现场模拟光照条件对南极大磷虾冬季耗氧率的影响研究
2021-05-12胡光照刘子俊朱国平
胡光照,刘子俊,朱国平,3,4
(1.上海海洋大学海洋科学学院,上海 201306;2.上海海洋大学极地研究中心,上海 201306;3.大洋渔业资源可持续开发教育部重点实验室极地海洋生态系统研究室,上海201306;4.国家远洋渔业工程技术研究中心,上海 210306)
南极大磷虾(Euphausiasuperba,以下简称磷虾)为磷虾目大型海洋甲壳类浮游动物。该种在南大洋生态系统中起核心作用,被认为是南大洋生态系统的关键物种,是连接初级生产者和脊椎动物捕食者之间的主要生物[1]。在南大洋大部分地区被冰覆盖、水体中浮游植物含量极低的冬季,磷虾如何生存是南大洋海洋生态系统动力学计划(SO GLOBEC)研究的关键问题之一[2],也一直是全球极地生态学家极为感兴趣的重要科学问题。总的来讲,成体磷虾的越冬机制通常可分为两类:一类是非捕食机制,包括使用体内已储存的脂质[3],降低体内的代谢率[4-6]及逆生长[7]等;另一类则是改变捕食策略以及转换食物来源,即由夏季的主要摄食对象(如硅藻等浮游植物)转变为冰藻以及海冰底下的小型浮游动物等[8-9]和海底碎屑[4]。其中,降低代谢率被认为是成体磷虾最有效的冬季节能机制之一[5,9]。与夏季相比,冬季生物体内代谢关键酶——柠檬酸合成酶(CS)的活性显著降低,这也导致磷虾的CS活性和核糖核酸∶脱氧核糖核酸(RNA∶DNA)比值出现明显的季节性变化[10]。代谢率的降低可能是冬季磷虾的主要生理反应,但导致这种降低的机制仍不清楚。有报道[11]认为,光周期的变化是磷虾代谢率季节性变化的主要诱因之一。
为探究光周期对磷虾代谢周期的影响,一些学者通过不同方法进行了研究。TESCHKE等[11]通过陆基暂养实验发现,光照强度会使磷虾的耗氧率升高并提升代谢酶苹果酸脱氢酶(MDH)的活性,验证了环境光照状态的变化对磷虾的生理参数(如摄食和代谢率)变化具有重要影响。BROWN等[12]的实验却得出了相反的结论,其结果显示,黑暗条件下的磷虾耗氧率高于处于光照条件下的磷虾,可能是由于黑暗条件下磷虾更活跃,游泳行为增多,由此使得耗氧率增加。上述研究均说明了光照条件会影响磷虾的行为和生理。总的来讲,有关冬季磷虾生理的现场研究仍较为有限。我国学者通过现场实验对磷虾的行为和耐受性等进行了研究[13-16],并分析了光照条件对磷虾垂直分布的影响[17-19],但尚未有研究涉及到生理。因此,通过现场实验了解光照变化对磷虾生理的影响很有必要。有氧代谢时释放的热量与消耗的氧量呈正比,因此在海洋生物生理代谢研究中,耗氧率可直接作为衡量代谢率高低的一个指标[20]。鉴于此,本研究通过海上现场暂养实验就不同光照条件对磷虾代谢和生理状态的影响进行分析,以期了解冬季磷虾代谢率对光照条件变化产生的反馈,为揭示磷虾越冬机制提供基础数据。
1 材料与方法
1.1 实验设置
