中日沿海部分鱼类DNA条形码研究
2021-03-18黄志基张玉宁
罗 纯,章 群,黄志基,张玉宁
(暨南大学生态学系,广州 510632)
中国与日本海域相连,是南北暖冷鱼类的重要交汇区。一方面,由于同源扩散的结果,两国沿海部分鱼类特别是近海种类呈现高度相似性;另一方面,由于地理环境有所区别,再经过历史演替,两国沿海鱼类在共性的基础上也呈现一定差异[1]。近年来,随着中国沿海经济不断发展,海洋渔业资源被过度利用,渔获物低龄化现象严重[2],对海洋鱼类资源开展深入研究有助于渔业的长期、可持续性发展。
为了有效保护和管理鱼类资源,对鱼类进行鉴定分类必不可少。由于传统分类基于外部形态特征,对分类者的专业知识和经验要求较高,同时有些物种形态结构易受到地理环境和不同发育阶段的影响;还有部分鱼类存在雌雄转换现象,具有性别二态性[3]:如隆头鱼科鱼类为世界性分布,多数种存在幼鱼期、成鱼期及超雄期之间以及雌雄之间的体色和形态的差异[4],给物种分类鉴定造成了一定困难。
DNA条形码不仅可以识别整条鱼,还可以识别鱼片、鱼鳍和其他难以根据形态学识别的鱼体碎片[5]。2005年WARD等[6]利用COI基因对澳大利亚207种鱼类进行鉴定,表明COI基因能够有效区分不同物种。动物线粒体COI基因5′端约650 bp序列已被公认为是有效区分不同物种的DNA条形码标准区域[7-8]。大量研究也表明,线粒体DNA条形码技术在解决同种异名、异种同名、发掘隐藏种方面较传统形态分类具有显著优势。
目前,对中日沿海鱼类的研究报道多为生态资源调查[1]、遗传结构与分子系统地理学研究[9-11]等。在分类方面,日本学者河野光久等[12]对日本海西南部山口县的鱼类进行了统计学研究,共确认了39目197科870种。荒见真一郎等[13]运用DNA条形码技术鉴定了神奈川县市场31种鱼类。李昂等[14]对中国近海和日本近海的白姑鱼分类地位和系统进化进行了研究。本研究测定了在日本沿海采集的37种鱼类的184条COI基因序列,并结合GenBank下载的序列,共分析了中日沿海共有种327条COI基因序列,以期确定日本及中国沿海鱼类的分类地位,为相关海域鱼类种质资源保护和可持续利用提供科学指导。
1 材料与方法
1.1 样品来源及分子实验
研究样品于2018年4月—2019年7月采集于日本太平洋侧濑户内海明石冲(34°38′41″N、134°59′23″E)和日本海侧若狭湾舞鹤(35°27′01″N、135°18′52″E)等地,并置于95% 乙醇中保存。依据Fishbase数据库,并参照《中国鱼类系统检索》[15]、《黄渤海鱼类图志》[16]等资料进行形态鉴定,测序标本和下载序列信息见表1。
取0.2 g鱼尾鳍肌肉晾干后,参考乐小亮[17]改进的苯酚/氯仿法提取DNA。参考WARD[18]方法进行PCR 扩增。使用引物FishF1:5′-TCAACCAACCACAAAGACATTGGCAC-3′,FishR1:5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′,反应体系为20μL:上下游引物各1μL,PCR Mix 10 μL,DNA模板1μL,ddH2O 7μL。PCR反应条件为:95℃预变性5 min,95℃变性40 s,55℃退火40 s,72℃延伸50 s,共35个循环,最后72℃延伸10 min。4℃保存。扩增产物经1% 琼脂糖凝胶电泳检测后送至北京六合华大基因公司正向测序。
1.2 数据处理
通过Bioedit 7.0.9.0[19]软件上对测序峰图进行人工校对。运用MEGA 7.0[20]软件分析序列特征,计算碱基组成、变异位点、简约信息位点、单一可变位点等分子多样性指数;基于Kimura 2-parameter(K2P)替代模型,分别计算种内、属内种间、属内、科内属间遗传距离,经1 000次重复抽样(bootstraps)检测分支置信度构建邻接(neighbor-joining)树。采用自动条形码间隙检索方法 (automatic barcode gap discovery,ABGD)[21]将全部序列自动分组进行假设种分析,同时利用种内遗传距离及种间遗传距离绘制条形码间隙图。
2 结果与分析
2.1 COI基因序列特征
参照国际上通用的DNA条形码长度,截取平均长度为638 bp(552 bp ~652 bp)的5′端COI序列进行分析[22]。在本研究测定序列和GenBank下载的7目24科35属40种327条序列中,没有发现碱基插入和缺失现象。共检测到315个变异位点,其中简约信息位点有308个,单一可变位点有7个。碱基T、C、A、G的平均含量分别为29.4%、28.5%、23.2%、18.8%,A+T含量(52.7%)高于G+C含量(47.3%),表现出明显的碱基组成偏向性,该结果与动物COI基因碱基组成中普遍存在的AT含量高于GC含量的特征一致[18]。
2.2 各阶元平均遗传距离概况
各分类阶元的K2P遗传距离如表2和图1所示。40种鱼类种内遗传距离为0~3.91%,平均遗传距离为0.42%;属内种间遗传距离为0.32%~17.64%,平均遗传距离为6.25%;属内种间平均遗传距离是种内平均遗传距离的14.88倍,符合HEBERT[22]提出的物种鉴别“十倍法则”,即种间遗传距离远大于种内遗传距离的10倍以上。图1显示:种内遗传距离98%集中在0~2%区间;属内种间遗传距离集中在2%~18%区间内,二者形成了明显的条形码间隙。
2.3 条形码分子系统树分析
从图2邻接树看,327条序列形成了41分支,平均遗传距离分支间21.81%(1.97% ~28.29%)是分支内0.30%(0~0.81%)的73倍(图3),40种鱼类有36种形成了单系分支,支持其物种有效性。白姑鱼(Pennahia argentata)形成2个分支,中国和日本全部个体各自聚类成一支,分支间遗传距离为3.28%。花鲈(Lateolabraxspp.)形成2个自展支持率为100%的分支,日本和中国个体分别成支,分支间遗传距离为6.98%。赫氏高眼鲽(Cleisthenes herzensteini)和松木高眼鲽(C.pinetorum)混杂分布在同一分支上,分支内的遗传距离为0.23%。在ABGD分析中(图4),当种内先验遗传距离P为0.021 5% 时,40种鱼类被划分为40个假设种。瓦氏(Callionymus valenciennei)和饰鳍斜棘(Repomucenus ornatipinnis)被归为一个假设种,其余种的划分与邻接树形成的分支相一致。

