小麦PLT2基因的克隆及功能分析
2021-06-09刘会文宋少帅尹华燕贺小彦刘家斌林琪穆平
刘会文,宋少帅,尹华燕,贺小彦,刘家斌,林琪,穆平
(青岛农业大学农学院,山东 青岛 266109)
小麦是世界上的重要粮食作物之一,保持小麦高产、稳产对保障我国粮食安全和农业可持续发展具有重要意义。盐胁迫是自然界主要非生物逆境之一,严重影响小麦生长和发育[1]。盐胁迫对植株的危害主要表现在渗透胁迫、离子毒性和氧化胁迫[2]。土壤盐分过多导致细胞离子失衡引发渗透胁迫,而这种离子失衡又进而引发活性氧的产生,造成氧化胁迫[3]。
目前已发现的糖转运蛋白分为三类:单糖转运蛋白(monosaccharide transporters,MSTs)、蔗糖转运蛋白(sucrose transporters,SUTs)、SWEET蛋白(sugars will eventually be exported transporters)。SWEET蛋白为新发现的蛋白,含有MtN3/saliva保守结构域,属于MtN3家族。除SWEET外,大多数已鉴定的糖转运蛋白含有MFS(major facilitator superfamily)保守结构域,属于协助扩散超家族MFS中的糖转运子亚族[4-6]。该亚族蛋白具有疏水性的特点。研究表明,糖转运蛋白在维管束介导的糖运输以及应答多种逆境胁迫中发挥作用[7,8]。Miao等[9]研究发现,在高盐胁迫下油菜中BrSWEET11-LF和BrSWEET17-MF1的表达量均显著提高。Feng等[10]发现在高温、高盐、低温条件下番茄糖转运蛋白家族成员在叶片、根等组织中的表达量明显改变。拟南芥糖转运蛋白基因受低温、干旱和高盐胁迫的诱导表达[11-13]。