实验所用磷虾活体由大型拖网渔船“龙腾”从事渔业生产过程中取得,采集时间为2019年6月20—22日,取样地点为南设德兰群岛水域(62°33′S、59°28′W),采样海域海水温度为-1.8℃,水深为800~1 000 m,虾群深度为50~140 m,空气温度为-11℃。采集到的活体磷虾暂养于100 L的水槽中,暂养水体为现场采集的自然海水(水温为-1.2℃~0.5℃)。暂养箱放置于平均室温为0℃的隔离空间内,光照强度为90~150 lx(与所在海域自然光照强度基本相符)(图1)。暂养期间(4~6 d)未投饵料,暂养箱每12 h换水一次,每次换水4/5,并将箱底的死亡个体、虾壳和粪便等由底部排水口排出。至6月26日,水箱内每日死亡个体的数量趋于稳定(5尾),剩余磷虾个体状态保持良好,随机选取活性较好的9尾磷虾进行光照对比实验。
1.2 实验过程
实验开始于2019年6月26日23∶00。将9尾磷虾分成2组,A组5尾磷虾,B组4尾磷虾(在实验进行到第3天时,磷虾编号2死亡,故加入样本9)(表1)。所有实验个体分别置于容积为630 mL并注入当地海水的白色塑料瓶内后封口。封口后的实验塑料瓶放入装有当地循环海水的100 L箱体内,使得实验个体所处的温度始终与当地海水保持一致。实验期间,使用温盐深仪(CTD,型号为RBR)对每次实验的海水温度和盐度进行记录,海水盐度在32.878~32.968之间,较为稳定。使用TES-1322A型电子照度计测量实验场地光照强度,并对实验期间的照度进行调控,以保持光照强度尽量恒定,暂养水箱水体表面光照强度约为100 lx。除光照条件外,实验期间的其他各项环境数据均相同。直至实验完成,剩余所有磷虾个体均保持较好活性。

图1 南极大磷虾暂养装置示意图Fig.1 Schematic map of aquarium tank for Euphausia superba

表1 实验南极大磷虾样本基本信息Tab.1 Basic information about Euphausia superba specimens used in this study
针对A组个体,在光照条件下放置约24 h后,使用溶解氧检测仪(proODO,YSI 626281)测量瓶内海水溶解氧浓度(OT,精度0.01 mg·L-1);测完后,采用锡箔纸将试验瓶完全密封包裹,以进行黑暗条件下的实验。再经过约24 h后,测量试验瓶内水体的溶解氧浓度(OT+1);测完后,将A组实验个体置于同等光照条件下,以进行光照条件下的实验。以此类推,A组实验共持续11 d(6次光照条件和5次黑暗条件)。
针对B组个体,采用锡箔纸将试验瓶完全密封包裹,以进行黑暗条件下的实验。密封后的试验瓶放置约24 h后,使用溶解氧检测仪测量瓶内海水溶解氧浓度(OT);测完后,将试验瓶放置在光照强度为100 lx的光照条件下。再经过约24 h后,测量试验瓶内水体的溶解氧浓度(OT+1)。完成前2 d的实验后,B组实验与A组保持相同条件同步进行,直至实验结束。最终,B组实验也持续共11 d(6次光照条件和5次黑暗条件)。
上述两组实验均设置空白对照组,即在试验瓶中不放置磷虾,经同时段后测定实验开始时水体的溶解氧浓度(O′T)以及结束时背景溶解氧浓度(O′T+1)。实验结束后,将所有9尾磷虾样本收集,置于-20℃冷冻条件保存。待运回陆基实验室后转移至-80℃超低温冰箱内保存。
1.3 样本处理
