毛果杨GATA基因家族全基因组水平鉴定及表达分析
2021-02-26任梦轩王瑞琪魏志刚
任梦轩 张 洋 王 爽 王瑞琪 刘 聪 魏志刚
(1. 林木遗传育种国家重点实验室,东北林业大学,哈尔滨 150040;2. 国家林业与草原局盐碱地研究中心,中国林业科学研究院,北京 100091)
转录因子在植物生长发育以及应对环境变化中发挥极为重要的作用[1]。研究表明,植物基因组有6%~10%基因编码产物为转录因子,主要包括AP2/ERF、ARF、bZIP/HD-ZIP、C2H2、GRAS、MYB/MYC、WRKY、bHLH、NAC 和GATA 等家族[2]。如拟南芥(Arabidopsis thaliana)具有34 个转录因子基因家族,共有1 533 基因,其中约45%的转录因子属于植物特有的[3]。
1988 年,Evans 等[4]首次报道(T/A)GATA(A/G)序列是在鸡的珠蛋白基因启动子上,1991 年发现了转录因子GATA-1。随后,又鉴定出GATA-2、GATA-3、GATA-4、GATA-5 和GATA-6 等基因,由此形成了GATA 转录因子家族[5]。随后,在植物中也鉴定出一批能结合WGATAR 序列(W 为T 或A;R为G或A)的转录因子,且大多含有1个CX2CX18CX2C结构域,有些成员则含有一个CX2CX20CX2C结构域或编码一个2 锌指结构域[6~7]。目前为止,已先后对拟 南芥[7]、蓖麻(Ricinus communis L.)[8]、苹 果(Malus domestica)[9]、水稻(Oryza sativa L.)[10]、番茄(Solanum lycopersicum)[11]和辣椒(Capsicum annu⁃um L.)[12]等植物的GATA 家族成员进行了全基因水平的鉴定。如拟南芥GATA 基因家族包含30 个基因,可分成4 个亚族,并且在光响应调控和叶绿素合成[13~16]、细胞分裂素响应[17~18]、花发育[19]和种子萌发[20]等多个生物学过程扮演着重要角色。
