橡胶草E2泛素结合酶基因TkUBC2基因的克隆及其表达分析
2021-02-26王肖肖杨玉双聂秋海张继川刘实忠
王肖肖 覃 碧 杨玉双 聂秋海 张继川 刘实忠*
(1. 海南大学热带作物学院,海口 570228;2. 中国热带农业科学院橡胶研究所,海口 571101;3. 北京玲珑蒲公英科技发展有限公司,北京 110000;4. 北京化工大学材料科学与工程学院,北京 100029)
在逆境条件下,植物会激活多种信号通路来调节自身的生长发育,其中泛素介导的蛋白质降解途径在调控植物应对逆境胁迫过程中具有重要作用[1]。该途径通过泛素活化酶E1(ubiquitin acti⁃vating enzyme,E1)、泛素结合酶E2(ubiquitin-con⁃jugating enzyme,UBC/E2)和泛素连接酶E3(ubiqui⁃tin ligase,E3)多步反应实现。其中E2 酶起到“承上启下”的作用,接收从E1 活化的泛素蛋白,催化泛素蛋白直接与底物结合或与E3 结合。因此,E2在泛素蛋白酶途径(UPP)中发挥重要作用。
泛素结合酶E2 由一个高度保守的泛素结合结构域(UBCc)和末端组成,根据不同的末端可将UBC 基因家族分成4 个亚族[2~3],Ⅰ类只有UBCc结构域,可降解短命蛋白或泛素化异常蛋白;Ⅱ类具有UBCc 结构域和C-末端,后者可识别E3 和靶蛋白;Ⅲ类具有UBCc 结构域和N-末端;Ⅳ类除UBCc结构域外同时具有C-末端和N-末端,N-末端结构功能尚不清楚。末端序列的差异使得真核生物中E2 基因家族的功能具有多样性。E2 主要参与植物的胁迫响应、生长发育和DNA 损伤修复等生理过程[4]。植物的胁迫响应研究主要是干旱、高低温、盐胁迫以及激素胁迫研究。拟南芥AtUBC22 突变体表达了与植物防御相关的基因,表明UBC22在植物发育和胁迫反应中具有多种功能[5]。马铃薯StUBC12 在干旱和盐胁迫下上调表达,提高了植株的抗逆性[6]。大豆GmUBC13 基因在干旱、高盐、低温和ABA 等胁迫下表达量升高,在烟草中过表达GmUBC13 可以提高植株的抗旱性[7]。在拟南芥中过表达绿豆VrUBC1 基因可以提高植株对干旱胁迫的耐受性[8]。龙葵SorUBC 基因参与干旱、高温、低温、盐等非生物胁迫的早期响应[9]。在蒜介茄中,叶片和根中的SsUBC基因对非生物胁迫的响应不同,高温、干旱、镉、ABA 诱导叶片中SsUBC 基因上调表达,而在根中该基因在干旱胁迫下呈现下调表达趋势[10]。菜心BclUBE2基因参与低温胁迫的响应过程[11]。除此之外,E2还参与调控植物的生长发育过程,如花期、光周期、木质部形成等。荔枝LcUBC12 基因在烯效唑处理下对花穗发育具有负调控作用[12]。Xu 等发现在拟南芥中泛素结合酶UBC34以光依赖的方式调控蔗糖转运蛋白2(Sucrose Transporter 2,SUC2)的转化率,并调控SUC 提高植物的生产力[13]。Zheng 等[14]发现毛白杨泛素结合酶PtoUBC34 与木质素相关的转录抑制因子PtoMYB221 和Pto⁃MYB156 相互作用,参与转录后的调控,并发现PtoUBC34 受盐和热激诱导。在E2 基因家族中,RAD6/UBC2 蛋白的研究较为深入,大量研究表明,酵母UBC2 蛋白即RAD6 具有多种功能,在DNA 损伤修复、细胞周期、细胞增殖、染色质加工等过程发挥关键作用[15]。在人基因组中也鉴定到两个RAD6 同源基因HHR6A 和HHR6B,它们同样具有DNA 损伤修复的功能[16]。