返回实验室后,将置于-80℃超低温冰箱的磷虾样本取出并于常温解冻。测量解冻后的磷虾标准体长(total length,TL;mm)、湿体质量(wet mass,MW;mg)和 性 成 熟 度。标 准 体 长 据MAUCHLINE[23]的标准,即由眼最前端至尾节末端之间的长度。性成熟度的划分结合MAKAROV和DENYS[21]与BARGMANN[22]的划分标准进行判定,文中的标准也修改为未成体(雌性分为2BF、2CF、2DF 3期,雄性分为2AM、2BM、2CM 3期)、成体(雄性分为3AM、3BM两期,雌性分为3AF、3BF、3CF、3DF、3EF 5期)。本实验中的9尾磷虾均为未成体。测量后的磷虾个体在实验室冷冻干燥后利用微量天平(MITTLER TOLEDO XPR2)称取其干重(dry weight,DW;mg)。
1.4 数据分析
耗氧率(respiration rate)指动物单位时间的耗氧量,可分为单位体质量耗氧率(weight specific respiration rate)和单位个体耗氧率(individual respiration rate)[20]。因此,每尾磷虾在实验中的单位干重耗氧率RT+1[μL·(mg·h)-1]及单位个体耗氧率RdT+1(μL·h-1)的估算方法分别为:

式(1)、式(2)中,ρ为氧气密度;V为实验暂养瓶的容积;W为实验磷虾经干燥后的体质量(mg);t为实验所进行的时间(h);OT为试验组开始时暂养瓶内海水的含氧量浓度(mg·L-1);OT+1为试验组结束时暂养瓶内海水的含氧量浓度(mg·L-1);O′T为空白对照组开始时暂养瓶内海水的含氧量浓度(mg·L-1);O′T+1为空白对照组结束时暂养瓶内海水的含氧量浓度(mg·L-1)。
为了分析各尾磷虾耗氧率随时间的变化是否存在显著性差异,本研究首先利用Shapiro-Wilk检验分析每尾磷虾耗氧率数据是否符合正态分布。如符合,则利用单因子方差分析(ANOVA)分析各尾磷虾耗氧率之间是否存在显著性差异;如不符合,则利用Kolmogorov-Smirnov test(K-S检验)分析数据。若方差齐次,利用LSD事后检验进一步分析组内两两个体之间是否存在显著性差异;若方差非齐次,则利用Tamhane T2事后检验分析组内两两个体之间是否存在显著性差异。显著性水平P=0.05。统计分析利用R4.0.0进行,所使用的程序包包括ggplot 2和MASS。模拟单位个体呼吸率随实验天数变化时,考虑到异常值存在,本研究使用了稳健线性模拟方法,以消除异常值对趋势的影响。
2 结果与分析
2.1 光照与黑暗条件对耗氧率的影响
磷虾单位个体耗氧率处于16.13~74.54 μL·h-1之间,平均为(29.460±9.343)μL·h-1(表2),其中黑暗条件下的磷虾单位个体耗氧率处于16.13~46.55μL·h-1之间,平均为(28.93±7.28)μL·h-1,而光照条件下的磷虾单位个体耗氧率处于17.76~74.54μL·h-1之间,平均为(29.89±10.83)μL·h-1。对于实验中磷虾单位体质量耗氧率,所有个体处于0.102 6~0.557 5μL·(mg·h)-1之间,平均为(0.269 0±0.078 4)μL·(mg·h)-1(表3);其中黑暗条件下的磷虾单位体质量耗氧率处于0.102 6~0.557 5μL·(mg·h)-1之间,平均为(0.267 3±0.084 0)μL·(mg·h)-1,而光照条件下的磷虾单位体质量耗氧率处于0.128 6~0.466 4μL·(mg·h)-1之 间,平 均 为(0.270 5±0.074 2)μL·(mg·h)-1。