本研究基于抗旱耐盐小麦品种青麦6号转录组数据,筛选到一个盐胁迫下差异表达的糖转运蛋白基因序列,命名为TaPLT2。本研究拟通过对该基因的克隆与功能分析来探究小麦糖蛋白基因与耐盐性的关系,为小麦耐盐遗传改良提供新的基因资源及理论支撑。
1 材料与方法
1.1 试验材料
TaPLT2基因克隆及表达水平分析采用的试验材料为耐盐小麦青麦6号;基因多态性分析选用72份黄淮麦区冬小麦新品系。
1.2 TaPLT2的筛选
本研究根据实验室自存的青麦6号盐胁迫处理前后转录组数据(https://pan.baidu.com/s/1VEhmkf4n7qY7rw6JhG2tgA,提取码1234),在差异倍数以及显著水平两个层次上进行筛选,设定阈值为且q-value<0.005,筛选出不同处理时间点的差异表达基因。将所有差异表达基因按照差异表达倍数降序排列,并在NCBI网站上分别对差异表达倍数大的基因进行BLAST比对,筛掉前人研究过的基因,筛选出Traes_2AS_E158810C6基因,预测功能为糖转运蛋白基因,命名为TaPLT2,并进行下一步基因克隆及后续研究。
1.3 TaPLT2基因的克隆及测序
将青麦6号幼苗按照TaKaRa公司的RNA提取试剂盒、cDNA反转录试剂盒说明书提取RNA并反转为cDNA。采用Primer Premier V5.0软件进行引物设计,引物见表1中cPLT2F/R,由擎科生物股份有限公司合成。采用TaKaRa公司的高保真酶进行TaPLT2基因扩增。PCR反应程序为98℃预变性3 min;98℃变性10 s,71℃退火30 s,72℃延伸30 s,循环35次;72℃延伸10 min,12℃保存。扩增片段回收产物送擎科生物股份有限公司测序。将测序结果放入小麦全基因组数据库进行比对,分析该基因在染色体中的位置、内含子个数及拷贝序列数。
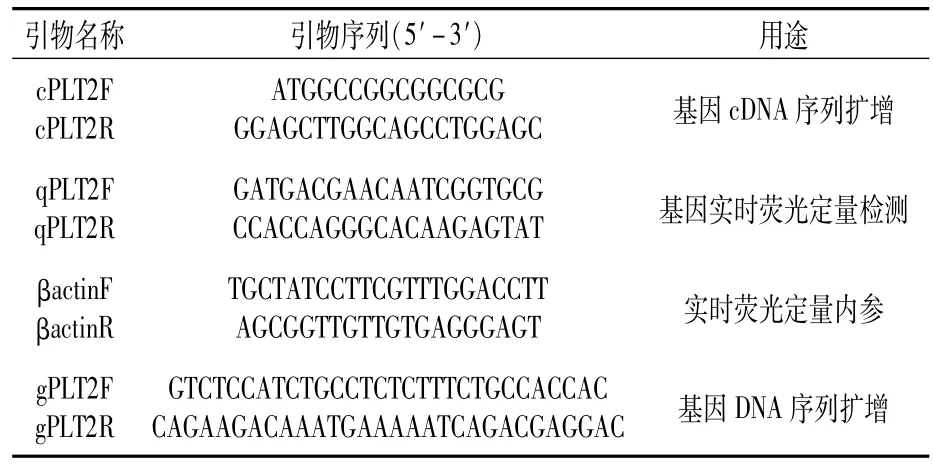
表1 所用引物