·续表1·
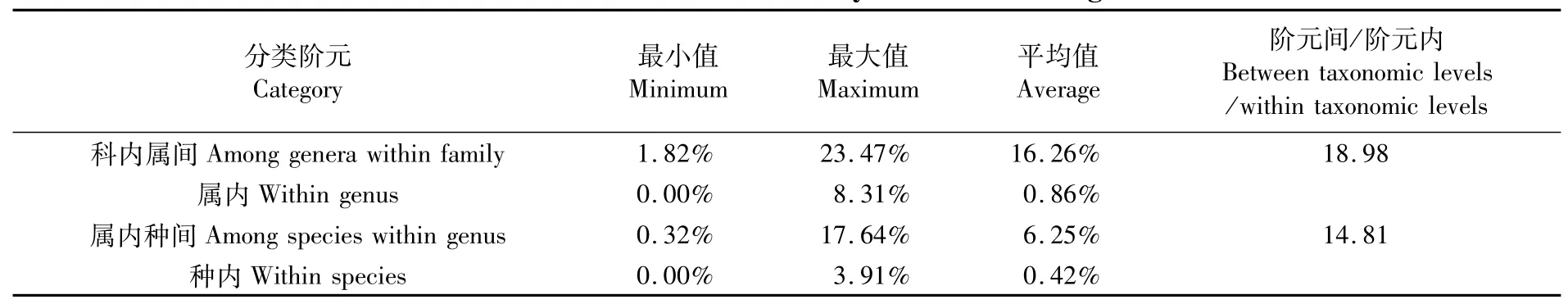
表2 不同分类阶元的遗传距离Tab.2 Genetic distance summary of different categories

