边有丰富的湿地资源。但是随着洱海周边旅游业的开发,洱海湿地生态环境遭到了一定的破坏影响。目前洱海生态保护已经受到人们的广泛关注,湿地建设工作也在紧张进行。本文选取洱海3个典型的湿地作为研究对象,3个湿地拥有不同的植被类型,受人为干扰程度也大不相同。通过调查和分析2018年11月至2019年10月不同生境下的昆虫群落多样性、均匀度、优势集中性指数等,探讨昆虫多样性的季节性动态变化和适应能力。该研究将为洱海湿地生物多样性保护与湿地建设方案提供理论支持和数据分析样本。
1 研究区域与昆虫环境样本数据
洱海位于云南省大理市郊区,面积约251 km2,总蓄水量30亿m3。大理属亚热带高原季风气候,是孟加拉湾西南季风带的迎风区,所以风较大,紫外线较强,光照充足,年平均气温15.7℃,年均降雨量1 000~1 200 mm,降水充沛〔7〕。
本文选取的3个湿地为洱源东湖湿地、洱海月湿地和罗时江湿地。湿地信息如表1所示,各区域环境差异较大。
本文数据样本由国家自然科学基金项目“洱海湿地昆虫群落普适生境网络生态行为研究”提供。该项目的主要研究目标是通过构建昆虫生境关系网络,探讨各个生境因子之间、生境因子与昆虫群落结构之间、人类行为与生境因子之间的相关性。目前已经完成了昆虫生态网络模型的定义,生成了3个湿地的昆虫群落网络拓扑图,并对网络特征参数进行了分析。本文将会在原有昆虫生态网络模型的基础上进行改进,生成2018年11月至2019年10月的昆虫生态网络关系拓扑图。样本采集信息见表2。
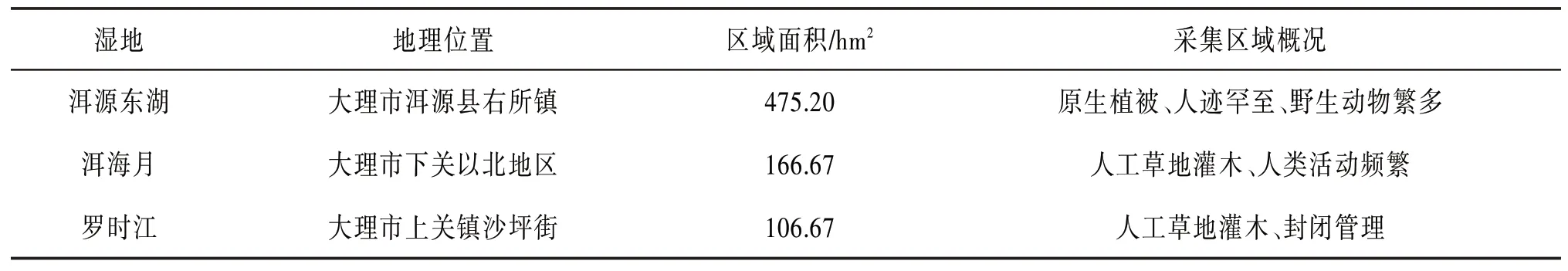
表1 洱海湿地信息
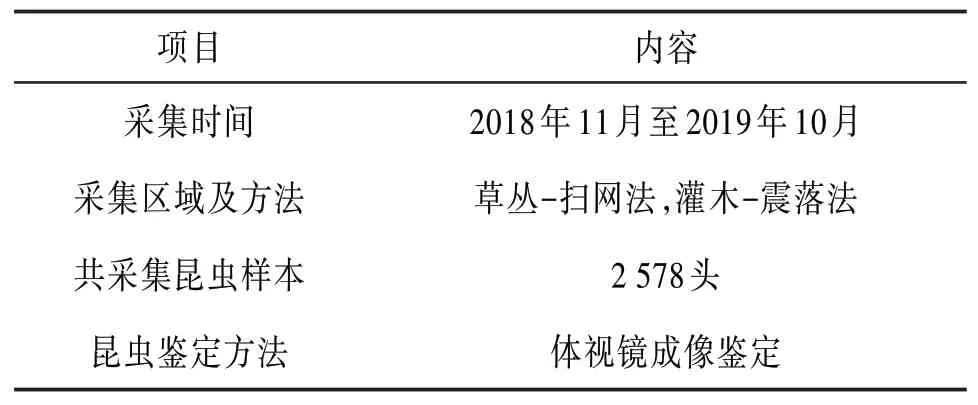
表2 昆虫数据样本采集信息
昆虫捕捉采用自制捕虫网,网口直径约33 cm。草丛每5 m进行一次扫网,一共扫20网。采集到的昆虫将其置于鉴定盘里进行初步分类。初步分类后将会对采集物种进行标号并置于离心管中保存。离心管中注入乙醇防止昆虫尸体毁坏。最后,根据昆虫分类法〔8〕利用体视镜成像鉴定出昆虫的纲、目、科、种等信息。鉴定信息汇总后得到各个湿地不同采集区域每个月的昆虫样本数据。体视镜成像照片标号与昆虫样本标号对应,方便后期数据核查。
环境数据包括气象数据和生境植被数据。其中气象数据是通过放置生态环境监测点,周期性采集了温度、湿度、降雨量、光照等10余项环境数据样本。生境植被数据通过湿地调研、取样鉴定,共鉴定出60余种植被种类。3个湿地的植被种类见表3。