1.4 TaPLT2的生物信息学分析
根据克隆出的TaPLT2基因序列找到蛋白质编码区(CDS),利用BioXM2.6翻译成蛋白质。利用ExPASy网站在线工具ProtParam(https://web.expasy.org/protparam/)进行蛋白基本理化特性分析,包括氨基酸组成、相对分子量和理论pI值等;利用PROTCOM网站(http://www.softberry.com/berry.phtml?topic=protcomppl&group=programs&subgroup=proloc)进行蛋白质的亚细胞定位;利用ProSWEETale工具(http://ca.expasy.org/tools/proSWEETale.html)进行蛋白质疏水性分析;利用TMpred工具(http://www.cbs.dtu.dk/services/TMHMM-2.0/)进行跨膜区和跨膜方向预测;通过ExPASy Proteomics Server系统中的软件在线预测其功能性位点;利用CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)进行蛋白质保守结构域分析;通过NCBI网站进行BLASTP比对,下载相似度较高的序列,结合MEGA 10.0对下载的序列构建NJ系统进化树。
1.5 TaPLT2的表达模式分析
取青麦6号种子50粒,先用蒸馏水清洗干净,再用75%酒精消毒5 min,最后用蒸馏水清洗至无酒精异味。将处理完成的小麦种子均匀平放在铺有两层湿润滤纸的培养皿中,置于光照培养箱中培养。培养条件:光/暗为16 h/8 h,昼夜温度为25℃/19℃。培养至两叶一心期,用200 mmol/L的NaCl溶液胁迫处理0、24、48、72 h,3次重复;剪取胁迫处理后的小麦幼苗,液氮速冻,存放于-80℃超低温冰箱中备用。