毛果杨(Populus trichocarp)是第一个全基因测序并注释的木本植物,并且具有相对较小的基因组(422.9 Mb)、广泛的适应性、容易的转基因操作和较高生长速率等特点,因此其已成为研究木本植物复杂性状形成分子机制、基因功能解析、比较基因组学等分子生物学与分子遗传学研究中的模式物种[21]。研究表明,毛果杨基因组含有58 个转录因子基因家族,共有4 287 个转录因子基因[22]。然而,目前为止,尚无有关毛果杨GATA 转录因子家族基因基本特性的分析报导。为此,本项研究从基因组水平对毛果杨GATA 基因家族的基因与编码蛋白的基本特性,以及各基因的组织与盐胁迫响应表达特性进行初步研究。上述工作将对毛果杨GATA 家族各基因在其生长发育与盐胁迫响应中的生物学功能解析奠定了基础。
1 材料与方法
1.1 植物材料
毛果杨材料来源于林木遗传育种国家重点实验室(东北林业大学)温室(长日照16/8 h、25 土2℃)培养的3 个月大小的植株。选择生长良好且长势一致的植株平均分为两组,一组为处理组(200 mmol·L-1NaCl 处理48 h),另一组为对照组(野生型)。上午10:00,对两组材料的初生茎节(1~2 茎节)、过渡茎节(3~4 茎节)和次生茎节(5~8茎节)进行取样,每个样品3次生物学重复。
随后,用锡箔纸封好迅速放入液氮速冻,-80℃保存,用于后续RNA 提取,以作为GATA 基因家族组织表达特异性实验材料。
1.2 毛果杨GATA转录因子家族成员鉴定
从植物转录因子数据库(http://planttfdb.cbi.pku.edu.cn/)下载的拟南芥GATA 家族各基因编码的氨基酸序列。利用HMMER 软件(http://www.ebi.ac.uk/Tools/hmmer/)和SMART软件对基因组数据库进行搜索,从而鉴定出39个候选基因。
1.3 毛果杨GATA 转录因子家族染色体定位分析
利用Phytozome 数据库(https://phytozome.jgi.doe.gov/pz/portal.html)获得染色体位置,通过软件MapInspect(http://www.plant- breeding.wur.nl/UK)[23]来制作染色体定位图。
1.4 毛果杨GATA转录因子家族的进化分析
运用BioEdit 软件对拟南芥、毛果杨的GATA氨基酸序列进行多序列比对,然后通过进化树软件MEGA6.0,运用相邻连接法(neighbor joining),执行参数为Poission correction、pairwise deletion 和bootstrap 1 000 次重复[23],构建毛果杨和拟南芥的系统进化树。