洱源东湖湿地作为人为干扰最小的湿地,将其作为另外两个湿地植被种类的对照组具有较强的意义。

表3 湿地植被种类
3个湿地鉴定出的植被种类数量相当,但是洱海月湿地的灌木、乔木种类及数量占比要高于洱源东湖湿地,其原因是洱海月湿地为人为建设的湿地保护区,目前作为湿地公园向游客开放。湿地内的大部分植被都为外来引进。灌木、乔木不仅具有生态功能,同时还具有为人类遮阳防风等功能,并且作为湿地生态景观,其草本植物种类虽然较少,但草本植被分布均匀,空间结构较好。
罗时江湿地虽然也是人为建设的湿地保护区,但目前已封闭管理。该湿地最主要的功能为生态保护。植被类型多为草本植物,其数量与洱源东湖相当。相比另外两个湿地,该湿地灌木、乔木分布较稀疏,所以视野较开阔,地面光照较充足。
洱源东湖湿地作为最原始的湿地保护区,其植被类型少有外来引进。由于没有人为干扰,该区域植被分布较为紧密。
2 昆虫群落结构参数描述与定义
参数1 相对多度Pi指第i个物种个体数占总个体数的比例。其中,M i表示第i种物种的个体数,M表示昆虫群落内采集的物种总个体数。计算公式为

参数2 物种多样性指数H′表征昆虫对一定环境的适应性和物种在特定空间范围里的演化程度,也表征昆虫群落内所拥有物种种类数目多样的程度〔9〕。计算公式为
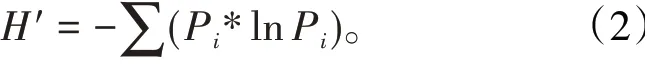
参数3 物种丰富度指数lnS,S为该区域采集到的物种数,丰富度指数lnS越大,表明该区域的物种种类越丰富,反之物种就越单一〔4〕。
参数4 物种均匀度指数J指所有昆虫个体在群落中的分布情况。计算公式为

群落稳定性与物种均匀度指数密切相关,J越高表明昆虫群落的稳定性越强〔10〕。
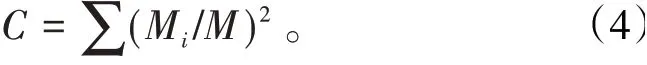
参数5 优势集中性指数C用来描述昆虫群落组成状况。计算公式为C反映了昆虫群落内物种发展的优势,C越大,群落中的个体数越集中在少数类群上,H′就会越小〔11〕。
参数6 昆虫群落稳定性指标,计算公式为

S s∕S i反映种间数量上的制约作用,比值越大,相对稳定性越强。S n∕S p反映食物网关系的复杂程度和相互制约的程度,比值越大,天敌物种数所占比例越大,群落稳定性越强〔12-13〕。
定义1 优势类群指昆虫群落中更适应环境的物种,即个体数占采集总个体数比例大于10%的物种〔14〕。
定义2 常见类群指昆虫群落中常见的物种,即个体数占总采集个体数比例大于等于1%的物种〔14〕。