用引物qPLT2F/R、βactinF/R(表1),采用TaKaRa公司的实时荧光定量试剂盒进行TaPLT2基因的表达模式分析,3次生物学重复。qRT-PCR反应程序:95℃预变性30 s;95℃变性5 s,57℃退火延伸30 s,同时在退火过程中检测荧光信号变化,共进行39个循环。
1.6 TaPLT2的多态性分析
挑选72份小麦材料饱满一致的种子各60粒,消毒处理后于培养箱中培养至一叶一心时,剪取叶片,用CTAB法分别提取基因组DNA。采用引物gPLT2F/R(表1)对各品种TaPLT2基因组DNA序列进行扩增,测序由擎科生物股份有限公司完成。PCR反应程序为94℃预变性4 min;98℃变性10 s,60℃退火30 s,68℃延伸3 min,循环35次;68℃延伸10 min,12℃保存。基因多态性用DNAMAN软件进行比对分析。
当小麦长到两叶一心时,用200 mmol/L的NaCl溶液胁迫处理3、5 d后分别取样,用刻度尺测量小麦根长、株高;用蒸馏水分别将小麦幼苗冲洗干净,吸水纸吸干后装入牛皮袋,于烘箱内105℃杀青30 min后70℃烘干至恒重,测量干重;将烘干样本置于组织破碎机中打成粉末,用1/10000天平称取地上部约0.0500 g、根系约0.0200 g于消煮管中,加入5 mL HNO3,180℃消煮炉上恒温消解60 min直至颜色消失,冷却定容,过滤后用火焰光度计测量K+、Na+含量;参照蒽酮比色法[14]测定可溶性糖,称取0.3 g样品放入大试管中,加入15 mL蒸馏水,沸水浴20 min,冷却,过滤入100 mL容量瓶中,用蒸馏水冲洗残渣数次,定容至刻度。取提取液1.0 mL加蒽酮试剂5 mL,在沸水浴中煮10 min,取出冷却,在620 nm波长下测定光密度,3次重复;从标准曲线上查出每毫升被测样品液可溶性糖的含量,再计算出鲜样中可溶性糖的平均值。