1.5 毛果杨GATA 转录因子家族基因结构的分析
基因内含子和外显子模式图用GSDS 网站绘制,内含子外显子信息下载于植物转录因子数据库(http://planttfdb.cbi.pku.edu.cn/)和SGN(https://solgenomics.net/)[23]。
1.6 毛果杨GATA转录因子家族保守原件分析
通过在线软件MEME(http://meme-suite.org/)预测蛋白保守基序[23]。
1.7 毛果杨GATA转录因子家族保守结构域分析
利用序列分析软件BioEdit 分析蛋白保守域序列[23]。
1.8 毛果杨GATA 基因qRT-PCR分析
根据毛果杨GATA 家族各基因序列设计特异定量引物(见表1),利用CTAB 法提取总RNA,反转录得到cDNA 为模板,进行实时定量RT-PCR 分析。体系如下:2×TransStart®TOP/Tip Green qPCR Supermix 10 μL、PtrIDP1-QF/QR 混合引物(10.0 μmol·L-1)0.4 μL、cDNA 1.5 μL,Passive Reference Dye(50x)0.4 μL,加ddH2O 至20 μL。PCR 反应程序如下:94℃30 s;94℃5 s,60℃15 s,72℃35 s,40次循环;95℃15 s,60℃1 min,95℃30 s。每个样品进行3 次生物学重复,利用2-△△CT法进行数据处理。
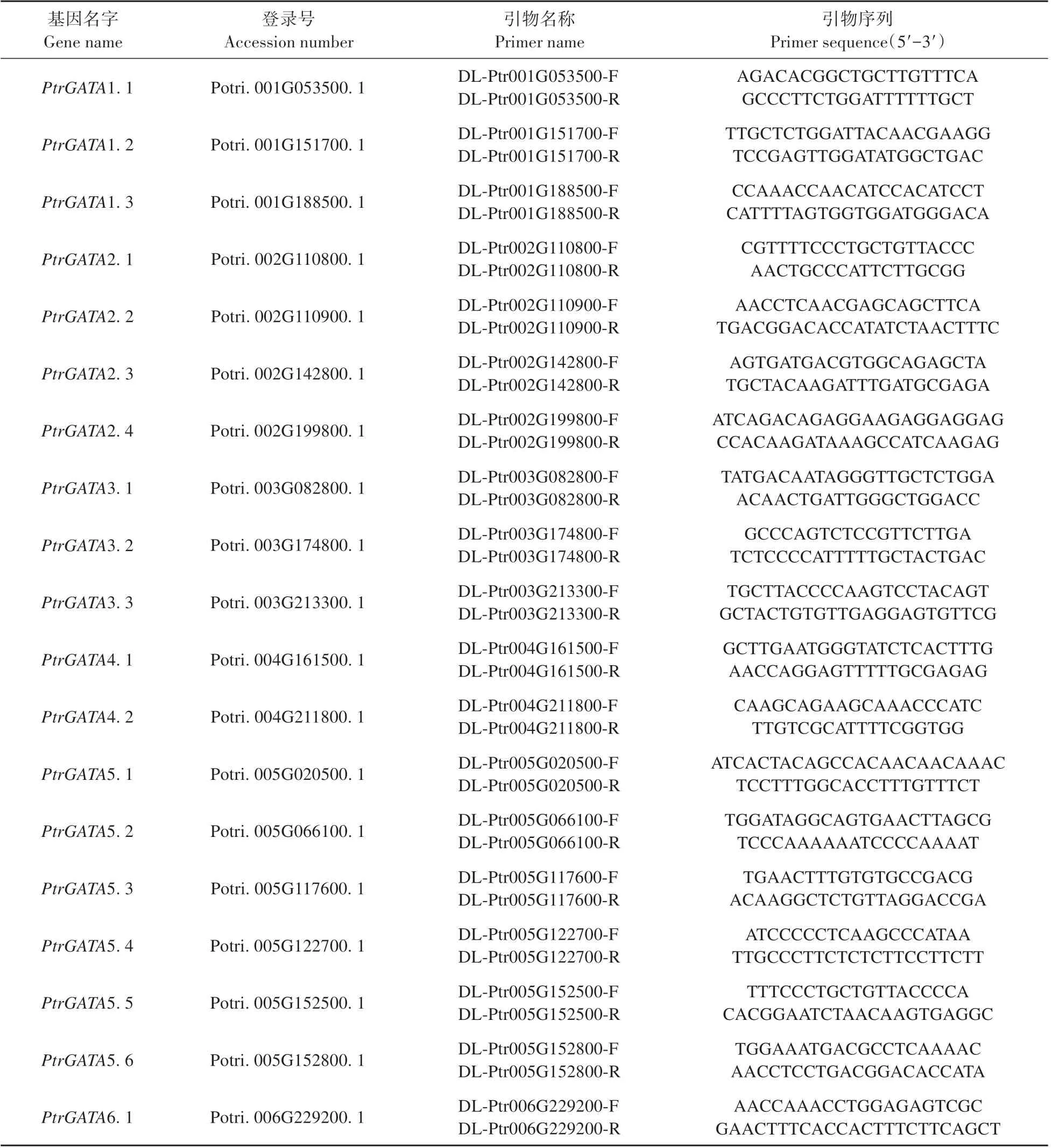
表1 毛果杨GATA家族基因qRT-PCR的引物Table 1 Primers for quantitative real-time PCR of P.trichocarpa GATA family
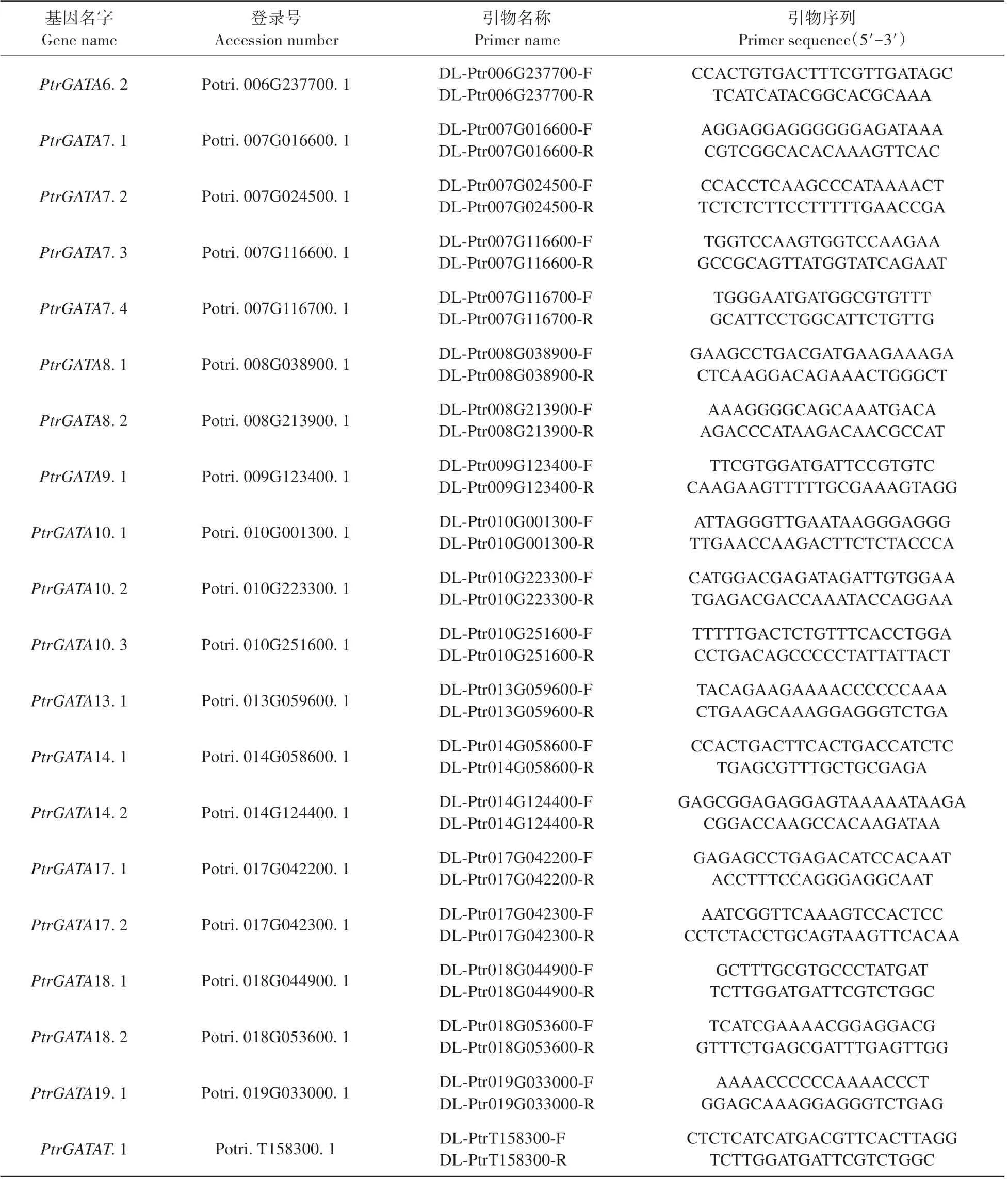
续表1 Continued table 1
2 结果分析
2.1 PtrGATA 家族成员基本信息及其在染色体上的分布