从整体上来讲,磷虾单位体质量耗氧率与体长之间无相关性(R2=0.176,n=9,P=0.261)。就单位个体耗氧率而言,随着实验天数的增加,A组磷虾的个体耗氧率先下降再保持稳定,而B组磷虾的个体耗氧率呈增加趋势(图2)。

表2 南极大磷虾单位个体耗氧率的平均值、最小值和最大值Tab.2 Average,m inimum and maximum values of individual respiration rate of Euphausia superba
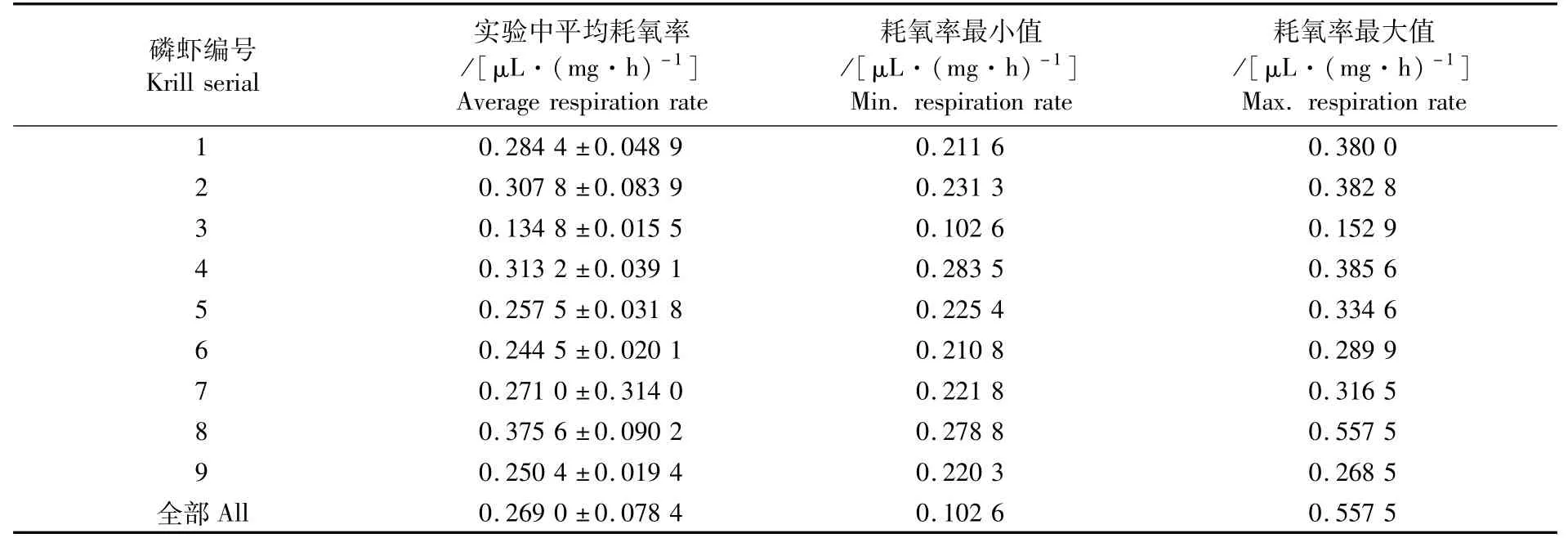
表3 南极大磷虾单位体质量耗氧率的平均值、最小值和最大值Tab.3 Average,m inimum and maximum values of weight specific respiration rate of Euphausia superba
2.2 两组磷虾耗氧率的个体差异
整体上来看,A组磷虾单位个体耗氧率变化较B组更为显著(图3)。A组中实验个体2在死亡前的单位个体耗氧率出现剧烈的波动。A组中,ANOVA结果显示,个体1、3和4的单位个体耗氧率存在显著性差异(F=25.478,df=32,P<0.001);LSD检验也显示,两两个体间也存在着显著性差异(个体1与个体3:P<0.001;个体1与个体4:P<0.001;个体3与个体4:P=0.022<0.05)。B组中,ANOVA结果显示,4个实验个体的单位个体耗氧率之间存在显著性差异(F=14.090,df=43,P<0.001);Tamhane T2事后检验显示,两两个体间并非均存在显著性差异,实验个体5和8(P=0.865)、6和7(P=0.839)以及7与8(P=0.119)之间并无显著性差异(表4)。

图2 各组南极大磷虾单位个体耗氧率随天数的变化Fig.2 Variations in individual respiration rate of Euphausia superba w ith days in two groups

图3 光照(黑暗)条件下南极大磷虾单位个体耗氧率变化Fig.3 Variations in individual respiration rate of Euphausia superba
如仅考虑同等实验条件,从加入9号个体后开始,ANOVA结果显示,8组实验磷虾(除去2号虾)的单位个体耗氧率仍存在显著性差异(F=20.134,df=48,P<0.001),但Tamhane T2事后检验显示,A组的个体1、3和4之间的单位个体耗氧率存在显著性差异,而B组中的个体6、7以及8之间的单位个体耗氧率则不存在显著性差异。
3 讨论
本次实验基于海上暂养实验完成了现场光照条件对磷虾冬季耗氧率的影响研究。在此之前,针对冬季磷虾耗氧率的研究极为有限(表5),
