果树器官脱落的研究进展
2020-09-24郝紫微吴潇刘哲谢智华张绍铃王鹏
郝紫微 吴潇 刘哲 谢智华 张绍铃 王鹏

摘要:器官脱落是植物通过可控方式将某一器官与母体分离的过程,在植物界普遍发生,也是果树重要的农艺性状。器官脱落发生的位置称为离区,离区的发育是多种基因参与的精细而复杂的调控过程。本文通过总结近年来对果树器官脱落的研究报道,在生理生化和分子机制方面阐述了果树器官脱落最新研究进展,以期提高对器官脱落的理解,为进一步深入研究果树脱落机理提供思路与参考。
关键词:果树;器官脱落;调控机制;研究进展;工作假说
中图分类号:S660.1
文献标志码:A
文章编号:1002-1302(2020)16-0058-09
器官脱落是植物界普遍存在的生理现象。果树中常见花、果、叶和枝的脱落。这些器官的非正常脱落最终均会影响果品的质量和产量,甚至导致严重的经济损失。张全军等研究发现,砂梨的返花返青现象与梨树早期落叶密切相关[1-2],严重影响翌年果实的产量和品质。所以在果树生产中,实时控制好花果的脱落不仅能减少疏花疏果的人工费用支出,还能提高果实品质,增加收入。
在脱落的起始阶段,脱落器官的基部分化出离区,离区会进一步分化产生离层和保护层[3]。离层是器官脱落母体的位置,植物器官的离层因物种和器官存在差异,一般由5~50层细胞组成[4]。而果树中的离层细胞一般有5~17层[5-6],其中酸樱桃的果实与果柄的离层细胞有5~8层,柑橘的果实和果柄,叶柄、叶片分别有10~17、15层,日本柿花萼与花梗的脱落层由7层细胞组成。
本文从影响果树器官脱落的内外因素及相关基因进行阐述,并对器官脱落提出工作假说及展望,希望为进一步深入研究脱落机理,改良果树农艺性状提供理论依据。
1 影响器官脱落的内外因素
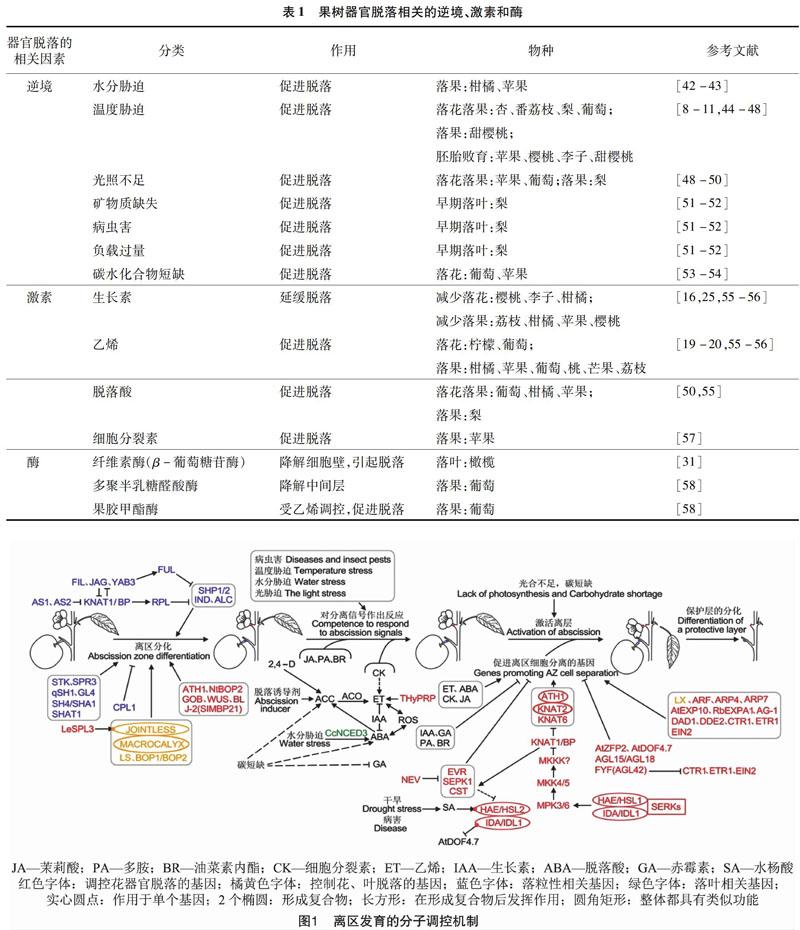
果树器官脱落是一个由多种因素协调控制的复杂的生理过程,可分为衰老或成熟引起的正常脱落、植物自身生理活动引起的生理脱落和逆境条件引起的胁迫脱落3种类型[7]。逆境条件、植物内源激素及相关的酶均可以影响果树器官脱落。其中,逆境条件产生的脱落最容易发生且危害严重,也是引起果树器官非正常脱落的主要原因,包括水分胁迫、温度胁迫、光照不足、矿物质与碳水化合物的缺失以及病虫害等(表1)。这些不利的外界条件通过影响果树体内的生理机能,尤其是影响果树体内的激素平衡和糖代谢2个过程,从而引起果树器官的脱落。在水涝条件下,根系进行无氧呼吸产生乙烯运输到果柄离层处造成落果;光照不足、营养物质的缺失以及病虫害的发生影响树体的养分供应,造成树势衰弱;反常的气候条件会使果树生长受损,发生早期落叶;在温度胁迫下,苹果[8]、樱桃[9]、李子[10]、甜樱桃[11]等还表现出胚胎败育等现象。这些非正常脱落对果树生长和生产造成不利影响,导致果实产量和品质的下降。