利用拟南芥中已鉴定的30 个GATA 基因编码氨基酸序列在毛果杨基因组数据库中同源比对筛选到39 个PtrGATA 基因,根据拟南芥直系同源基因对应分别命名为PtrGATA1.1-PtrGATA19.1。PtrGATA 家族各成员基因及其编码蛋白的基本信息分析表明,各成员CDs 长度变化范围内402~1 659 bp,编码蛋白的氨基酸序列长度为133~552 aa、分子量为12 867.23~61 305.85 Da、等电点4.87~9.77(见表2)。上述结果表明,PtrGATA 家族基因及其编码蛋白基本特性存在较大变化,预示家族各成员的生物学功能可能也产生了分化。
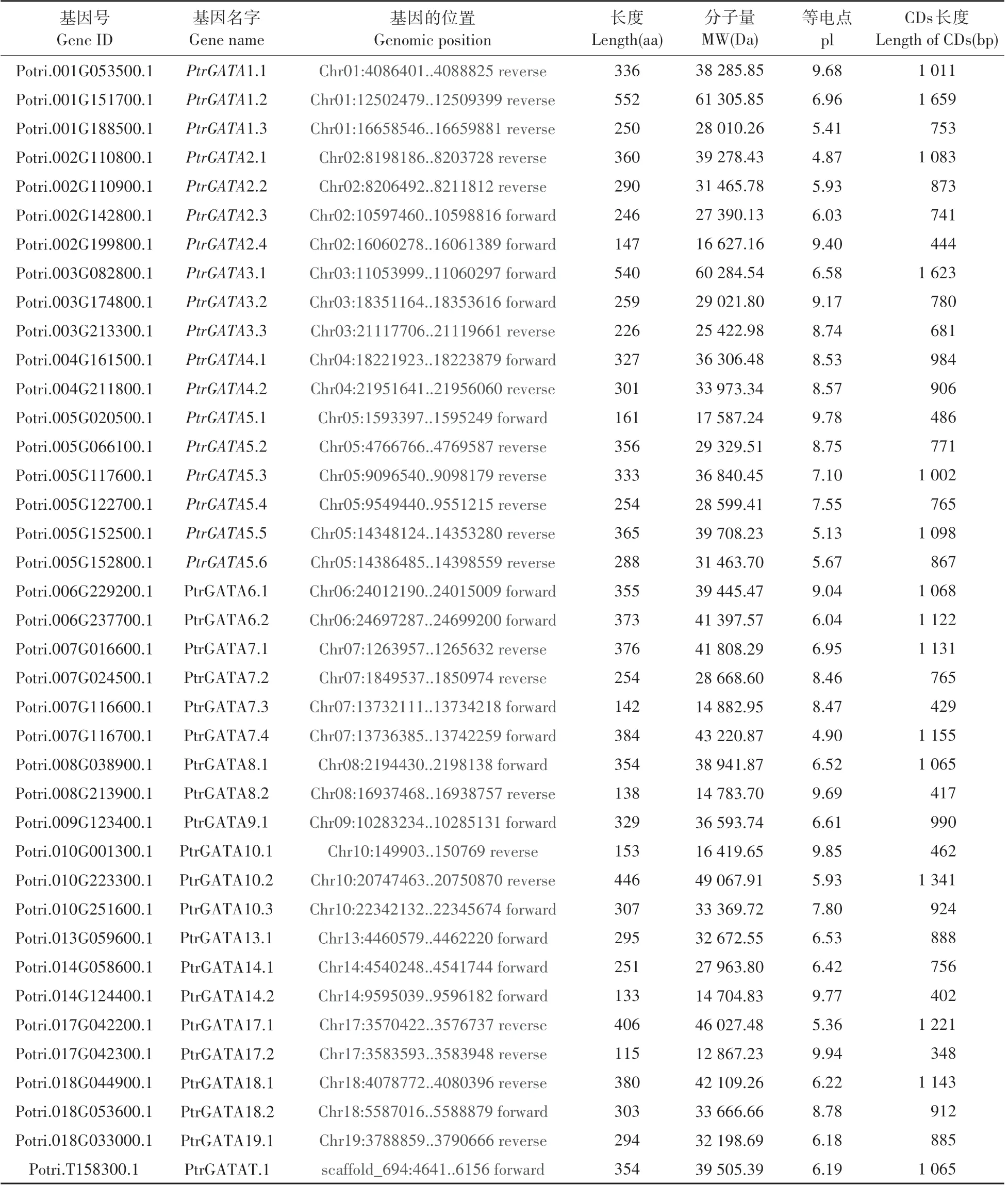
表2 毛果杨GATA家族各基因概况Table 2 Summary of GATA genes in P.trichocarpa
基于PtrGATA家族成员基因组的基本信息,我们对PtrGATA 家族成员在毛果杨染色体上的分布情况进行了分析,结果(见表2,图1)表明,PtrGATA家族在19 条染色体呈不均匀分布,其中5 号染色体上分布了6 个PtrGATA 基因;2 号和7 号染色体均分布4 个PtrGATA 基因;1 号、3 号和10 号染色体上分布了3 个PtrGATA 基因;4 号、6 号、8 号、14 号、17 号和18 号染色体各分布2 个PtrGATA 基因;9 号和19 号染色体各自分布一个基因;11 号、12 号、15号和16 号染色体上无PtrGATA 分布。上述结果表明,毛果杨中PtrGATA基因的扩增模式为分散复制与片段复制。
2.2 毛果杨PtrGATA家族的系统进化
构建系统发育进化树将基因分类对于基因家族各成员功能预测与相互关系分析极为重要。由于拟南芥GATA 家族各基因研究较多,因此本项研究将结合拟南芥GATA 家族进化信息对毛果杨初步分析。为此,利用MEGA 6.0 软件制作了69 个GATA 转录因子的蛋白序列(毛果杨39 个,拟南芥中30 个)进化树,如图2 所示,毛果杨39 个GATA基因分成4 个亚族,其中亚族Ⅰ包含为:PtrGA⁃TA1.1、 PtrGATA1.3、 PtrGATA2.3、 PtrGATA3.2、PtrGATA4.1、PtrGATA4.2、PtrGATA5.2、PtrGATA5.3、PtrGATA6.2、PtrGATA7.1、PtrGATA8.1、PtrGATA9.1、PtrGATA10.2、PtrGATA13.1、PtrGATA14.1、PtrGA⁃TA18.1、PtrGATA19.1 