且仅有的研究多在陆基实验室内完成。为了尽可能地模拟实际情况,本研究模拟了磷虾自然条件下的生理状态,实验所使用的密闭瓶浸没于当地循环海水中,以保证实验环境水温与当地海水完全一致,这在一定程度上可以保证实验环境更接近磷虾生活的自然环境。

表4 B组实验磷虾单位个体耗氧率两两差异的Tamhane T2事后检验Tab.4 Tamhane T2 post hoc test for exam ining individual respiration rate of Euphausia superba in group B

表5 光照影响磷虾耗氧率研究Tab.5 Effect of light on respiration rate of Euphausia superba
3.1 磷虾冬季耗氧率
高密度的磷虾分布海域位于南极大西洋扇区的威德尔海、南奥克尼群岛、南设得兰群岛周围以及南桑维奇群岛西部水域[26],这些水域的水温、饵料、海冰覆盖情况及光周期和光强度[27]均存在着周期性的年际变化。环境特征的强烈季节性变化意味着磷虾的生长与发育也呈现出相应的季节性特征。但作为南大洋生态系统中的关键种,磷虾已进化出相应的生命周期以适应这种高度季节性的环境。冬季,磷虾通过降低代谢速率、缩小体型以及改变摄食策略等方法,由夏季的高代谢、高消耗生存模式向冬季的低代谢、低消耗模式转化,这种季节性生命周期变化使得磷虾能够在一年中较好地分配新陈代谢能力,即在食物源丰富且自然环境有利的夏季进行消耗较大的繁殖过程,而在条件极为苛刻的冬季转换为节能模式[28]。耗氧率作为衡量代谢率的一个重要指标,其变化可以较好地反映磷虾体内新陈代谢水平。
本研究结果显示,冬季黑暗条件下磷虾的单位体质量耗氧率平均为(0.267 3±0.084 0)μL·(mg·h)-1,这明显低于夏季黑暗条件下的单位体质量耗氧率0.63~0.88μL·(mg·h)-1[24],也低于实验室模拟的冬季黑暗条件下的单位体质量耗氧率0.35μL·(mg·h)-1和夏初黑暗条件下的单位体质量耗氧率0.56μL·(mg·h)-1[12]。而对于自然光照条件,本实验结果显示,磷虾的单位体质量耗氧率平均为(0.270 4±0.074 0)μL·(mg·h)-1,这略高于实验室模拟的冬季自然光照条件下的单位体质量耗氧率0.22μL·(mg·h)-1,但却低于夏初黑暗条件下的单位体质量耗氧率0.46μL·(mg·h)-1[12]。针对单位个体呼吸率,本研究结果与SWADLING等[25]类似。由此,可以初步判断,冬季磷虾的单位体质量耗氧率较夏季低,也进一步说明冬季磷虾的新陈代谢率有所下降[29]。但需要注意的是,实验室模拟条件下所获得的相关数据与自然条件下仍有所差别,造成该结果的可能原因之一在于磷虾的环境适应能力。经过长期进化,磷虾已适应并反馈南极特有的季节性光周期变化。正因为如此,通常在南极现场开展磷虾生长率等方面的实验时,仅设定较短的时间尺度(如4 d)[5],以防止磷虾因适应新的环境而在生理上做出改变。
3.2 光照对冬季磷虾耗氧率的影响
磷虾耗氧率存在较为显著的季节性变化(表5),而引起这种季节性变化的机制仍未明确。ATKINSON和SNYDER[30]就食物是否影响不同季节磷虾代谢速率开展了研究,通过将夏季磷虾分组暂养并以食物浓度作为条件变量检验两组磷虾耗氧率的差异,结果显示处于高浓度食物环境的磷虾耗氧率和清除率迅速上升。但针对秋季的实验却得到了不同的结果[31],后者将磷虾放置于食物充足的环境11 d,耗氧率并未发生显著升高,且耗氧率仅为夏季的三分之一。这两个实验在一定程度上说明食物浓度并非导致磷虾代谢率变化的主要原因。
