岩溶区固氮植物适应土壤氮素变化的潜在策略
2021-03-24潘复静章润阳秦国鑫陈英倩梁月明刘坤平
潘复静, 章润阳, 秦国鑫, 陈英倩, 梁月明, 刘坤平, 张 伟
(1.桂林理工大学 a.环境科学与工程学院; b.广西环境污染控制理论与技术重点实验室科教结合科技创新基地,广西 桂林 541006; 2.中国地质科学院岩溶地质研究所, 自然资源部、 广西壮族自治区岩溶动力学重点实验室,广西 桂林 541004; 3.中国科学院环江喀斯特生态系统观测研究站, 广西 环江 547100)
0 引 言
我国西南岩溶地区面积达540 000 km2[1]。该地区的土壤主要由碳酸盐岩风化而成, 成土速率慢, 形成1 cm厚度土壤所需的时间高达几千年至几万年[2-3], 而且土-石二元结构的光滑接触面使得水土容易流失[4], 导致该地区土壤总量偏少。此外, 由于土壤是碳酸盐岩风化而成, 其钙含量较高, 而钙易与有机质和磷结合形成稳定化合物, 氮磷等养分释放的难度大, 氮磷有效性变低[5]。土壤N和P元素是影响植物生长的重要营养元素, 其主要来源是土壤有机质和母岩[6-7], 植物获取这两种养分也主要是从土壤中来。尽管岩溶地区土壤氮磷含量较同纬度地带性土壤高, 然而土壤浅薄、 高钙镁和pH等制约其养分供给能力, 导致植被恢复早期的草丛阶段受到氮限制、 灌木阶段受到氮磷共同限制和后期的乔木阶段受到磷限制的状况[8-9]。
当植物受到N限制或当土壤中的可利用氮含量较低时, 一些植物可以利用空气中的氮进行生物固氮, 这一类植物即固氮植物。固氮植物能够与固氮微生物形成根瘤菌, 将空气中的氮气还原为氨[10]。植物固氮过程受土壤环境中氮素浓度的影响, 不同的固氮植物采用不同的氮素获取策略[11-13]。因此, 基于土壤环境可利用氮含量的响应灵敏性程度, 固氮植物适应土壤氮素变化的固氮策略分为兼性固氮策略、 专性固氮策略、 调节过度固氮策略(介于兼性和专性的固氮策略)共3种。兼性固氮策略是固氮植物的固氮速率随着土壤环境中可利用氮含量的提高而降低, 固氮量以满足自身需求为主; 专性固氮策略是固氮植物的固氮速率不随土壤环境中的可利用氮含量的变化而变化, 即使在氮含量高的土壤中仍然具有较高的固氮速率; 调节过度固氮策略是固氮植物的固氮量少于自身需求量, 其固氮速率不会很高[12]。在土壤中添加无机氮的情况下, 大豆在不同生育期的根瘤数量下降13%~20%, 固氮酶活性下降可达19%~32%[14]。这种情况可能是改变了固氮菌的群落结构, 降低了其多样性, 导致固氮速率下降[15]; 另外, 生物固氮是一种耗能很高的生物化学过程, 植物和固氮微生物在较高氮素浓度情况下直接吸收土壤中的有效氮, 以降低能耗[16]。在自然生态系统中, 兼性固氮策略可能比较常见。另外, 土壤环境的氮素浓度也会影响固氮植物对土壤磷素养分的获取策略。根据前期研究中布设的21个灌丛样方(10 m×10 m), 发现灌丛广泛分布有固氮植物[17]; 岩溶区灌丛生态系统中固氮植物根际土的有效磷含量显著高于非固氮植物[18]。而丛枝菌根真菌与磷酸酶是植物获取土壤磷素的两种重要策略[19-20]。可见, 固氮植物的引种不仅可以消减土壤氮素的限制, 还可以活化土壤磷素[21], 因而被广泛应用于岩溶等生态系统的植被恢复。