和PtrGATAT.1;亚 族Ⅱ为:PtrGATA2.4、PtrGATA3.3、PtrGATA5.1、PtrGATA5.4、PtrGATA6.1、PtrGATA7.2、PtrGATA8.2、PtrGATA10.1、PtrGATA14.2 和PtrGATA18.2;亚 族Ⅲ为:PtrGA⁃TA2.1、PtrGATA2.2、PtrGATA5.5、PtrGATA5.6、PtrGA⁃TA7.3、PtrGATA7.4、PtrGATA10.3、PtrGATA17.1 和PtrGATA17.2;亚族Ⅳ:PtrGATA1.2 和PtrGATA3.1。毛果杨GATA 家族成员被分成了4 个亚家族和拟南芥一致[8],且各亚族同源性较高,暗示着毛果杨和拟南芥的直系同源基因具有相似功能。
2.3 毛果杨GATA 转录因子家族基因结构和保守基序的分析
为进一步分析毛果杨GATA 家族各基因结构及其编码蛋白的特征,我们利用在线软件MEME预测了毛果杨39 个GATA 转录因子的保守基序(见图3A),参数设置为:保守基序最佳匹配长度为6~100,保守基序个数为10,其他参数设置为默认。利用在线网站GSDS绘制毛果杨GATA 转录因子家族基因结构示意图(见图3B)。毛果杨GATA家族不同亚族基序分布如下:亚族中毛果杨GATA转录因子具有相似的保守基序(见图3A),比如在Ⅰ亚族中,除了含有特有的Motif 1 基序,还含有Motif 2、Motif 4、Motif 6 和Motif 9 基序。在Ⅳ亚族中,除了含有Motif 1 基序之外,还含有Motif 3、Mo⁃tif 5 基序。这说明不同亚族中蛋白基序的不同可能是其功能的分化的原因或动力。同时,在相同亚族中GATA 转录因子相似的保守基序表明其含有相似的功能;内含子-外显子结构图是基于毛果杨GATA 的CDS 序列和DNA 序列之间的关系,毛果杨GATA 基因从1 个外显子(PtrGATA17.2)到11个外显子(PtrGATA7.4)的结构变化,毛果杨第Ⅰ亚族中每个基因的内含子平均个数是最少的(2.3个),在Ⅳ亚族中最多(8 个)。在同一亚族家族成员中,内含子个数差异不大,且在同一亚族中具有相似的结构,在结构上的变化暗示着其基因组可能在进化过程中出现了变化(见图3B)。