定义3 稀有类群指在昆虫群落里很少采集到的物种,即个体数占总采集个体数比例小于1%的物种〔14〕。
定义4 昆虫群落时序演化结构指昆虫群落的物种数、个体数、均匀度等参数随时间(如季节等)变化所表现出来的动态结构。
3 昆虫群落网络拓扑结构建模
在文献〔3〕原有昆虫生态网络模型中,根据复杂网络理论,以昆虫种群为网络节点,以每月采集的昆虫物种的个体数量构建长度为12的时间序列,分别求出两个物种间个体数量变化的皮尔逊相关系数,并根据相关系数的数值大小定义边的权重,最后使用Gephi软件〔15〕建立昆虫生态网络节点和边的数据资料库,以此生成昆虫生态网络关系拓扑图〔3〕。
为了降低由于采集人员不同带来的误差,本文在原有昆虫生态网络模型的基础上做出改进。以每个月昆虫种群所占该区域采集昆虫个体的数量比例构建时间序列,求出各物种间数量比例的相关性作为边。构建出的昆虫生态网络模型拓扑图的节点面积大小表征某区域昆虫种群大小,节点颜色深浅表征节点度大小。边表征昆虫数量比例间的相关性。生成的3个区域的昆虫生态网络关系拓扑图见图1~3。
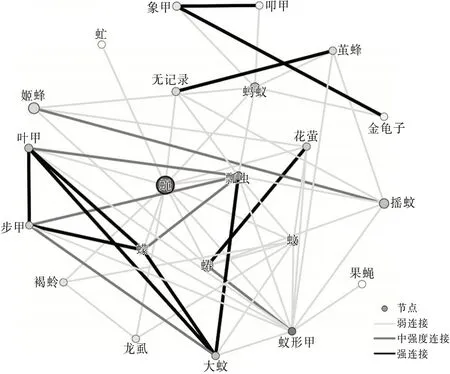
图1 洱源东湖湿地昆虫生态网络关系拓扑图
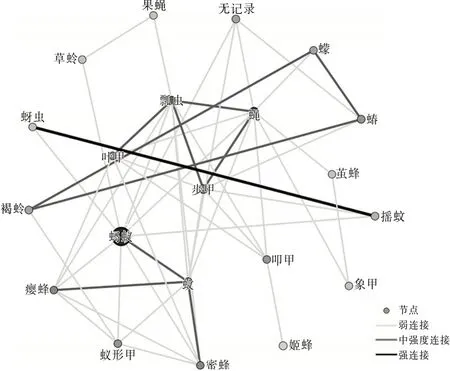
图2 洱海月湿地昆虫生态网络关系拓扑图
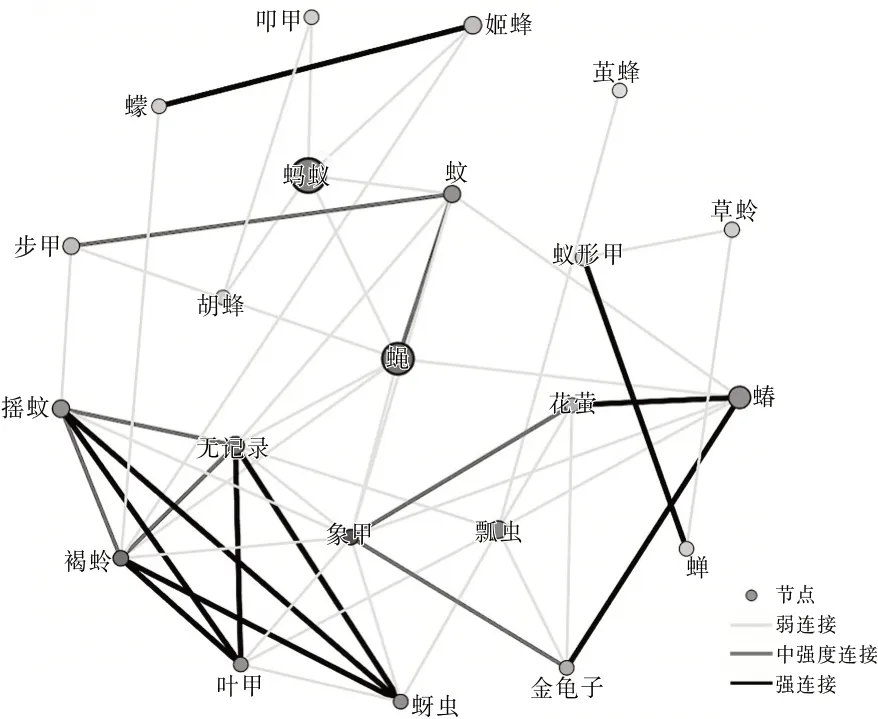
图3 罗时江湿地昆虫生态网络关系拓扑图
从3个区域的网络关系拓扑图可以看出蝇的节点颜色较深,可见蝇和其他物种的相关性比较强,是网络中的中心(HUB)节点。这里的HUB节点的节点度值较高,表征该节点与其他节点连接紧密。果蝇在洱源东湖和洱海月湿地中都为低节点度节点,并且在罗时江湿地中没有捕获到该物种。草蛉在洱海月和罗时江湿地中都为低节点度节点,并且在洱源东湖湿地中没有捕获到该物种。这类型节点与其他物种的相关性较低。同时有明显特点的还有蚂蚁节点,洱海月和罗时江的蚂蚁种群都比较大。可见蝇、蚂蚁都是湿地昆虫生态网络的关键物种。
根据文献〔3〕计算方法得到网络特征参数。平均加权度:洱源东湖>洱海月>罗时江;平均聚类系数:罗时江>洱海月>洱源东湖。可见:洱源东湖湿地昆虫群落的昆虫节点连接最丰富,关系最密切,网络的鲁棒性较好。罗时江节点之间的相互影响力强,当网络节点种类、数量发生变化时能够迅速做出调整,具有较强的调节能力。
4 昆虫群落结构及特征分析