用SPSS和Microsoft Excel进行相关性分析。不同品种的耐盐性差异采用各性状指标的相对值进行比较,用盐胁迫处理的测定值与对照组的测定值比较得到相对值。SNP位点与耐盐性状指标相关系数大于0.3表明有相关性[15]。为了消除单个指标带来的片面性,用隶属函数法[16,17]将各小麦材料的各耐盐指标扩展到[0,1]闭区间内,用下式分别计算各耐盐指标的隶属函数值:
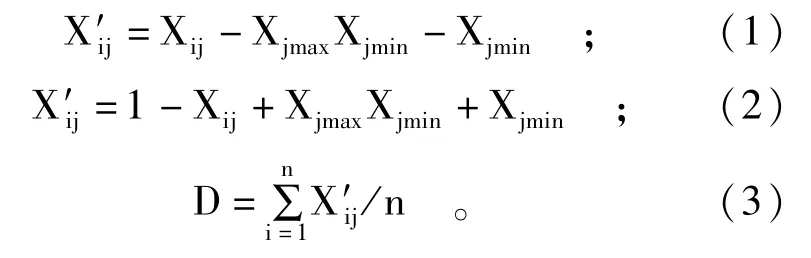
2 结果与分析
2.1 TaPLT2基因的克隆
以青麦6号cDNA为模板,利用引物组合cPLT2F/R(表1)进行PCR扩增,得到大小为1 300 bp左右的单一条带(图1)。将目的片段回收、测序后,得到长度为1 385 bp的序列,开放阅读框为1 155 bp,编码385个氨基酸,将该基因命名为TaPLT2。
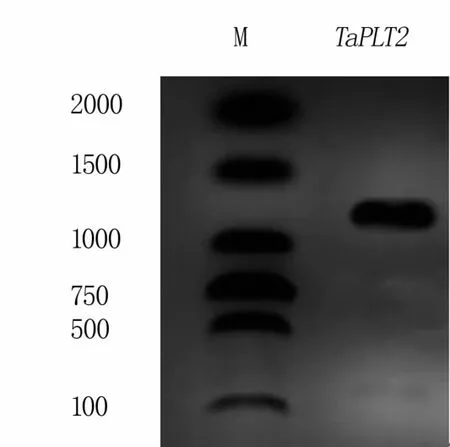
图1 TaPLT2的扩增
2.2 TaPLT2生物信息学分析
分析表明,TaPLT2定位于2AS染色体,基因组序列全长2 194 bp,存在2个内含子,在小麦染色体中为单拷贝。TaPLT2编码385个氨基酸,丙氨酸(Ala)和亮氨酸(Leu)是占比最高的两种氨基酸,分别达到10.0%和10.2%;负电荷残基总数(Asp+Glu)为27,正电荷残基总数(Arg+Lys)为33,相对分子质量42 269.99 Da,理论等电点为8.99。TaPLT2多肽链第250位的具有最高分值4.500,疏水性最强;第138位的具有最低分值-4.500,亲水性最强,整条多肽链表现为疏水性(图2A),因此可推断TaPLT2是一种疏水性蛋白。其定位于细胞质,有7个跨膜结构域(图2B),分析该多肽链的功能性位点发现多肽链含有糖转运蛋白功能位点Sugar transport proteins signature 1(图3)。