拟南芥中Rad6 同源基因有3 个,AtUBC1、AtUBC2、AtUBC3,前两者参与组蛋白H2B 的单泛素化并通过上调FLC 相关基因的表达抑制开花,在酵母实验中AtUBC2能够部分恢复酵母突变体对UV 照射的敏感性和降低其在高温条件下的生长速度[17~19]。Carsten 等发现一种SUMO-E2 酶UBC9 参与DNA 合成和修复[20]。水稻RE2 可以参与苯丙类化合物的合成代谢对UV 辐射 进行 防 护[21]。另 外,Song 等[22]和Nong等[23]的研究表明E2 基因可作为内参基因进行基因表达研究。
橡胶草(Taraxacum kok-saghyz Rodin)是原产于哈萨克斯坦和中国新疆的多年生草本植物,其根部富含优质的天然橡胶,是极具研究和开发潜力的产胶作物。目前,相关研究主要集中在天然橡胶合成相关基因的分子机制研究[24]、遗传转化[25]和组织培养等技术开发[26]。有关抗性基因的研究较少,而抗性基因的筛选和克隆将为橡胶草的遗传改良和抗性基因工程育种奠定基础。本研究克隆到橡胶草泛素结合酶TkUBC2基因,并对其基因与蛋白序列的结构特征和表达模式进行分析,为进一步研究橡胶草E2 基因的功能、选育抗逆品系提供理论依据。
1 材料与方法
1.1 实验材料
实验材料为橡胶草品系1151 组培苗,定植在实验室人工气候箱。在盛花期同时取植株的根、叶、花和花梗,用液氮速冻,保存于-80℃超低温冰箱,用于TkUBC2 的克隆和不同组织的表达分析。组培苗移栽在人工气候箱经水培培养2个月,取长势一致的植株将其根部分别浸泡在15%PEG6000、250 mmol·L-1甘 露 醇(Mannitol)、300 mmol·L-1NaCl、200 μmol·L-1茉莉酸甲酯(MeJA)、10 μmol·L-1脱落酸(ABA)、1%(v/v)乙烯利(ET)溶液中进行处理,在处理后0、6、12、24、48、72 h 分别取其叶片,样品用液氮速冻,用于非生物胁迫和激素诱导下基因的表达模式分析。组培苗放在紫外灯下进行UV(40 μW·cm-2)处理,分别于处理后0、0.5、2、6、12、24 h取其叶片,用于UV辐射下基因的表达模式分析。
1.2 实验方法
1.2.1总RNA的提取和cDNA的合成
橡胶草不同组织(根、叶、花、花梗)和处理样品(叶片)的总RNA 采用多糖多酚植物总RNA 提取试剂盒(天根生化科技(北京)有限公司)提取,用1%琼脂糖凝胶电泳检测浓度与纯度,采用反转录试剂盒(Thermo Scientific RevertAid First Strand cDNA Synthesis Kit,购于赛默飞世尔公司)合成得到cDNA,具体步骤参照说明书进行。
1.2.2基因编码区的克隆
用拟南芥AtUBC2(Arabidopsis thaliana,NP_565289)的氨基酸序列对橡胶草转录组数据进行tBlastn 搜索,获得橡胶草UBC2 的DNA 序列,参照转录组搜索获得的序列设计引物(见表1),以橡胶草品系1151 叶片的cDNA 为模板,用Premix TaqTM(Ex TaqTMVersion 2.0)(Takara)进行PCR 扩增,反应体系:cDNA 模板1 μL(30 ng),2×Premix Taq Mix 10 μL,正向和反向引物(10 μmol·L-1)各0.5 μL,ddH2O 8 μL,扩增程序为95℃预变性30 s;94℃变性30 s,55℃退火30 s,72℃延伸1 min,共35 个循环;最后72℃延伸10 min。扩增产物用1%琼脂糖凝胶电泳后,目的片段采用凝胶回收试剂盒(OMEGA,E.Z.N.A.TMGel Extraction Kit)进行回收,并连接到pMD-18T(Takara)载体上转化大肠杆菌DH5α进行克隆,菌落PCR检测后挑取阳性单克隆送广州艾基生物技术有限公司测序验证。