4.1 昆虫群落组成分析 3个湿地在一年内共采集到2 422头昆虫,隶属于11目。3个湿地在一年内均采集到约30种昆虫类群,其中约2∕3的昆虫类群在3个湿地均有采集到,但是3个湿地昆虫目个体数所占比例差异较大。3个湿地昆虫群落结构组成成分见表4。
根据表4可知:洱源东湖湿地的双翅目、膜翅目、半翅目和鞘翅目昆虫类群,洱海月湿地膜翅目、鞘翅目和双翅目昆虫类群,罗时江湿地双翅目、膜翅目、鞘翅目和半翅目昆虫类群,均超过所在区域总数量的10%。
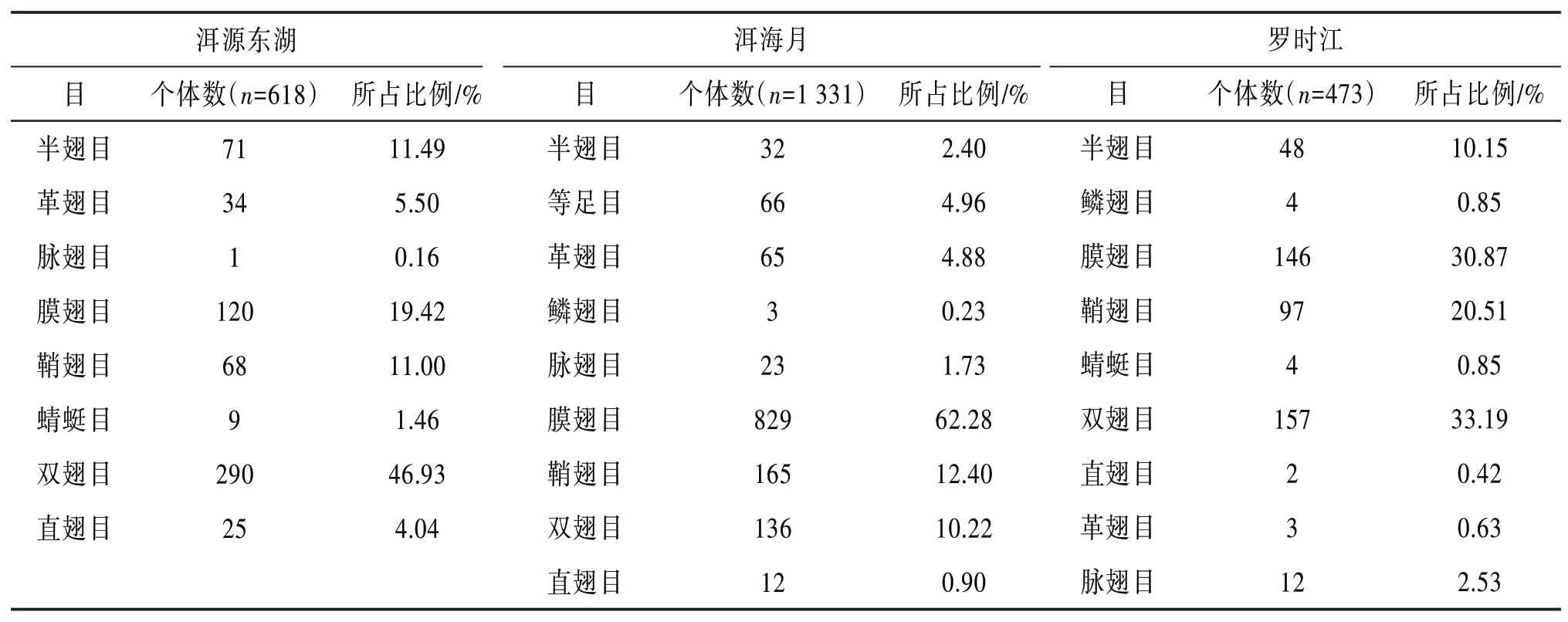
表4 昆虫群落结构组成成分
表5为一年中3个湿地昆虫群落的物种占比表。
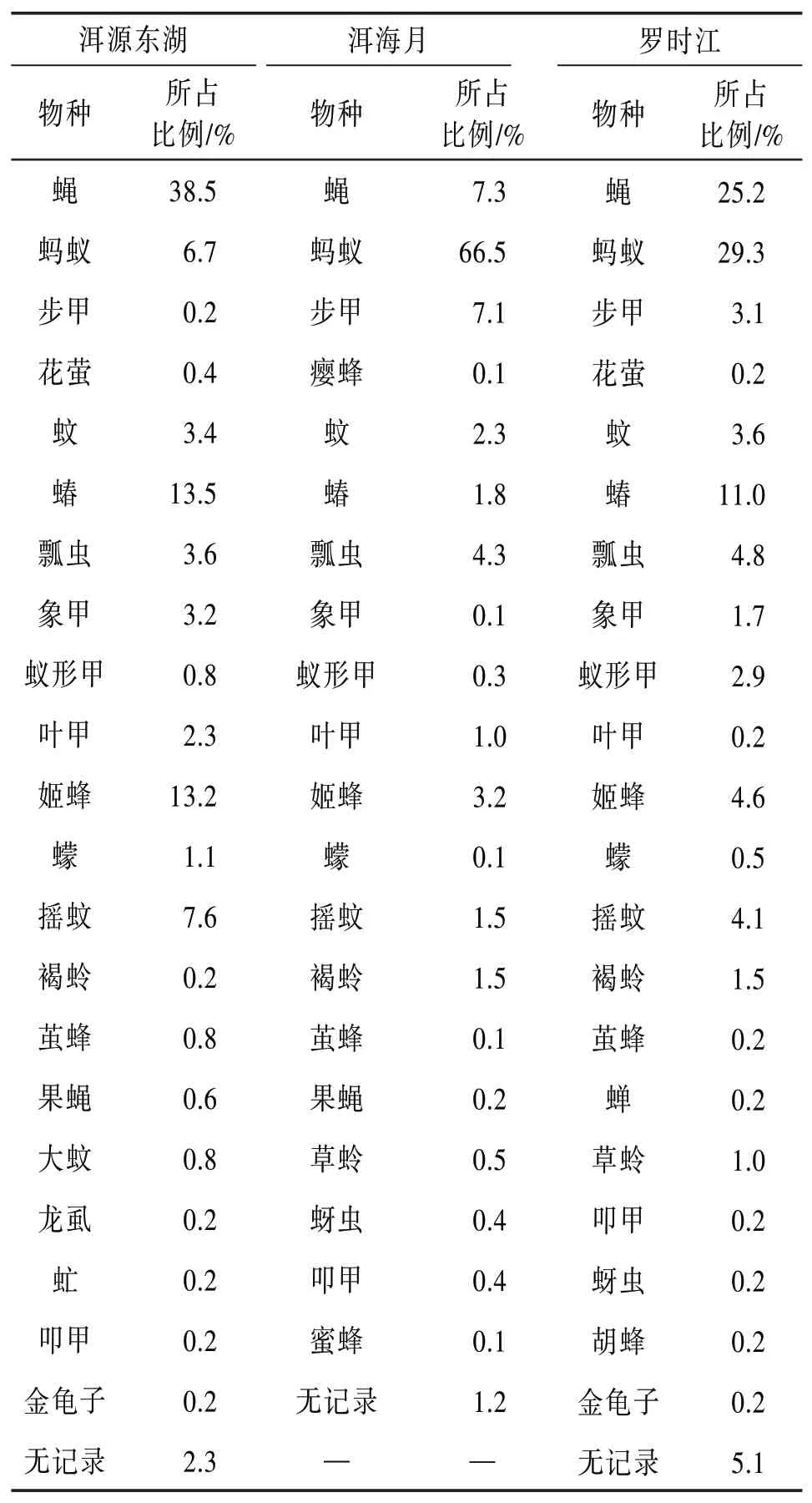
表5 洱海湿地昆虫群落物种占比
根据定义1~3,再结合表5所示结果,整理出3个湿地的种群类别见表6。
由表6可知,蝇是洱源东湖和罗时江的优势类群,但却是洱海月的常见类群。常见蝇类多以腐败的动植物以及人与动物的食物、排泄物等为食。洱源东湖湿地最接近原始生态,没有人为清理腐败植物,没有占领野生动物的活动区域,区域内有大量动物的粪便,这为蝇的生存繁殖提供了很好的场所。而罗时江和洱海月的植被都有人为定期修剪清理,并且洱海月的人类活动频繁,野生动物相对较少。这些因素导致蝇在3个湿地的数量占比差距悬殊。同理:蚂蚁是洱海月和罗时江的优势类群,但却是洱源东湖的常见类群。蚂蚁通常生活在干燥的区域,而大理降雨充沛,湿地多有积水。洱源东湖由于没有人工干预,多为坑洼的泥泞区域,容易积水。而另外两个湿地由于人工定期对草地灌木修整,阳光更容易照在地面上,地面水分蒸发较快,为蚂蚁提供了更好的栖息场所。
综上分析,人为干扰对昆虫群落结构造成了巨大的影响。优势类群决定了影响该区域生态环境的直接因素;常见类群反映出影响该区域生态环境的主要因素,该类群维持着区域生态环境的稳定性;而稀有类群表征了昆虫的多样性,反映出该区域生态环境在遭受到破坏以后的系统自我修复能力。
4.2 昆虫群落结构特征参数比值分析 取3个湿地一整年的昆虫群落数据样本,根据式(2)~(4)计算出洱海湿地昆虫群落的多样性特征参数,对比分析湿地昆虫群落的多样性及稳定性。以洱源东湖特征参数为基准“1”,其余两个湿地特征参数与洱源东湖特征参数比值的柱状图见图4。

表6 昆虫种群类型
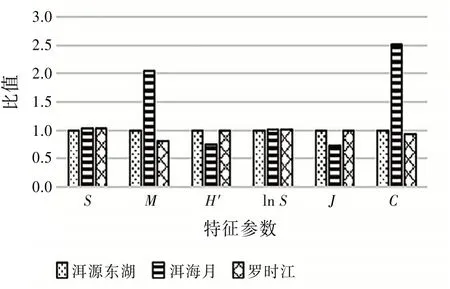
图4 3个湿地特征参数比值柱状图