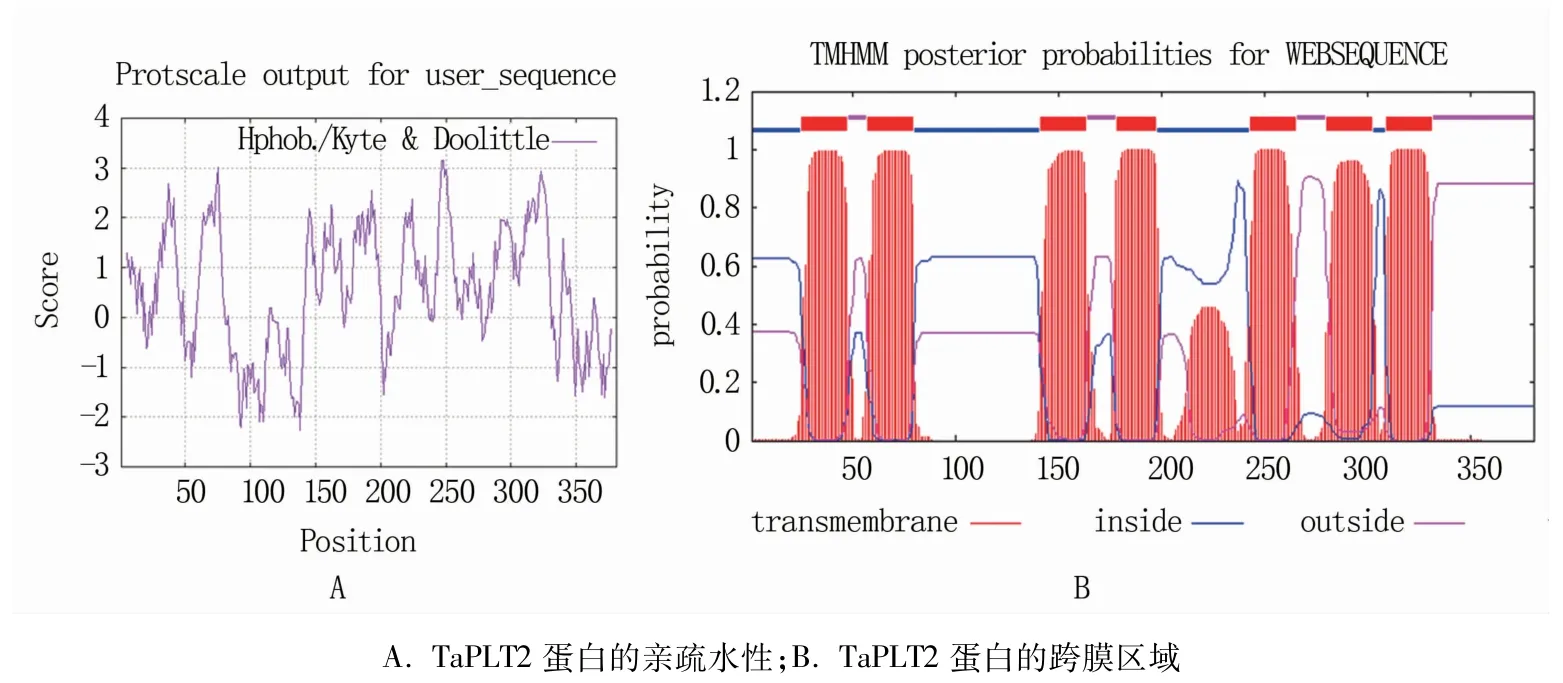
图2 TaPLT2蛋白的亲疏水性和跨膜区域
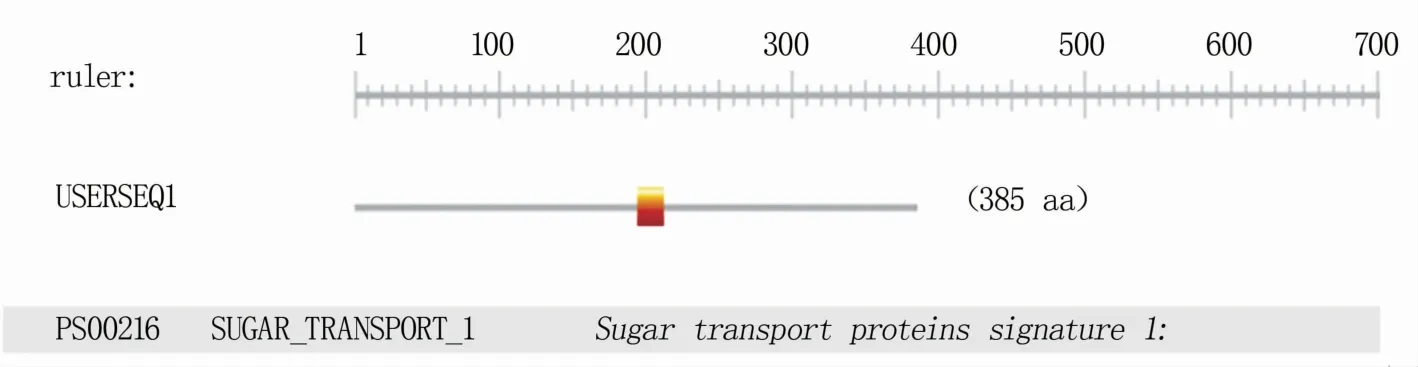
图3 TaPLT2蛋白的功能性位点分析
将TaPLT2编码的氨基酸序列在NCBI数据库中进行BLASTP对比,结果表明其与节节麦(XM_020325583.2)、小苔草(SWLB_01000017.1)、青 稞 (SDOW_01000136.1)、大 麦 (AK_363174.1)、高粱(XM_002462974.2)、兰花(KZ_453008.1)、香蕉(PYDT_01000002.1)、Ensete ventricosum(AMZH_03009594.1)的糖转运载体蛋白同源。利用DNAMAN软件将TaPLT2氨基酸序列和其他物种的氨基酸序列进行多序列比对,结果显示,该蛋白与其他物种蛋白的相似度为79%~99%,且都含有MFS_STP结构域,属于MFS家族的糖转运子亚族(图3—图5)。

图4 TaPLT2与其他物种氨基酸序列的多重比对
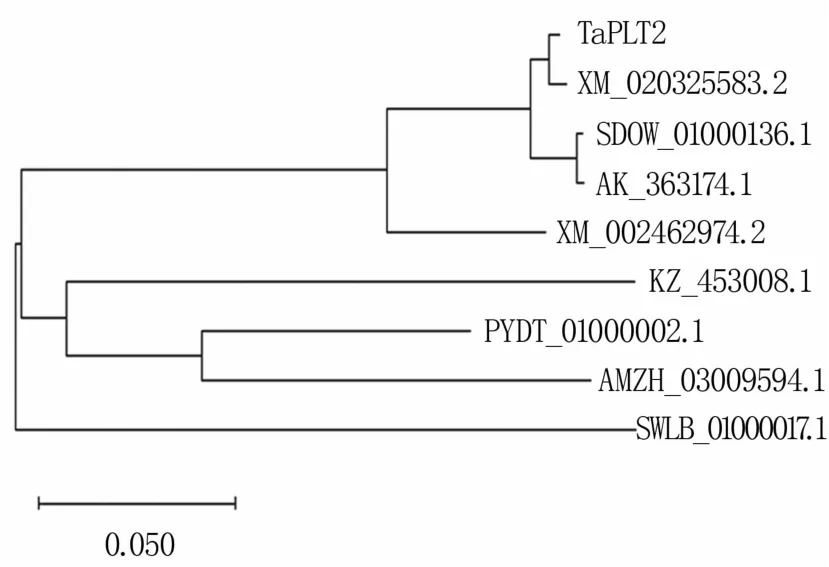
图5 糖转运蛋白系统进化树
2.3 TaPLT2的表达模式分析
以不同处理青麦6号总RNA反转录的cDNA为模板进行实时荧光定量分析,结果显示,TaPLT2的表达整体呈先升高后持平趋势,表达量在盐胁迫24 h时最高,为对照组的3.2倍,48 h及72 h时相对表达量大体保持一致(图6)。表明TaPLT2基因受到盐胁迫诱导。
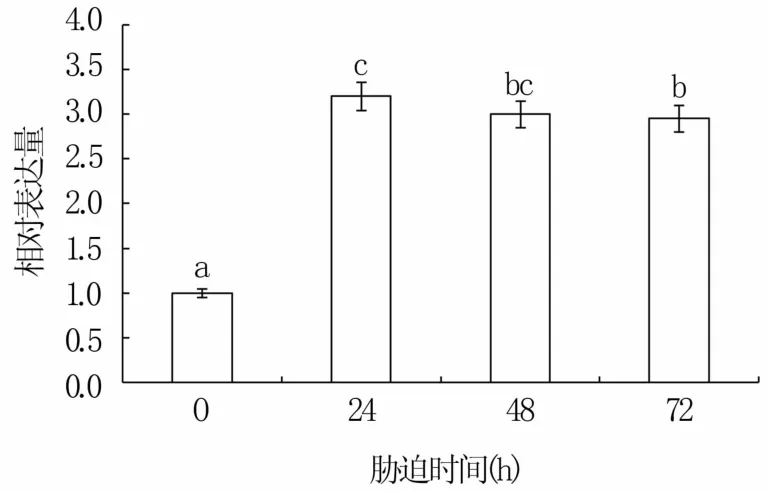