1.2 .3 生物信息学分析
通过NCBI 网站的ORF Fider(https://www.nc⁃bi.nlm.nih.gov/orffinder/)在线分析确定TkUBC2 基因的ORF及其编码氨基酸序列,在NCBI非冗余蛋白数据库中采用BLASTp(https://blast.ncbi.nlm.nih.gov/Blast.cgi)搜索TkUBC2 蛋白序列的同源性序列。采用DNAMAN 软件对多序列同源性和相似性进行分析。采用MEGA7 软件构建系统进化树。采用ProtParam 工具(https://web.expasy.org/protparam/)对TkUBC2 的蛋白质进行理化特性分析。采用SignalP-5.0 Server、TMHMM Serverv.2.0、NetPhos 3.1 Server 和PSIPRED V4.0 等 在 线 分 析 工具分别分析预测TkUBC2 的信号肽、跨膜蛋白结构、磷酸化位点、亚细胞定位、二级结构、三级结构和保守结构域。
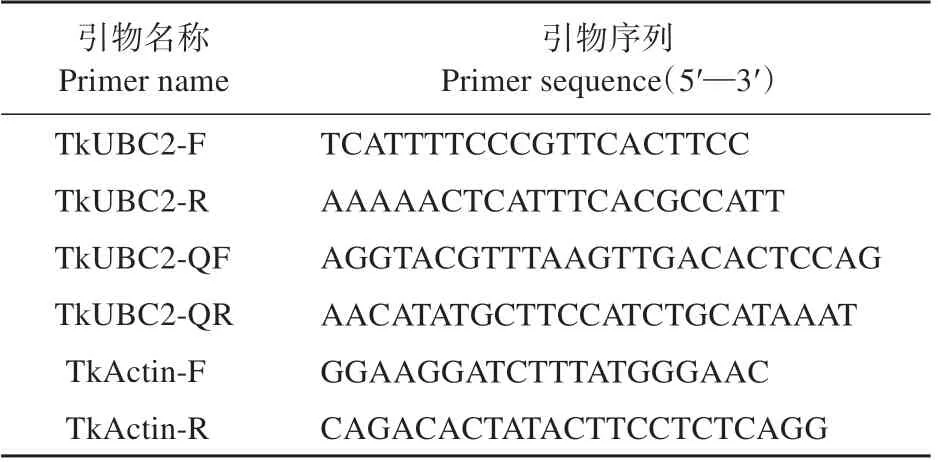
表1 TkUBC2全长扩增和qRT-PCR分析引物序列Table 1 Primer sequences used in cloning the full-length of TkUBC2 and qRT-PCR analysis
1.2.4荧光定量qRT-PCR分析
根据TkUBC2 基因序列设计qRT-PCR 引物(见表1),以橡胶草TkActin 基因为内参,采用SYBR Green 法进行qRT-PCR 扩增,反应体系为2×SYBR Premix Ex TaqTMMix(Takara)10 μL,正向与反向引物(10 μmol·L-1)各0.4 μL,cDNA 1 μL(30 ng),ddH2O 8.2 μL,在荧光定量PCR 仪(CFX96 TouchTMReal-Time PCR Detection System,Bio-Rad)上进行扩增,扩增程序为95℃预变性30 s;95℃变性5 s,60℃退火30 s,共40个循环。根据CT值用2-△△CT计算目的基因的相对表达量。
1.2.5数据统计分析
采用Excel 进行数据整理分析,用SAS 软件(The SAS System for Windows V8)进 行 单因 素ANOVA 完全随机(均衡)分析差异显著性,采用Origin Pro 2019(Origin Lab Corporation,Massachu⁃setts,USA)软件进行作图。
2 结果与分析
2.1 TkUBC2的克隆与序列分析
以橡胶草1151 叶片为材料,提取RNA,反转录成cDNA,以引物TkUBC2-F 和TkUBC2-R 进行PCR 扩增得到目的片段,经克隆后测序验证,获得橡胶草TKUBC2 基因序列,长度为706 bp,包含一个459 bp的开放阅读框(ORF),编码152个氨基酸(见图1)。采用ProtParam 工具对其编码蛋白分析发现,TkUBC2 蛋白相对分子量为37.97 kDa,等电点5.23,亲水性0.805,不稳定指数为33.03,表明该蛋白是稳定性蛋白质。