由图4可见:物种多样性指数H′和物种均匀度指数J规律保持一致,均为洱源东湖>罗时江>洱海月。而优势集中性指数C表现为洱海月>洱源东湖>罗时江。其中洱源东湖和罗时江的各个参数相差较小,与洱海月的参数相差较大。由于各个湿地采集到的物种种类数目近似,所以物种丰富度指数lnS也近似。各参数表明洱源东湖和罗时江湿地的昆虫群落物种丰富,物种数量也相对均衡,没有极端优势物种,与洱海月的昆虫群落比较要相对稳定。
罗时江湿地经过人为建设保护,种植了特定的植物。但是由于后期封闭管理少有人员活动,所以昆虫群落结构良好,与原生湿地洱源东湖相差无几,可见人工修复湿地能起一定的作用。但是由于人类活动频繁,导致洱海月湿地昆虫群落结构较为脆弱,所以人为干预湿地更应重视昆虫多样性保护,提高生态系统的自我修复能力。
4.3 昆虫群落时序演化结构分析 根据昆虫生态环境数据样本,按照式(1)~(4)计算,统计分析得到昆虫群落结构特征参数随时间动态演化趋势。
由图5可见:3个湿地全年的多样性指数大多集中在1.0~2.0这个范围内,其中最大值出现在9月(2.365 1)的洱源东湖湿地,最小值出现在5月(0.665 1)的洱海月湿地。洱源东湖湿地的多样性指数变化趋势相对平缓,从2018年11月(0.893 6)开始缓缓上升,说明该区域的昆虫对生态环境的适应力强,只有在2019年4月(1.595 5)和2019年8月(1.538 9)发生明显的下降趋势。从物种来看,洱源东湖4月采集的物种以膜翅目和双翅目为主,没有采集到以蝽为主的半翅目。蝽的第一代成虫一般出现在5月,第二代至第四代分别在6月上旬、7月中旬和8月中旬出现。蝽成虫的寿命多为30~50 d。5月至8月采集到的蝽数量分别为3、13、12、31头。这导致在8月蝽成了优势类群。短期内对昆虫群落的多样性造成了一定的影响。
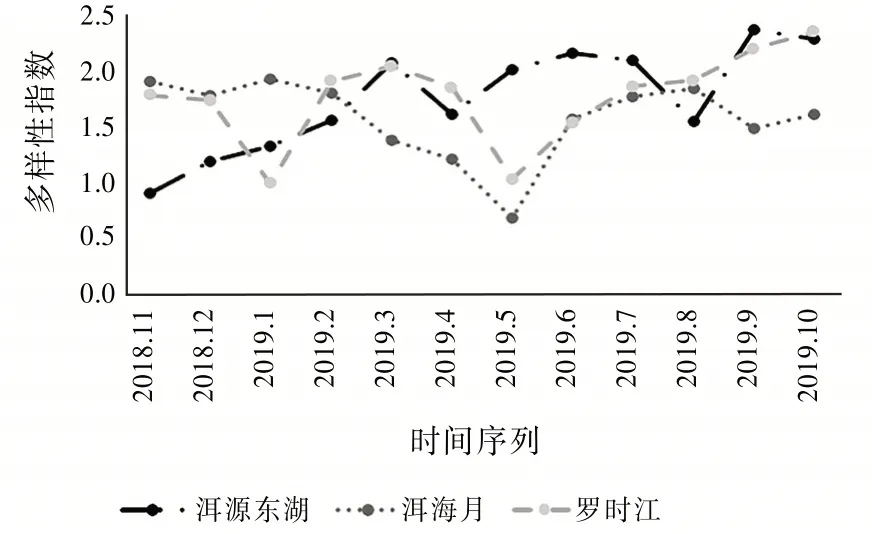
图5 多样性指数变化趋势
洱海月全年的多样性指数变化呈“V”形变化趋势,在2019年5月(0.665 1)达到全年的最低值,但是在2019年6月之后逐步上升至上半年的平均水平,说明洱海月湿地具有一定的自我调节能力,昆虫群落在受到一定的干扰破坏之后又能缓慢恢复。罗时江全年的多样性指数呈“W”形变化趋势,分别在2019年1月(0.976 3)和5月(1.013 5)到达谷底。说明罗时江湿地的昆虫群落结构变化率较大,但即便如此在全年的大多数时间里,罗时江湿地的多样性指数都高于洱海月湿地。罗时江湿地因为有更丰富的昆虫物种,即使在外在因素导致群落发生变化时也能迅速做出自我调整,从而保持对该区域环境的自适应能力。