大量研究表明,激素也是调节果树器官脱落的重要因素之一。脱落进程的启动,需要离区感受脱落信号,而植物激素可以作为信号调节剂影响植物器官脱落[12-15]。生长素(IAA)与离层形成直接相关,具有延缓脱落的作用[16],其对器官脱落的影响程度与浓度、时间、部位相关。“生长素梯度”学说指出决定脱落的是生长素相对浓度[17]。生长素相对浓度高时,即远轴端生长素含量高于近轴端时,离層对乙烯的敏感性小,器官不脱落;相对浓度低时,离层细胞对乙烯的敏感性增加,启动脱落进程,加速离层形成,引起器官脱落[7]。Xie等发现,IAA以浓度依赖的方式抑制柑橘果实脱落,而花柄离区中的IAA则可以直接抑制乙烯的生物合成和信号传导,从而抑制脱落[18]。另外,有研究报道表明,生长素刺激了乙烯的合成,参与衰老过程。例如,外源施用IAA加速了乙烯产生和康乃馨花的萎蔫[19],2,4-二氯苯氧乙酸(2,4-D)引起花柱、子房和花瓣中ACC(乙烯前体)合成酶基因的表达[20]。此外,生长素抑制或刺激乙烯生物合成依赖于多种因素,包括植物物种、器官和发育阶段等,并且显示出不同的调节机制和生物学功能[21]。目前在柑橘、葡萄等果树中较多研究表明,在叶片和花果自然脱落、幼胚败育、发生病虫害时,体内会产生大量乙烯,促进了果树器官的脱落[22](表1)。另外,乙烯可以反作用于生长素,可以通过上调番茄外向转运体基因SlPIN1(Solanum lycopersicum Pin-Formed1)降低离区生长素。芒果果实离区感知乙烯后,花柄中的极性生长素运输能力减弱,随后乙烯受体上调,蔗糖含量降低导致果实脱落[23-24]。此外,乙烯还可以通过调节一系列主要细胞过程促进荔枝果实脱落,包括乙烯生物合成和信号传导、生长素转运和信号传导、转录调节、蛋白质泛素化、活性氧反应,钙信号传导和细胞壁修饰等过程[25]。脱落酸(ABA)是植物在不同发育阶段非生物胁迫抵抗机制的激活剂和调节因子,可以加速植物器官脱落进程[26]。但这一作用会受到乙烯和生长素等其他植物激素的影响。柑橘果实经IAA处理后,编码脱落酸(ABA)生物合成酶基因Cs2g03270和ABA降解酶基因Cs8g18780在脱落过程中分别被显著地下调和上调,表明了生长素与ABA存在拮抗关系(图1)。目前,有大量研究表明,脱落酸可以促进葡萄、柑橘、苹果等果树落花落果(表1)。其他激素中,赤霉素(GA)和茉莉酸(JA)可以加速脱落,细胞分裂素(CK)、油菜素内脂(BR)为脱落的抑制剂。在柑橘中,与合成JA相关的3个基因(Cs7g12890、Cs5g28310、Cs2g21000)和与BR信号相关的5个基因(Cs9g18950、Cs9g15900、Cs6g20340、Cs5g08340、Cs4g02270)都受到IAA的影响[18]。除了一些激素外,植物生长发育调节物质多胺(PA)也为脱落的抑制剂。并且多胺含量与梨树二次开花过程中花芽萌芽率之间存在显著相关关系。在发生环境胁迫时,其含量增加可以使乙烯含量减少,延缓植物的衰老[27],但它们的具体作用机制尚不清楚。