图6 TaPLT2基因的表达情况
2.4 TaPLT2序列多态性分析
分别以72个小麦品系的基因组DNA为模板,以表1中A基因组特异引物gPLT2F/R扩增TaPLT2基因组全长序列(外显子+内含子)。经测序及比对分析(图7),72份材料存在一个SNP多态性位点,命名为TaPLT2-SNP1,位于TaPLT2基因1 326位,即基因第3个外显子上存在T/C两种多态性,氨基酸存在苏氨酸/丙氨酸两种情况,突变率为9.46%,通过对蛋白质亲疏水性进行分析,该突变使基因亲水性增加0.006。
2.5 TaPLT2-SNP1与耐盐相关表型的关联分析
由表2可以看出,盐胁迫处理3 d和5 d的小麦根长、株高及干重显著低于对照组;从渗透调节物质变化水平来看,盐胁迫3 d的K+含量显著低于对照,盐胁迫5 d的K+含量与对照差异不显著;盐胁迫处理下的Na+含量及可溶性糖含量均显著高于对照组。
从表3可以看出,C变异与根长呈显著负相关,并且随着盐处理时间的延长,相关性增大;C变异与株高无显著相关性;盐胁迫3 d时C变异与植株Na+呈显著正相关,盐胁迫5 d时C变异与植株K+呈显著负相关;C变异与单株干重无显著相关性,但相关系数随处理时间增加而增大;C变异与可溶性糖含量呈极显著正相关,与D值呈显著正相关。
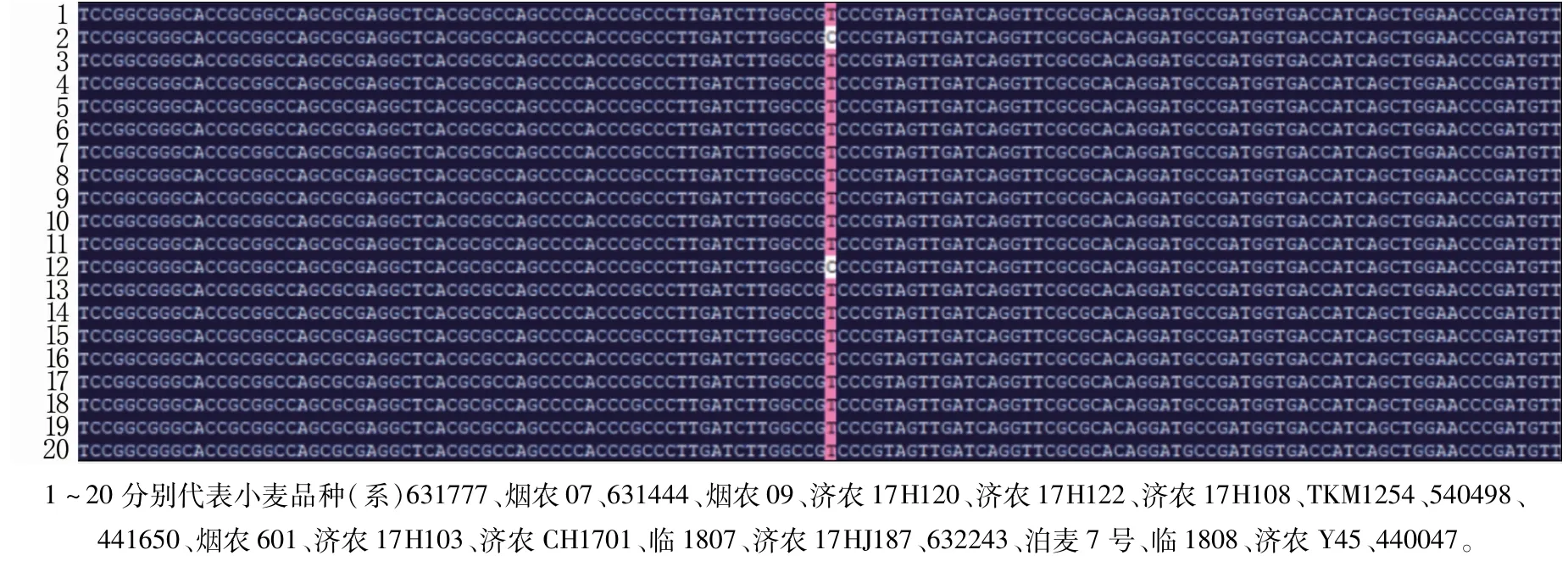
图7 部分小麦品系TaPLT2基因SNP位点
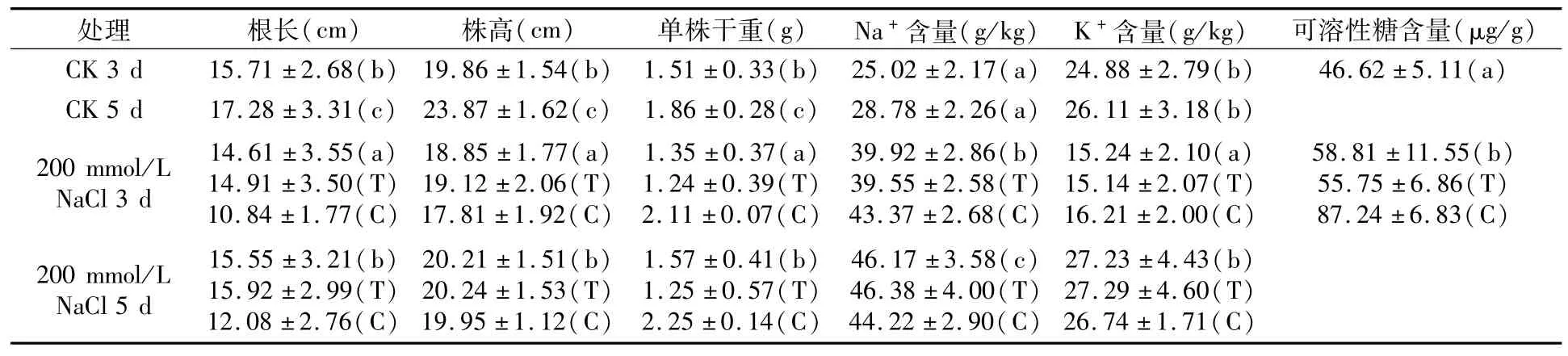
表2 盐胁迫对小麦相关性状影响

表3 TaPLT2基因C变异与耐盐性的相关关系
3 讨论与结论