图6的均匀度指数和图5的多样性指数的变化趋势相似。其中,洱海月的均匀度指数常年都低于另外两个湿地,说明该区域内的昆虫群落稳定性相对较弱。具有自然生态环境的洱源东湖变化趋势最平缓,可见原生的生态环境对昆虫群落稳定性的维持起着重要作用。
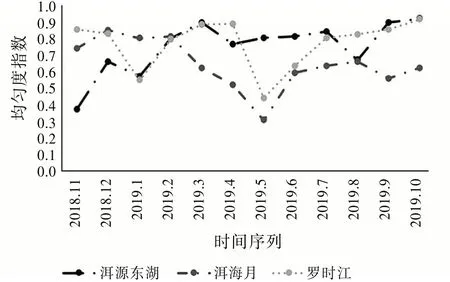
图6 均匀度指数变化趋势
图7的丰富度指数能直观反映采集到的物种数量变化。其中洱源东湖和洱海月的丰富度指数在2018年11月至2019年3月变化趋势雷同,在该时间段丰富度指数波动明显,并且整体有所下降。而罗时江的丰富度指数变化趋势虽然和另两个湿地近似,但是却滞后了一个月,这可能与罗时江温度低于其他两个湿地有关。在2019年4月至2019年10月洱源东湖湿地和罗时江湿地变化趋势近似,分别在6月至8月有一定的下降,而在8月之后又有所上升。可见,洱海湿地在冬季气温较低时物种丰富度指数较小,并且波动明显,而在5月至9月温度较高的季节,物种丰富度指数保持在较高的水平。该结果表明,丰富度指数反映了昆虫群落受季节变化影响明显。
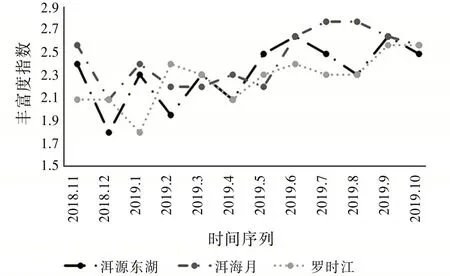
图7 丰富度指数变化趋势
同样明显受季节变化影响的还有昆虫数量。从图8可以发现:冬季温度较低,在2018年12月3个湿地的温度都达到全年的最低值。与此同时,3个湿地采集到的总昆虫数量也达到全年的最低值。而在4月之后,3个湿地均超过时间序列年平均温度,昆虫的数量开始增多。特别地,洱海月湿地采集的昆虫数量巨幅上升,这是因为采集到的蚂蚁数量增多。从2月的14头、3月的34头、4月的53头到5月的142头、6月的94头、7月的113头。可见,蚂蚁数量的激增是导致洱海月湿地昆虫群落参数变化的主要原因。
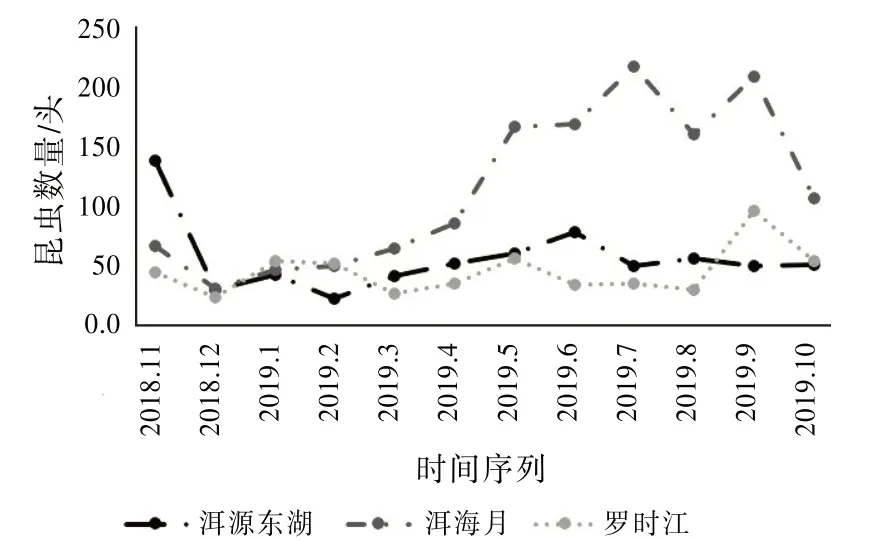
图8 昆虫数量变化趋势
由式(4)知,优势集中性指数和昆虫群落结构稳定性成反比。由图9发现:2019年5月洱海月湿地昆虫群落优势集中性指数达到最高0.729 0,这反映出该时间段内该区域出现了极端优势物种,也就是图8中提到的蚂蚁,此时洱海月昆虫群落稳定性最弱,正好对应图6该月均匀度指数达到最小值。同理不难发现:罗时江湿地在该时间段的优势集中性指数也达到全年最高0.598 2,其原因也是蚂蚁数量的激增,但是增长率相对于洱海月湿地要小。对应图6也反映了此时罗时江昆虫群落稳定性较弱。