通过SignalP-5.0 Server、TMHMM Serverv.2.0、NetPhos 3.1 Server和PSIPRED V4.0在线分析,表明TkUBC2 无信号肽和跨膜结构,含有7 个丝氨酸(Ser)磷酸化位点,5 个苏氨酸(Thr)磷酸化位点,4 个酪氨酸(Tyr)磷酸化位点,可能定位于细胞核中。通过蛋白质结构分析发现,二级结构有38.82%α-螺旋,3.29%β-转角,17.76%延长链,40.13%无规则卷曲(见图2A),表明TkUBC2 的二级结构主要是α-螺旋和无规则卷曲;由二级结构进一步卷曲、折叠形成三级结构(见图2B)。保守结构域预测如图2C所示,该蛋白有一个高度保守的泛素结合结构域(UBCc),属于泛素结合酶E2家族成员。
2.2 同源性及进化树分析
将TkUBC2的氨基酸序列输入NCBI蛋白数据库进行BLASTp 搜索,找到其他物种的同源蛋白,通过DNAMAN 进行多重序列比对,结果表明,UBC2 在不同物种间具有很高的同源性,橡胶草TkUBC2 与 莴 苣LsUBC2(Lactuca sativa,XP_023744018)、绒毛烟草NtUBC2(Nicotiana tomento⁃siformis,XP_009589859)的同源性都达到99.34%;其次是与洋蓟CsUBC2(Cynara cardunculus var.Scolymus,XP_024978185)、向日葵HaUBC2(Heli⁃anthus annuus,XP_022012684)的 同 源 性 达 到98.68%;与 本 生 烟NbUBC2(Nicotiana benthami⁃ana,AOF39395)、大豆GmUBC2(Glycine max,NP_001235621)、拟南芥AtUBC2(Arabidopsis thaliana,NP_565289)的同源性分别为98.04%、98.03%、96.05%(见图3A)。进一步采用MEGA7 软件构建系统发育树发现,8个UBC蛋白可以划分为两个分支,其中NtUBC2、CsUBC2、NbUBC2、GmUBC2 和AtUBC2 聚在同一个分支上,而橡胶草TkUBC2 与莴苣LsUBC2、向日葵HaUBC2聚在另一分支上(见图3B),它们在分类上同属于菊科植物,表明UBC2在同科植物中的保守性更高。
2.3 橡胶草TkUBC2基因的表达模式分析
为了揭示TkUBC2 基因的功能,我们采用qRT-PCR 技术对其表达模式进行系统分析。如图4A 所示,TkUBC2 在橡胶草盛花期的叶、根、花和花梗中均有表达,且不同组织中的表达水平差异显著,其中花梗中的表达量最低,叶中的表达量最高,是花梗的3倍。如图4B所示,TkUBC2在UV辐射处理下呈现明显地上调表达,12 h 的表达量达到未处理对照的2 倍。结果表明TkUBC2 与橡胶草适应UV 辐射胁迫有关。进一步地,以不同非生物胁迫和激素处理后橡胶草叶片为材料,系统地分析TkUBC2 在胁迫响应和激素信号传导中的作用。在PEG6000 模拟干旱胁迫、甘露醇介导的渗透压胁迫以及植物激素MeJA 处理后TkUBC2基因显著下调表达,且均在处理后12 h 降到最低水平,其中PEG6000 处理下降了70%、甘露醇处理下降了60%、MeJA 处理下降了50%,然后逐步恢复到未处理时的表达水平。在NaCl 高盐胁迫下TkUBC2 基因表达量在12 h 上调1.5 倍,在48 h 恢复到未处理时的表达水平,但在72 h 又上调1.5倍。在ABA 和ET 诱导下,TkUBC2 基因表达量先下降后上升,分别在处理后12 h 和24 h 下降至最低值,同时都是在72 h 显著升高且达到最高值(见图5)以上结果表明,TkUBC2基因广泛地参与橡胶草对干旱、渗透压、高盐胁迫的响应以及乙烯、茉莉酸和ABA信号转导过程。