在岩溶生态系统, 植物从植被恢复早期的N限制逐步过渡到恢复后期的P限制[9], 也伴随着土壤N含量的逐渐升高[22]。土壤氮含量水平的升高对固氮植物的固氮速率的影响是判断固氮植物的固氮策略的有效指标, 可通过人为添加N素到土壤中, 研究固氮植物的固氮速率。另外, 根据文献[18], 灌丛阶段中固氮植物根际土的有效磷也存在显著差异, 这可能与岩溶区不同固氮植物采用的氮磷养分获取策略有关。而当土壤N含量逐渐升高, 不同固氮植物的氮磷养分获取策略可能发生变化。因此, 本文根据已有研究数据, 确定香合欢(Albiziaodoratissima)与木豆(Cajanuscajan)两种固氮植物作为研究对象, 通过添加氮肥的盆栽试验, 探讨不同固氮植物的固氮策略以及土壤氮素浓度变化对不同固氮植物的氮磷养分获取策略的影响, 为岩溶区植被恢复选择合适的固氮植物提供科学依据。
1 材料与方法
1.1 盆栽试验设置
盆栽试验设在中国科学院环江喀斯特生态系统观测研究站(24°44′N, 107°51′E)。该地区属于亚热带季风气候, 年平均降雨量为1 389 mm, 年平均温度为18.5 ℃。雨季主要集中在每年的4—8月, 占年降雨量的70%[23]。
2017年10—12月, 在研究站区域内采集香合欢与木豆种子。 两种固氮植物是根据早期在岩溶区布设的21个灌丛样地的植被调查数据确定的, 二者是耐贫瘠、 耐干旱、 能够形成根瘤并分布广的固氮植物, 而区别则是香合欢为乔木树种、 木豆为直立灌木树种[17]。2018年2月, 从观测站附近选择岩溶典型灌丛阶段样地, 土壤类型为棕色石灰土。采集0~20 cm厚度的土样(约400 kg), 过2 mm筛后4 ℃保存。从过筛后的土壤随机选取了5份200 g的样品进行养分含量测定, 其全氮和有效磷含量分别为4.48和1.92 mg/kg。
2018年3月, 将过筛土壤用布袋装好, 放入灭菌锅用蒸汽灭菌法在135 ℃下灭菌10 min, 取出后自然降温, 立即把灭菌土填满育苗袋(容积约 0.3 L), 育苗袋预先用75%酒精进行了消毒。盆栽试验设计了2个氮肥添加处理: 添加氮肥(10 gN·(m2·a)-1, 化肥种类为氯化铵(NH4Cl))与不添加氮肥(0 gN·(m2·a)-1)。每个氮肥处理设置16盆, 共32盆, 分别把两种固氮植物的种子播到两种处理的育苗袋各8个重复(2个处理×2种植物×8重复)。种子出苗(大约2周)后, 每盆选择2株长势相似的幼苗移栽到与上述育苗袋的处理相同的塑料盆中(容积约 13 L, 每盆装土 10 kg, 共32盆), 待幼苗生长稳定(约1周)后, 每盆仅保留1株长势较好的幼苗, 每隔2~3天浇一次水以使土壤持水率维持在40%左右。