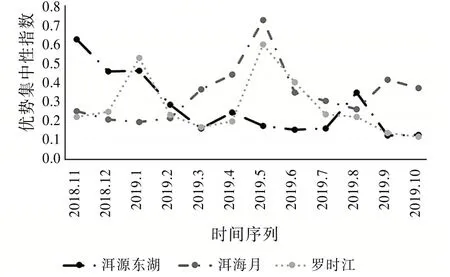
图9 优势集中性指数变化趋势
4.4 昆虫群落结构特征参数相关性分析 为进一步研究各个特征参数间的关系以及对昆虫群落稳定性及多样性的影响,这里引入相关系数R〔16〕,考察一年内各个特征参数时间序列间的关联程度。相关系数值介于-1和1之间,其中越靠近1说明正相关性越强,越靠近-1说明负相关性越强。得出的相关系数见表7。
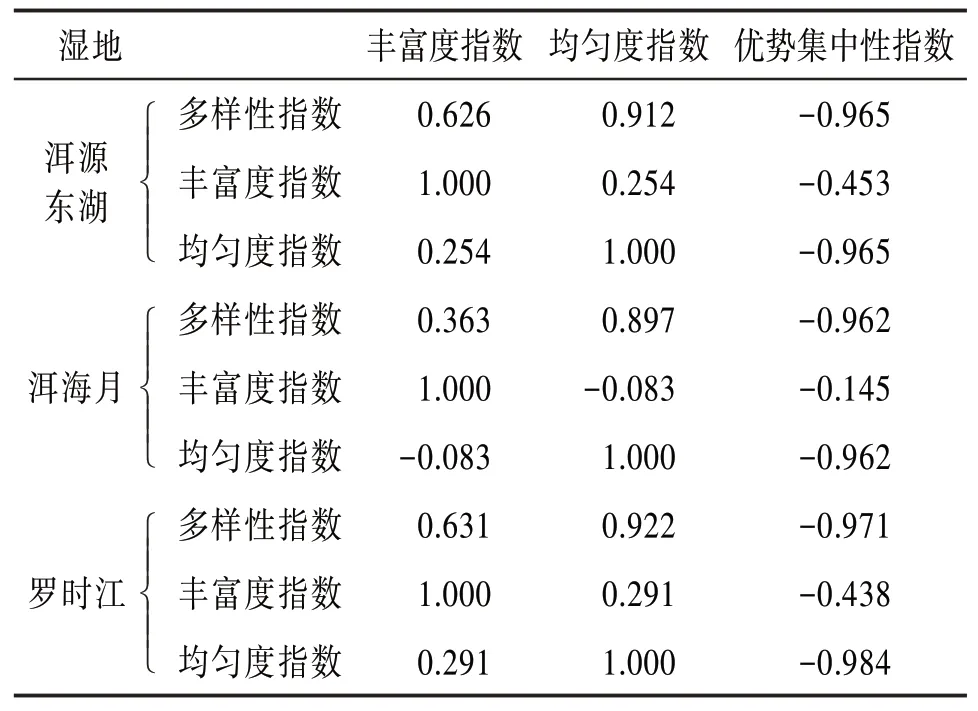
表7 特征参数相关系数表
由表7可见,3个湿地的多样性指数与均匀度指数的相关性均为0.9左右,具有较强的正相关性,说明洱海湿地昆虫群落的多样性指数与均匀度指数关系密切。而3个湿地的优势集中性指数分别与多样性指数和均匀度指数的相关性均小于-0.9,具有较强的负相关性。
综上分析,对于洱海湿地昆虫群落保护,首先要保证物种的丰富度,特别是稀有类群,稀有类群的多少直接决定了该湿地的昆虫生态网络的大小。较大的昆虫生态网络具有较好的鲁棒性。而优势类群则很大程度上限制了昆虫群落的多样性。所以洱海湿地昆虫群落的保护策略是:优先保护濒临消失的类群,重点监控优势类群的发展。
4.5 生境因子对昆虫群落结构特征参数影响分析根据3个湿地环境监测点采集的气象数据,分别求出昆虫采集前后3 d内的温度、环境湿度、紫外线瞬时值、降雨量、风速的平均值,并构建时间序列。考察一年内生境因子与昆虫群落结构特征参数的相关性。分别得出3个湿地生境因子与该湿地特征参数的相关系数的绝对值,并求出平均数。最后得到的相关系数见表8。
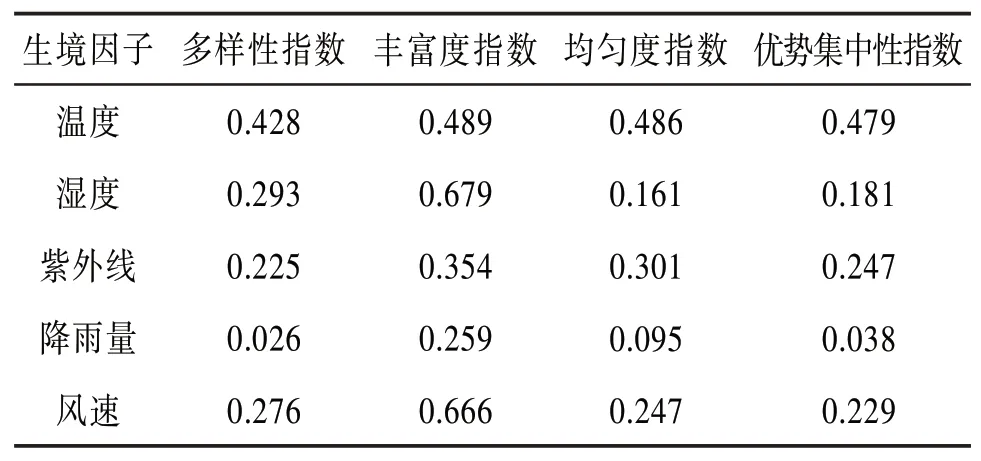
表8 生境因子与特征参数相关系数表
由表8可知,湿度和风速与丰富度指数的相关性达到了0.6以上,这可能与大理的地理因素有关,大理地处亚热带高原季风气候区,常年日照充足,水分蒸发量大,气候干燥;昼夜温差大,常有大风天气。其中,昆虫群落丰富度指数随着湿度的增加而降低,随着风速的降低而增加。湿度越大物种越丰富,而风速过大会导致飞行类昆虫的种类数量降低。
同时,温度与昆虫群落结构的特征参数更为密切,各相关系数都达到了0.4以上。图10展示了昆虫采集前后3 d内的日平均温度。
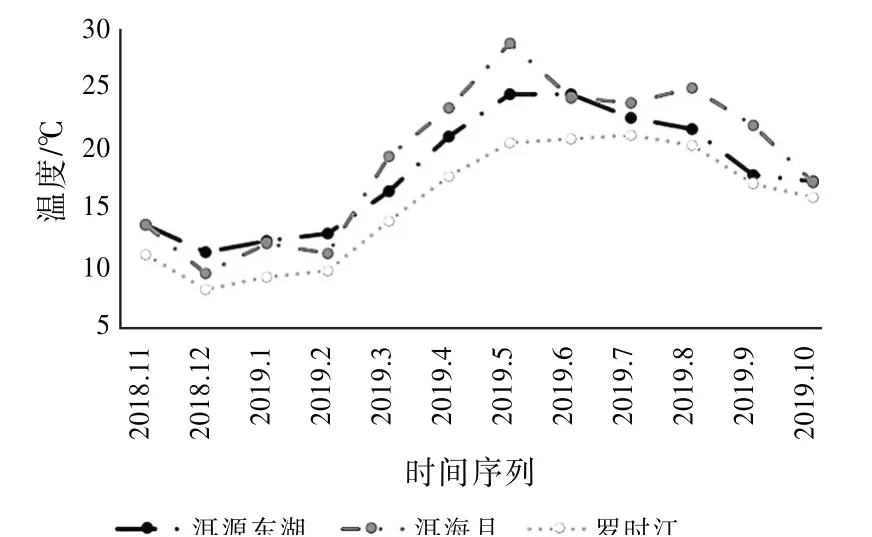
图10 湿地温度变化趋势