图1 不同分类阶元的遗传距离图Fig.1 Distribution chart of genetic distance for 40 species based on the K 2P model
本研究采取同一物种内多个个体比较分析的策略,计算每一种鱼类种内不同样本间的遗传距离,结合各种间遗传距离绘制40种鱼类条形码间隙图,以获得种内与种间遗传距离的差异度。图5显示松木高眼鲽(9)和赫氏高眼鲽(10)最大种内遗传距离未能与最小种间遗传距离很好的分离,其余98%的鱼类均能够形成良好的条形码间隙。
3 讨论
3.1 花鲈与白姑鱼分类地位的探讨
YOKOGAWAL等[23]研究发现,日本花鲈(L.japonicus)和中国花鲈(L.maculatus)在形态和遗传上具有显著差异,前者侧线鳞片数76 ~92,鳃耙数24 ~30,后者侧线鳞片数66 ~82,鳃耙数19 ~24;且基于同工酶等位基因频率计算的遗传距离为0.174,应为2个不同的物种。中山耕至[24]研究发现,中国花鲈和日本花鲈细胞色素类型与分布不同。更新世冰期海平面下降,日本海和中国东海成为2个独立的避难所,使分布其中的鲈鱼逐渐分化[25]。在本研究中,花鲈种内平均遗传距离为3.91%,大于2%的物种遗传阈值;在邻接树上形成中国群体和日本群体2个分支,分别对应于中国花鲈和日本花鲈,分支间平均遗传距离(6.98%)是分支内平均遗传距离(0.49%)的14.2倍,符合“10倍法则”;支持将二者作为2个物种的分类处理。
在邻接树上,中国和日本白姑鱼各自独立成支,平均遗传距离分支间(3.28%)是分支内(0.29%)的11.3倍;ABGD也将白姑鱼分为2组。目前,国内外学者已从生物学特性等方面对其进行了研究,韩志强等[26-27]通过白姑鱼线粒体DNA控制区和Cytb基因序列和AFLP技术对白姑鱼系统地理分布模式的研究,表明中日白姑鱼组群遗传分化显著;李昂[28]通过对中日白姑鱼的耳石形态和线粒体基因组全序列分析发现,二者存在一定程度的分化,认为边缘海的隔离是造成白姑鱼分化的原因,海洋洋流、海水温度以及东海深海沟是维持当前分化的主要因素[27]。本研究中,根据物种鉴定的种间遗传距离是种内遗传距离的10倍和种内遗传距离一般小于2%法则,推测中国沿海和日本的白姑鱼是两个亚种乃至不同种。

图2 基于线粒体COI基因序列的40种鱼类邻接树Fig.2 Neighbor-joining tree based on m tDNA COI sequences of 40 fish species

图3 41个分支的分支内(左)和分支间(右)遗传距离Fig.3 Genetic distance w ithin(left)and among(right)41 clades

图4 中日沿海部分鱼类的ABGD分析图Fig.4 Automatic barcode gap discovery analysis of some coastal fishes in China and Japan
3.2 高眼鲽属分类地位的探讨
赫氏高眼鲽和松木高眼鲽混杂在遗传距离为0.23%的分支内,种间平均遗传距离(0.32%)和种内遗传距离(0.23%),均在一般物种的种内遗传范围内。由于没有赫氏高眼鲽的凭证标本,推测可能存在鉴定错误的情况,但不排除存在谱系拣选尚未完成的可能性。
3.3 瓦氏与饰鳍斜棘分类地位的探讨

图5 40种鱼类DNA条形码间隙图Fig.5 DNA barcoding gap for 40 fish species
3.4 花鳍海猪鱼的分类探讨
本研究中,花鳍海猪鱼形态差异较大,其种内平均遗传距离为0.56%,且能形成单系分支。花鳍海猪鱼幼体表现为雌性,成体后转换为雄性,且雌雄个体之间存在体色和形态的差异[33-34],易被误认为不同的物种。福井行雄等[35]研究发现,雌性个体体色偏黄,体侧中央有一细的明显褐色纵带,自吻端经眼径直达尾鳍基,此纵带在胸鳍上方及尾鳍基处黑色加深;而雄性个体在胸鳍基底后方上具一大黑斑,且背鳍、臀鳍和尾鳍有红色斑纹。而本研究中花鳍海猪鱼的形态差异就是雌雄个体的差异。事实上,大部分隆头科鱼类存在性别二态性,了解雌雄之间的差异对于种类鉴定、形态研究以及判断雌、雄在繁殖过程中的不同角色有着重要的意义。
综上所述,基于COI基因的DNA条形码技术能够对日本大部分海洋鱼类进行有效的区分,说明线粒体COI基因作为日本海洋鱼类DNA条形码适用性广,对于目前研究尚不深入的非经济鱼类,尤其是易引起误鉴的雌雄异形的鱼类,展开线粒体COI基因的研究工作尤为重要。本研究中,由于西北太平洋独特的地理构造和复杂的洋流体系,分布于中日沿海的鱼类呈现多样的分布格局。对于中日海洋鱼类来说,绝大多数种类并没有显著的遗传分化,这可能与黑潮和亲潮的输送作用有关。而中日白姑鱼达到亚种乃至2个不同种的水平、日本花鲈和中国花鲈间的遗传分化,应是与具体类群的独特生理生态习性和边缘海的隔离有关。由于可参考的鱼类检索工具书的欠缺,样品的地理分布广泛,难以获得不同区域样品进行近缘种的比较,导致存在物种分类鉴定错误的可能,如松木高眼鲽和赫氏高眼鲽可能因错误鉴定而出现聚在同一分支上的情况。另外,母系遗传的线粒体COI基因不能反映双亲遗传的核基因信息,难以区分物种的杂交渗透状况。因此,准确的物种鉴定不仅需要DNA条形码进行辅助研究,在今后的工作中仍需补充其同属其他物种,同时结合形态测量、核基因标记以及生物学研究,以进一步明确日本与中国海洋鱼类的分类地位。