糖转运蛋白在植物体中广泛存在[18],而小麦糖转运蛋白基因在耐盐方面的研究相对较少。本研究通过RT-PCR技术首次克隆出小麦TaPLT2基因的全长cDNA,cDNA全长为1 385 bp,开放阅读框为1 155 bp,编码385个氨基酸。通过对编码氨基酸进行序列分析表明TaPLT2含有糖转运功能部位且与节节麦、大麦、高粱的糖转运蛋白序列高度同源,都含有MFS_STP结构域,属于MFS家族的糖转运子亚族[19]。生物信息学分析显示该蛋白具有疏水性,定位于细胞质且具有7个跨膜结构域。
研究表明,许多糖转运蛋白基因家族成员都能响应盐胁迫[20-23]。本研究结果显示,TaPLT2基因受盐胁迫诱导上调表达,在胁迫24 h后表达量达到最高,为对照组的3.2倍,这与Seo等[12]的研究结果一致,表明TaPLT2基因参与盐胁迫的调控。高盐分环境影响植物正常的新陈代谢和各项生理活动[24-27]。本研究同样发现高盐胁迫阻碍小麦植株根长及株高的伸长、干重的增加,提高小麦植株的可溶性糖含量及Na+含量;盐胁迫处理3 d的小麦植株K+含量下降,而胁迫处理5 d的K+含量与对照无异。这表明高盐胁迫会阻碍小麦根长及株高的伸长、干物质的积累,提高植株Na+及可溶性糖含量,影响植株K+含量。
相关研究表明,植物耐盐性与根长[28]、株高[29]、干重[30]、K+、Na+含量[31]和可溶性糖含量[32]存在相关性。D值作为通过隶属函数法对所测性状进行综合分析所得的数据能避免单一性状的局限性[15]。耐盐基因多态性会对该耐盐基因的耐盐能力产生影响[33]。对TaPLT2基因在72个小麦品种中的序列多态性进行分析,发现TaPLT2基因的第1 326位即第3个外显子处存在T/C两种多态性,氨基酸存在苏氨酸/丙氨酸两种变异,72个品种的突变率为9.46%。通过相关性分析发现,C变异与根长呈显著负相关,与可溶性糖含量呈极显著正相关,推测C变异通过增加可溶性糖含量提高细胞渗透势的方式来增强耐盐性。C变异与D值呈显著正相关,推测该多态性位点能够影响TaPLT2基因的耐盐性。