可以看出罗时江湿地的温度要低于另外两个湿地,洱源东湖湿地的温度在大部分时间介于洱海月湿地和罗时江湿地之间。洱源东湖湿地和罗时江湿地的温度变化趋势保持一致,温度变化曲线相对平缓,而洱海月湿地的温度变化较大,并且存在多个明显拐点,最明显的是2019年5月,平均温度达到最高的28.8℃,明显高于洱源东湖的24.5℃和罗时江的20.4℃。又因蚂蚁生长活动最佳的温度为25~35℃之间,所以洱海月湿地采集到的蚂蚁数量大于另外两个湿地,优势物种的出现对群落结构的稳定性产生较大的影响。而罗时江常年温度低于另外两个湿地,采集到的物种种类与物种数量都低于另外两个湿地。由此推测:洱源东湖湿地的温度更适中,全年温度变化更加平缓,更适合昆虫群落均衡发展。可见,温度是影响昆虫群落结构参数的主要因素。
4.6 昆虫群落稳定性分析 本文采用式(5)、(6)对昆虫群落的相对稳定性进行研究。分别对3个湿地每个月的稳定性指标进行计算并求取平均值,得到3个湿地的昆虫群落相对稳定性数值,见表9。
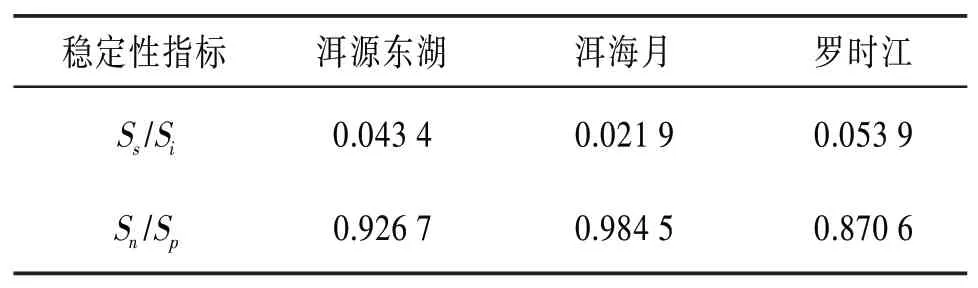
表9 昆虫群落的相对稳定性数值表
由表9可知S s/Si的大小顺序为罗时江>洱源东湖>洱海月,Sn/S p的大小顺序为洱海月>洱源东湖>罗时江。其中洱源东湖湿地的相对稳定性指标数值都位于第二,且接近第一,而另外两个湿地的稳定性指标数值都有偏小项。所以综合分析可知,洱源东湖湿地的昆虫群落网络内部关系较为复杂,稳定性最高。另外两个人为建设的湿地昆虫群落稳定性虽然低于洱源东湖,但水平接近洱源东湖湿地。
5 结论
本文通过昆虫群落结构及其特征参数研究,结果发现:对湿地昆虫群落影响较大的除了四季变化带来的温度等自然生境变化,就是人类活动对其产生的干扰。3个洱海湿地的昆虫群落结构具有相似性,物种较为丰富。其中,洱源东湖湿地的昆虫群落结构最稳定,其余两个湿地昆虫群落也具有较强的自我调节修复能力。洱海湿地的昆虫群落多样性指数、丰富度指数以及均匀度指数越大,优势集中性指数越小,反映出昆虫群落物种丰富,结构稳定,所监测的栖息地生态环境越好。
同时,洱海月湿地昆虫群落结构多样性表明,通过人为建设改造后的湿地,从长远来看并不一定会破坏湿地环境。相反,如果能够维持昆虫群落结构稳定性和自适应性,也能达到原生自然生境的生态标准。所以合理的建设规划对于洱海湿地保护很有必要。如果人类活动过于频繁,将导致极端优势物种的产生,势必将在一定程度上破坏昆虫群落的稳定性。所以合理地划分人类活动区域和原生湿地区域,对保护湿地生态环境具有重要意义。