1.2 样品采集
连续培养15周后(2018年6月), 每个处理中选择长势相似的5盆植物(共20盆, 2个处理×2种植物×5重复), 分别采集固氮植物根瘤、 根、 茎、 叶及根际土样品。根际土采集步骤为: 首先把土壤连同植株从培养盆取出, 捏松土壤, 再轻轻抖动植株和土壤使松散的土掉落, 最后将仍粘附根系表面的土壤轻轻取下则为根际土样品[24]。将收获到的一部分根, 全部的茎、 叶样品在65 ℃下烘干48 h至恒重, 称量, 用于测算生物量和根冠比。烘干后的根、 茎、 叶片粉碎过100目筛(0.15 mm), 用于测定植物各器官的碳、 氮和磷含量。未烘干根样品的一半浸泡于95%乙醇保存, 用于丛枝菌根侵染率的测定; 另一半根样品在-20 ℃保存, 用于酸性磷酸酶活性的测定。一部分根际土壤样品在风干后用于土壤养分含量的测定。
1.3 样品测定
1.3.1 酸性磷酸酶活性测定 植物根系酸性磷酸酶活性采用MUB荧光光度法[25]测定。称取0.5 g植物细根(每个样品3份),剪碎后放入5 mL离心管中。第一份加入2 mL的50 mmol·L-1醋酸钠缓冲液(用冰醋酸调节pH 5.0);第二份加入 800 μL 醋酸钠缓冲液和 200 μL 10 μmol·L-1的标准物(4-甲基伞形酮,4-Methylumbelliferone)溶液;第三份加入 800 μL醋酸钠缓冲液和 200 μL 200 μmol·L-1的底物(4-甲基伞形酮磷酸酯,4-Methylumbelliferyl phosphate)溶液。37 ℃孵育1 h,加入10 μL NaOH(1 mol·L-1)终止反应,用酶标仪(Tecan Infinite M200 PRO,Tecan, 瑞士)于 365 nm波长处激发、405 nm处检测。
1.3.2 菌根侵染率测定 丛枝菌根真菌侵染率采用根段频率常规法[26]测定。将待测根系清洗干净, 并用已消毒的手术剪把根系剪成1 cm长的根段, 再将根段放入50 mL离心管中, 加入足够的10 % KOH溶液浸没根段, 放入90 ℃水浴锅中煮至透明, 用清水洗净根段, 再加2%盐酸酸化, 倒掉盐酸后加入0.01%酸性品红染色, 染色后挑选50个细小根段压片, 显微镜下观察根段侵染状况。
根段侵染率(%)=∑(0×根段数+10%×根段数+20%×根段数+…+100%×根段数)/观察总根段数, 其中: 0, 10%, 20%, …,100%表示根段被侵染的程度。