3 讨论
泛素结合酶UBC/E2 是泛素—蛋白酶体途径(UPP)的关键酶。本研究从橡胶草中克隆的TkUBC2 基因,其编码的氨基酸序列与同为菊科的莴苣和向日葵高度相似,氨基酸序列间的相似性达99.34%和98.68%,表明E2 酶蛋白在物种间具有很高的保守性。蛋白结构域分析发现,TkUBC2只有一个UBCc结构域,属于UBC基因家族中的Ⅰ亚族,可直接降解短命蛋白或已被泛素化的异常蛋白。通过qRT-PCR 技术分析TkUBC2 基因的表达模式,结果显示,该基因在各组织中均有表达且差异显著,其中花梗中的表达量最低,叶中的表达量最高。尹丽娟等[27]发现小麦TaE2 基因在小麦根、茎、叶和种子中均有表达,且表达量一致;蔡佳文等[9]发现龙葵SorUBC 基因在龙葵根、茎、叶、花和果实中均有表达,在叶和果实中表达量最高;曾小玲等[11]发现菜心BclUBE2 基因在根、茎、叶和叶柄中也均有表达,在叶中的表达量最高;安红强等[28]发现铁皮石斛DoUBC24基因在叶中得表达量最低。这些结果表明,泛素结合酶E2基因在不同器官中均有表达,不同物种间存在组织表达特异性。
大量的研究表明,泛素结合酶E2 基因与植物的抗逆性相关。比如,小麦TaE2 基因在干旱、高盐和ABA 胁迫下均上调表达,参与逆境胁迫响应[27];龙葵SorUBC 基因在高低温和干旱等非生物胁迫下呈现下调表达趋势[9];大豆GmUBC2基因受干旱和盐胁迫后上调表达,在拟南芥中过表达GmUBC2 可提高非生物胁迫响应基因的表达从而提高植株对干旱和盐胁迫的抗性[29];花生AhUBC2基因转入拟南芥后受PEG6000、高盐、ABA 和低温胁迫诱导,且该基因能通过不依赖ABA 信号通路提高植株的耐旱性[30];甜瓜CmUBC 基因在干旱和盐胁迫下转录水平升高[31]。本研究中,橡胶草在PEG6000 模拟干旱、甘露醇介导的渗透压胁迫处理后,TkUBC2 基因显著下调表达;而NaCl 高盐胁迫处理下,TkUBC2上调表达,表明TkUBC2对不同胁迫的响应机制不同,胁迫条件下TkUBC2蛋白可能通过对不同响应蛋白的泛素化修饰间接参与植物对胁迫条件的反应过程。TkUBC2 经ABA 诱导的早期(0~12 h)表达模式与其在干旱、渗透压胁迫下的表达模式类似,而在ABA 诱导的后期(48~72 h)表达模式与其在高盐胁迫下的表达模式类似,表明TkUBC2 受ABA 诱导且能不依赖于ABA信号通路参与逆境胁迫响应过程,其参与调控的信号通路通过修饰的靶蛋白决定。除此之外,Me⁃JA 可诱导TkUBC2 下调表达,而ET 诱导下,TkUBC2 表达量先下降后上升。UV 辐射会诱导植物产生一些与胁迫相关的生理反应,如DNA 损伤[32]。UV-B和UV-C都会直接与DNA碱基反应产生光离子,阻碍DNA 复制和转录[33~35]。在UV 辐射处理下,TkUBC2 呈现上调表达,在12 h 开始上调表达,说明该基因对橡胶草DNA 损伤修复具有正调控作用。赖燕等[36]在辣椒中的研究表明E2 基因与辣椒适应UV-B 照射胁迫有关。橡胶树HbRad6 在DNA 损伤剂H2O2处理下呈现上调表达并且能够部分恢复酵母rad6 突变体对UV 的敏感性,表明其参与DNA 损伤修复过程[37]。本研究结果表明TkUBC2 广泛参与橡胶草对多种胁迫响应和激素信号转导过程,对DNA 损伤修复具有积极的作用,为进一步阐明TkUBC2在橡胶草抗逆反应和激素信号转导过程中的功能打下基础。