2.4 毛果杨GATA 转录因子家族保守结构域分析
为了进一步分析39 个毛果杨GATA 转录因子蛋白序列的结构,我们使用DNAMAN 进行了氨基酸序列比对。序列比对结果表明(见图4):大多数毛果杨GATA 转录因子都包含C-X2-C-X17-20-CX2-C锌指结构域,其中第Ⅰ、第Ⅱ和第Ⅲ亚族的锌指结构域为C-X2-C-X18-C-X2-C,第Ⅲ亚族的锌指结构域为C-X2-C-X20-C-X2-C,没有发现C-X2-CX17-C-X2-C 锌指结构域。其中第Ⅲ亚族的PtrGA⁃TA7.3 无C-X2,没有完整的保守结构域;第Ⅵ亚族的PtrGATA1.2 和PtrGATA3.1 无C-X2-C-X20-C-X2-C,没有保守结构域,可能是在进化上发生了变化。该序列特征与在拟南芥中的研究一致。另外,毛果杨GATA 锌指结构域的大部分氨基酸位点是高度保守的。亚族与亚族之间的氨基酸位点也是较为保守,例如,Cys-3、Cys-6、Thr-13、Pro-14、Gly-19和Pro-20,以及第二个半胱氨酸对的侧翼序列(LCNACG)。分析发现,该家族在整体上具有较强的保守性。可以看出,毛果杨PtrGATA1.2 和PtrGATA3.1 这2 个成员与其他37 个成员在保守性上有一定差异,亲缘关系较其他成员较远,表明这些成员在进化演变中在保守域上发生了一定的改变。
2.5 毛果杨GATA 基因家族组织表达特异性分析
为了研究GATA 家族各基因在毛果杨不同组织的功能,我们利用MeV 软件对phytozome 网站中39 个GATA 基因在各个组织中的表达量绘制色温图并进行分析,结果(见图5)表明,39 个GATA 基因在各组织表达量存在差异,如除PtrGATA5.2 在各个组织中表达量都较低外,其余38 个基因在检测的组织中均有较明显的表达。大部分基因的表达具有组织特异性,如第Ⅰ亚族、第Ⅲ亚族和第Ⅵ亚族中个各家族成员均在茎部相对表达量较高,暗示了同一亚族亲缘关系可能比较相近,在植物生长过程中发挥的功能相似;第Ⅱ亚族中各家族成员基因组织偏爱性表达明显在叶部,相比较其他组织部位,叶部表达量较高。特别是PtrGA⁃TA8.2、PtrGATA14.2和PtrGATA18.2三个基因,大量文献报道GATA 家族转录因子参与了光相关的调节过程,而叶片是植物进行光合作用以及光应激的主要器官,暗示着第Ⅱ亚族中的基因可能参与了光相关的调节,如光响应调控和叶绿素合成。