1.3.3 固氮速率测定 根瘤的固氮速率采用乙炔还原法[20]测定。具体步骤为: 从每个处理中(每株植物)中选取10~15颗颗粒完整、 带有部分须根的粉色、 浅黄色或者褐色的根瘤(为确保根瘤活性, 用于培养的根瘤不提前采摘), 放入125 mL透明的玻璃培养瓶中, 塞紧瓶盖, 用高纯乙炔气体置换瓶中10 %的顶空气体, 在相对一致的环境条件下培养0.5 h后, 抽取瓶中30 mL气体装入已抽真空的12 mL规格储气瓶中, 用GC-FID仪(Agilent GC 7890A, Agilent, USA)测定乙烯含量; 每个样品的根瘤固氮速率做2个技术上的重复、 1个空白对照。测定乙烯后, 通过转化可得到单位质量根瘤的固氮速率(μmol·L-1)。
1.3.4 土壤与植物养分的测定 植物碳含量采用重铬酸钾容量法-外加热法测定。土壤与植物氮含量采用凯氏定氮法-流动注射仪(FIAstar 5000, FOSS, Hillerød, Denmark)测定。土壤氨态氮和硝态氮利用2 mol·L-1KCl浸提, 用流动注射仪(AA3, Seal, 德国)测定。土壤全磷(TP)采用氢氧化钠(片状)熔融, 土壤有效磷(AP)用0.5 mol·L-1NaHCO3浸提, 植物磷含量则采用H2SO4+H2O2消煮, 三者都用钼蓝比色法测定[27]。
1.4 统计与分析
在Microsoft Excel里整理数据, 用SPSS 20.0对数据进行统计分析。利用独立样本t检验及多重比较分析(LSD)方法, 比较施肥与不施肥对两种固氮植物固氮速率、 根系磷酸酶活性、 丛枝菌根侵染率、 植物各器官养分、 土壤养分的差异。采用Pearson相关关系方法分析固氮植物固氮速率、 磷酸酶活性、 菌根侵染率与土壤养分含量之间的相关性。
2 结 果
2.1 氮肥添加对岩溶区固氮植物根际土壤氮磷有效性的影响
氮肥添加下, 香合欢与木豆根际土的全磷、 有效磷、 铵态氮含量显著提高(图1a、 b、 c); 木豆的硝态氮含量提高, 但香合欢的降低(图1d), 香合欢的根际土有效磷含量显著低于木豆(图1b); 不添加氮肥条件下, 香合欢的根际土硝态氮含量显著高于木豆(图1d)。 总体上, 添加氮肥情况下根际土的全磷、 有效磷、 铵态氮含量显著高于不添加氮肥的情况, 而硝态氮的差异不明显。
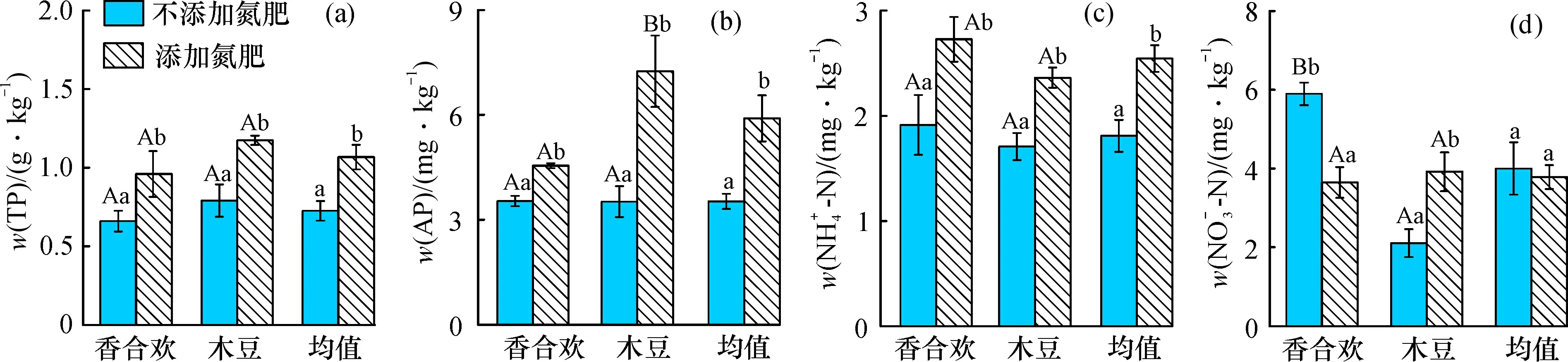
图1 氮肥添加对两种固氮植物根际土的全磷、 有效磷、 铵态氮和硝态氮含量的影响
2.2 氮肥添加对岩溶区固氮植物生物量、 根冠比和养分含量的影响
添加氮肥情况下, 香合欢的总生物量显著提高, 木豆的提高不显著, 总体上生物量有一定的提高(图2a); 香合欢的根冠比显著降低, 木豆的降低不明显, 但总体上显著降低(图2b)。添加和不添加氮肥两种情况下, 木豆的总生物量显著高于香合欢, 但根冠比显著低于香合欢(图2)。
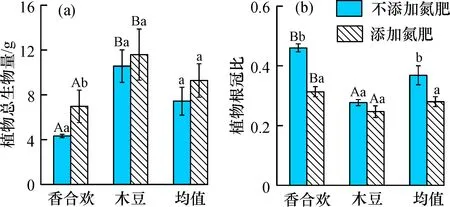
图2 氮肥添加对两种固氮植物总生物量与根冠比的影响