南大洋日照时长具有明显的季节性差异,在磷虾的主要分布纬度(50~60°S),日照时长由12月的20 h转变至6月的6 h,这被认为可能是调节磷虾代谢状态的环境信号。TESCHKE等[11]发现,4—7月磷虾的耗氧率的确受到了光照条件的影响,处于光照条件下的磷虾耗氧率明显高于处于黑暗条件下的磷虾。这表明光照强度高时,磷虾耗氧率也会升高,并由此推断光照强度及其持续时间的年度过程可为控制磷虾的生理过程提供可靠且可预测的信息。这与KAWAGUCHI等[32]的研究结论相同,其认为在低水平的食物条件下,磷虾耗氧率的增加与太阳辐射增加有关。然而,BROWN等[12]的实验却得到了相反的结论,即黑暗条件下的磷虾耗氧率要高于光照条件,而该文作者分析认为实验水箱过大(100 L、600 L、1 600 L)是造成这种差异的原因,即冬季磷虾因已适应黑暗环境而增加游泳活动,其耗氧率也因活动量增加而高于夏季(光照组)磷虾。由此,为了降低因游泳活动增加而带来的影响,本研究使用了体积较小的实验瓶(500 mL)作为容器,这减少了游泳行为对耗氧率结果带来的影响,但结果并未显示出黑暗与光照条件下的磷虾耗氧率存在较为显著的差异。本研究中,在接近一个自然日的周期内,通过黑暗和100 lx光强两种条件的变化,对试验组磷虾进行反复实验,结果表明这种条件的改变对磷虾的耗氧率影响并不显著,除2号样本外(可能因捕捞过程对个体造成的影响),磷虾的耗氧率水平始终未发生较明显的波动。这在一定程度上说明,冬季时期,磷虾的呼吸速率已降到一个较低的水平,日变化尺度上的光照条件变化并不会对它的新陈代谢速率产生显著影响,进一步表明磷虾低代谢模式可能具有一定的稳定性。考虑到本实验的样本较少,今后可进一步增加样本证实这种机制假设的可靠性。此外,本次实验还发现,黑暗组磷虾的耗氧率变化(标准差)低于光照组,表明光照条件下磷虾的耗氧率波动更大,TESCHKE等[11]的实验中也发现了类似现象,且光照时间越长,光照强度越大,这种波动也越强。这侧面说明了冬季时磷虾低代谢模式较稳定,而本实验中提供的光照条件在一定程度上影响了这种稳定,从而使得磷虾的耗氧率出现相应的波动。
总体上来看,影响磷虾耗氧率的因素较多,有生物因素和非生物因素,生物因素有个体大小、饥饿与摄食、活动与应激及生物周期,非生物因素有温度、盐度、氧张力(PO2)及pH等[20]。由于海上条件所限,较难对各项实验条件均做到精确控制。在实验处理上仍需进一步完善,如实验中所用海水为当地自然循环海水,温度不易控制。为此,为防止对实验结果产生影响,今后的实验中应加以改善,尽可能地稳定实验条件,减少误差。为了深入研究光照条件对磷虾代谢模式的影响,今后可从光强、光照时长等因素进行设置,并在不同季节展开相应的现场实验,以期全面地揭示磷虾耗氧率季节性变化的机制。
致谢:感谢中国水产总公司“龙腾”轮船长、政委、各位船员实验过程中给予的帮助。本项目还得到了农业农村部南极海洋生物资源开发利用项目的支持。