2.6 盐处理对毛果杨GATA 家族基因在各茎节表达的影响
为了研究PtrGATA 家族各基因是否在盐胁迫中具有一定的生物学功能,我们利用qRT-PCR 检测了各基因在盐胁迫下不同发育茎段中的表达变化情况,结果(见图6A)表明,PtrGATA5.1、PtrGA⁃TA5.2、PtrGATA7.2、PtrGATA10.2、PtrGATA17.1 和PtrGATA17.2 基因胁迫处理后表达量无明显变化;PtrGATA1.1、PtrGATA1.2、PtrGATA2.3、PtrGATA2.4、PtrGATA3.1、PtrGATA3.3、PtrGATA6.1、PtrGATA7.3和PtrGATA14.1 基因胁迫处理后表达量变化较小;PtrGATA4.1、PtrGATA5.4、PtrGATA5.6、PtrGATA9.1、PtrGATA10.1、 PtrGATA10.3、 PtrGATA19.1 和PtrGATAT.1 基因胁迫处理后在初生茎节表达量明显变化,其中PtrGATA10.1 表达量变化最为明显,是对照的6.4 倍(见图6B);PtrGATA1.3、PtrGA⁃TA2.2、PtrGATA3.2、PtrGATA4.2、PtrGATA5.5、PtrGA⁃TA6.2、PtrGATA8.1、PtrGATA8.2、PtrGATA13.1、PtrGA⁃TA14.2、PtrGATA18.1和PtrGATA18.2基因胁迫处理后在过渡茎节表达量明显变化,其中PtrGATA18.1盐胁迫处理后在茎部整体表达量变化最为明显,过度茎节最明显,是对照的35.6 倍(见图6C);PtrGATA2.1、PtrGATA5.3、PtrGATA7.1和PtrGATA7.4基因胁迫处理后在次生茎节表达量明显变化,其中PtrGATA5.3 表达量变化最为明显,是对照的18.2 倍(见图6D)。表明毛果杨GATA 基因可能通过复杂的机制参与植物盐胁迫适应过程。
3 讨论