氮肥添加下, 木豆的根系氮和磷含量显著提高(图3b、 c); 木豆和香合欢的茎的氮含量显著降低(图3e)。总体上, 植物茎和叶的碳含量显著高于不添加的情况, 而茎和叶的氮含量显著低于不添加的情况(图3d、 e、 g、 h)。另外, 氮肥添加下香合欢的根、 茎和叶的碳、 氮和磷的含量与木豆的不存在显著差异; 而不添加氮肥情况下, 香合欢根、 茎和叶的氮含量和根系的磷含量显著高于木豆(图3b、 c、 e、h)。

图3 氮肥添加对两种固氮植物根、 茎和叶的碳、 氮和磷含量的影响
2.3 氮肥添加对岩溶区固氮植物固氮速率、 菌根侵染率和根系磷酸酶活性的影响
添加氮肥条件下, 香合欢和木豆的固氮速率显著降低(图4a); 香合欢的菌根侵染率显著降低, 但是木豆的菌根侵染率变化不明显, 总体上显著降低(图4b); 木豆的根系磷酸酶活性显著升高, 但香合欢的根系磷酸酶活性变化不明显, 总体上显著升高(图4c)。
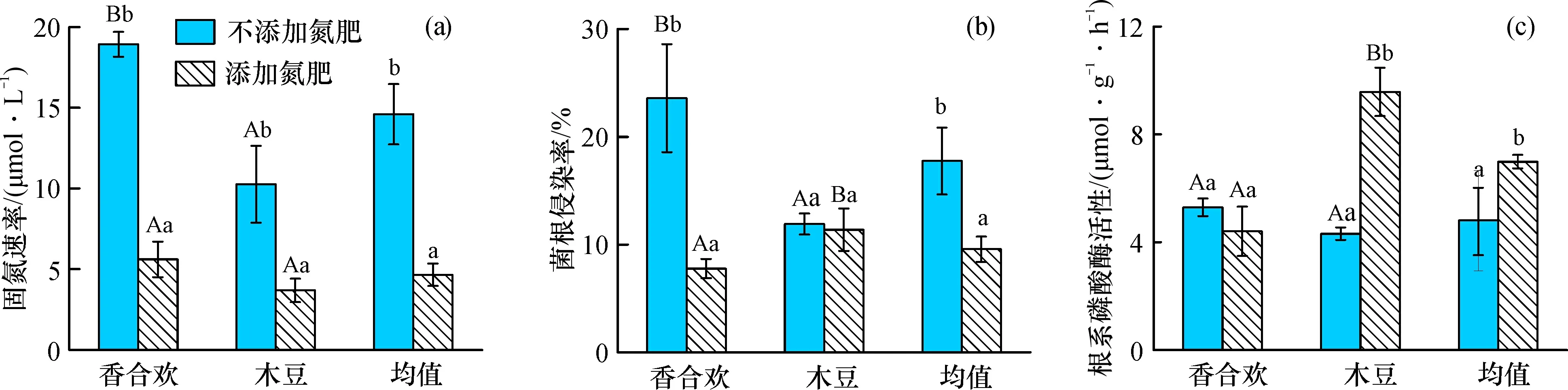
图4 氮肥添加对两种固氮植物固氮速率、 菌根侵染率与根系磷酸酶活性的影响
添加氮肥条件下, 香合欢的菌根侵染率和根系磷酸酶活性低于木豆, 但两种植物的固氮速率无显著差异。不添加氮肥条件下, 香合欢的固氮速率、 菌根侵染率、 根系磷酸酶活性均高于木豆。固氮植物的固氮速率与菌根侵染率和硝态氮存在显著正相关关系, 但与有效磷存在显著负相关关系; 菌根侵染率与硝态氮存在显著正相关关系; 根系磷酸酶活性与有效磷存在显著正相关关系(表1)。
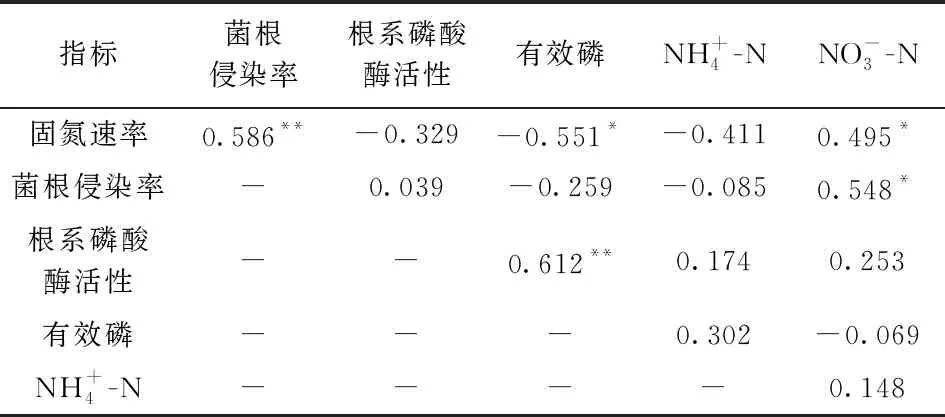
表1 植物的固氮速率、 菌根侵染率、 磷酸酶活性与土壤氮磷有效性的相关性
3 讨 论
3.1 氮添加对岩溶区固氮植物生长的影响
氮肥添加情况下, 总体上岩溶区两种固氮植物总生物量显著提高, 而根冠比显著降低。 具体表现为香合欢的总生物量显著提高, 但是根冠比显著降低, 木豆的总生物量和根冠比变化不明显(图2)。可以看出, 添加氮肥影响了岩溶区固氮植物的生物量和根冠比的变化, 而对香合欢这两方面的影响更大。氮素是植物生长必须的生源要素, 而在岩溶地区, 土壤低氮状况是该地区植被恢复早期植物生长受到限制的一大因素。在较低土壤含氮量的情况下, 植物为了提高吸收养分的效率和数量, 往往通过把生物量分配到地下, 更多的根系生物量使得根系的数量、 根尖的数量、 根系的长度等得到提高[28]。当氮肥添加到土壤当中, 会使得土壤中的供氮水平提高。添加氮肥的土壤有效磷与铵态氮含量显著高于不施氮肥的情况(图1)。在较高土壤含氮量情况下, 充足的供氮量使得植物吸收更加容易或高效, 有利于植物提高光合作用效率而加快生长, 生物量增长迅速(图2a)。氮肥添加情况下植物茎和叶的碳含量显著提高(图3), 且总生物量显著提高(图2a)。植物的养分状况与土壤氮磷含量密切相关[29], 氮肥的添加能够增加植物的磷含量[30]。本研究中, 氮肥添加情况下植物根系和叶片的磷含量得到一定程度的提高(图3)。在营养较为充足的情况下, 植物则会分配更多的生物量在地上部分, 而降低对地下部分的分配。因此, 一般情况下, 氮肥添加到土壤, 植物的根冠比会降低[31]。综上可以得出, 土壤氮素的变化会影响到岩溶区固氮植物的生长以及在养分吸收的生物量分配。
3.2 氮添加对岩溶区固氮植物固氮策略的影响
植物的充足营养、 生物量增长和分配状况会影响到植物的适应性策略。研究发现, 添加氮肥情况下, 岩溶区两种固氮植物的固氮速率显著低于不添加氮肥的情况(图3)。在生态系统中, 固氮植物能够形成根瘤菌, 将空气中的氮气固定供植物生长利用或输入到土壤环境中。固氮植物的固氮速率与土壤环境中可利用氮水平是可以相互反馈的, 当土壤中可利用氮水平较低时, 植物会提高固氮速率, 反之则降低固氮速率[32-33]。例如, 桤木(Alnusincana)幼苗的固氮速率和根瘤生物量[34]和热带固氮植物(Ingapunctate)的共生固氮速率和根瘤密度[20]随着氮添加浓度的增加而降低。原因可能是: 生物固氮是一个极其耗能的过程[32], 植物在缺氮的环境中才会重点利用这个策略, 即在氮素贫瘠环境中固氮植物加强与根瘤菌的共生作用, 提高固氮速率以缓解土壤氮素供应的不足[35]。但是, 随着土壤环境中可利用氮水平的提高, 固氮植物通过根系就可以吸收足够的氮素满足自身的生长, 减弱与根瘤菌的共生作用, 表现为较低的固氮速率[36]。这种固氮策略为兼性固氮策略。综上可以得出, 本研究的两种岩溶区固氮植物的固氮策略为兼性固氮策略。
土壤氮素变化导致的固氮速率的变化也与一系列的根系生物化学特征变化有关。一方面, 土壤氮素变化导致固氮速率与菌根侵染率发生变化。固氮植物的固氮速率与菌根的侵染率具有显著的正相关关系(表1)。丛枝菌根真菌能够促进植物吸收氮磷养分, 已有研究表明, 陆地上80%的植物能够与丛枝菌根形成共生关系[37], 尤其是养分贫瘠的条件下, 植物会增加与丛枝菌根的共生关系, 促进植物对土壤氮磷养分的吸收[38], 从而缓解植物生长受养分的限制。但是, 在养分比较充足的条件下, 植物会减弱与丛枝菌根的共生关系, 降低根系的菌根侵染率, 减少能量消耗。本研究发现, 尽管添加氮肥情况下香合欢根系的丛枝菌根侵染率显著低于不添加氮肥的情况, 而木豆根系丛枝菌根侵染率的变化不大, 但是, 总体上两种岩溶区固氮植物的丛枝菌根侵染率降低明显(图4b)。发生这种现象的原因可能是不同植物种对氮肥添加的浓度范围灵敏性相异, 但是丛枝菌根侵染率与氮肥浓度之间的变化趋势还不够清楚, 未来的研究应布设一系列氮肥添加浓度梯度对固氮植物菌根侵染率的影响研究。