随着植物基因组研究的深入,利用生物信息学技术,通过广泛的基因家族分析,奠定基因功能研究的基础,成为现今基因功能研究的热点。研究植物基因家族中基因的分布、基因结构与表达分析,对进一步研究其在植物生长发育及抗性响应中的功能具有重要意义。GATA 转录因子是一类广泛存在于真核生物中的转录因子,具有特殊的锌指结构,根据前人对拟南芥[7]、蓖麻[8]、苹果[9]、水稻[10]、番茄[11]和辣椒[12]等植物的研究发现,除水稻分为7 个亚族外,其他均分成4 个亚族,在光响应调控和叶绿素合成[13~16]、细胞分裂素响应[17~18]、花发育[19]和种子萌发[20]等多个生物学过程中扮演着重要角色。GATA 家族的大部分氨基酸位点在大部分成员中是高度保守的,这与本研究中毛果杨GATA 转录因子的家族成员的保守性,基本相符。
本研究对毛果杨GATA 家族39 个转录因子进行了系统的生物信息学分析。染色体定位分析表明,毛果杨GATA 转录因子不均匀的分布在15 条染色体上。根据系统进化树分析,毛果杨GATA 家族成员被分成了4 个亚家族,这一点与拟南芥[7]相同,可根据拟南芥各亚族GATA 基因功能来推测毛果杨相应GATA 基因的功能。基因结构和保守基序分析结果表明,在同一亚族的GATA 基因含有同类型的蛋白保守基序和相同数量的内含子,表明同一亚组的基因可能具有相似的功能和结构,而不同亚家族间存在的差异可能是进化中发生变化导致的。保守结构域分析发现,毛果杨GATA 家族在整体上具有较强的保守性,除了第Ⅳ亚家族外,其他亚族都含有共同的保守区域。组织表达特异性分析表明,第Ⅰ亚族、第Ⅲ亚族和第Ⅵ亚族中各个家族成员均在茎部相对表达量较高,第Ⅱ亚族中各家族成员基因组织偏爱性表达明显在叶部,说明同一亚族亲缘关系可能比较相近,在植物生长过程中发挥的功能可能相似。此外,GATA 基因在植物中普遍表达,在细胞的发育分化、生长增殖和分解凋亡等许多重要的生长发育过程中扮演着重要的角色。已有研究表明,一些与逆境相关的锌指蛋白基因,具有对植物的抗逆性调控的功能[24~26]。通过盐胁迫处理发现,GATA 基因家族成员在不同发育阶段的茎部表达存在明显差异;同时盐胁迫处理后,大部分基因表达量变化显著,其中PtrGATA10.1、PtrGATA18.1和PtrGATA5.3分别在盐胁迫后的初生茎节、过度茎节和次生茎节表达量显著性升高。这说明毛果杨GATA 家族基因可能参与杨树茎的发育与盐胁迫响应。
综上所述,本研究对鉴定出的39 个PtrGATA家族基因进行了染色体定位、理化性质、结构特征、进化关系以及表达模式分析,为今后进一步研究该家族基因奠定了一定基础。