另一方面, 土壤氮素变化导致磷酸酶活性变化。磷酸酶是植物获取土壤磷素的一种重要策略, 能够把土壤中的有机磷转化为无机磷, 有利于植物磷素的吸收[39-41]。本研究中, 氮肥添加显著提高了木豆根系的酸性磷酸酶活性而稍微降低了香合欢根系的酸性磷酸酶活性, 总体上则是氮肥添加提高了根系的酸性磷酸酶活性(图3), 出现此现象可能与植物的生物量和养分含量的提高有关: 土壤中氮素增加导致植物的生物量提高, 养分的吸收量提高, 充足的氮素供应使得植物对磷素的需求持续增大, 可能会导致有效磷的供应缺乏。一般情况下, 土壤磷素有效性较低的环境将有利于刺激植物分泌酸性磷酸酶[42-44]。以上研究表明, 植物根系磷酸酶活性除了受环境中土壤磷素含量影响外, 还与植物种适应环境养分含量变化的适应策略有关。在岩溶地区, 植被演替早期的土壤氮含量较低, 植物受到氮限制, 而随着植被恢复的正向演替, 土壤的氮含量逐渐提高, 磷含量提升不明显, 演替后期的植物更容易受到磷限制[9, 45]。氮添加控制试验模拟了岩溶地区植被恢复土壤氮含量的变化, 氮素的充足使得植物更容易缺磷, 从而促使磷酸酶活性的提高。
3.3 岩溶区生态恢复中固氮植物的潜在选择
不添加氮肥情况下, 香合欢的根冠比、 根系的氮磷含量与茎、 叶的氮含量、 固氮速率、 菌根侵染率、 根系磷酸酶活性高于木豆。这种现象表明, 在岩溶地区植被演替早期土壤氮含量较低的情况下, 香合欢比木豆更容易适应土壤的氮限制。此外, 添加氮素使得木豆的菌根侵染率和根系磷酸酶活性高于香合欢, 表明在岩溶地区随着植被恢复后期土壤氮素的升高, 木豆比香合欢更能适应植被恢复后期的土壤环境。固氮植物可以使得生态系统中土壤氮含量升高, 为其他生物物种提供氮素资源。岩溶生态系统是一个生态脆弱、 养分缺乏、 植被可持续发展差的生态系统, 其生态恢复应需进行人工干预。基于以上结果和讨论, 在岩溶地区生态恢复早期进行人工干预时选择香合欢作为植物群落配置的物种更为合适, 而在生态恢复后期选择木豆作为植物群落配置的物种更为合适。
4 结 论
本文通过控制性试验研究发现, 氮肥添加有利于提高岩溶区固氮植物的总生物量与土壤铵态氮和有效磷含量, 但是降低了植物的固氮速率。植物共生固氮速率受环境氮素浓度的影响, 高氮会抑制固氮植物的固氮速率, 岩溶区固氮植物的固氮策略为兼性固氮策略。
不添加氮肥情况下, 香合欢的根冠比、 根系的氮磷含量与茎、 叶的氮含量、 固氮速率、 菌根侵染率、 根系磷酸酶活性高于木豆; 添加氮素情况下, 木豆的菌根侵染率和根系磷酸酶活性高于香合欢。可见, 在岩溶地区生态恢复早期进行人工干预时选择香合欢作为植物群落配置的物种更为合适, 而在生态恢复后期选择木豆作为植物群落配置的物种更为合适。但是, 岩溶地区不同植被恢复阶段生长有大量的固氮植物, 要揭示不同固氮植物的固氮策略以及为植被恢复提供适宜不同恢复阶段、 不同地区、 不同坡位的物种配置方案, 还需要广泛、 深入地开展野外定位监测和多因子控制试验的综合性研究, 这是今后工作的一个重要方向。