此外,器官脱落还受到一些酶的调控。有研究指出,果树中纤维素酶和果胶酶等水解酶活性增强,导致细胞壁松动,促进器官脱落(表1)。这些水解酶参与脱落区的细胞壁和中间层的降解,并且乙烯可以促进水解酶的生成,最终导致器官脱落[28]。这也说明激素可以促进一些水解酶合成,进而调控器官脱落。其中纤维素酶中的β-葡萄糖苷酶(BG)与器官脱落相关性较大,桃和柑橘果实离区中乙烯的增加能够诱导该酶含量升高,从而降解细胞壁,引起果实脱落。Qi等发现,将梨萼片脱落基因PsJOINTLESS过表达在转基因番茄中,编码纤维素酶基因CEL2上调且纤维素酶活性显著增加,引起了花柄脱落[29]。Yang等研究表明,使用H2O2清除剂可以抑制纤维素酶活性的增加,减少龙眼果实的脱落[30]。另外,Goldental等利用乙烯利处理橄榄叶、果实、花梗,发现仅在叶离区中纤维素酶基因上调[31],说明纤维素酶在脱落过程中具有组织表达特异性。果胶酶具有分解细胞壁间果胶质的功能,主要包括多聚半乳糖醛酸酶(PG)和果胶甲酯酶(PME)2种酶。PG是参与中间层降解的关键酶,多位于细胞发生分离的区域,乙烯能够诱导PG的生成,其活性的升高与离区脱落进程中强度相一致。
Chersicola等用1-甲基环丙烯(1-MCP)(乙烯抑制剂)处理番茄,发现1-MCP可以通过抑制PG的表达阻止细胞的分离[32]。Srivignesh等报道表明,番茄的3个PG基因(TAPG1、TAPG2和TAPG4)参与离区的形成[33]。PME基因的表达同样受乙烯的调控,但其在器官脱落方面的具体作用存在争议。较多研究结果表明,果胶甲酯酶对植物器官的脱落有促进作用[12,34],但也有不同观点,如该酶在柑橘叶柄和龙眼果实中并没发现具有调控脱落功能[35-36]。过氧化物酶(peroxidase,POD)是植物呼吸过程中呼吸末端氧化酶之一,对植物器官脱落有一定的促进作用。其调控机制主要有2点,一是POD参与生长素的氧化进程,降低离区生长素的水平,间接促进脱落;二是POD促进离区伤痕的木质化,进而促进脱落。其他对器官脱落起作用的几丁质酶(chitinase)和苯丙氨酸解氨酶(phenylalanine ammonialyase,PAL)等酶类,对于果树器官脱落后伤口的保护和脱落断面的木质化有重要作用[12,37]。
有研究指出,离区细胞的细胞质溶液碱性随着脱落过程的进行变强,且乙烯抑制剂的应用可以阻止胞质碱化。因此,pH值的改变可能为细胞壁降解酶提供了更好的细胞环境,影响其活性,从而影响脱落[38]。
此外,细胞凋亡(programmed cell death,PCD)过程是离层形成的重要调控机制。Bar-Dror等的研究结果表明,核糖核酸酶基因是影响器官脱落的主要基因,该基因参与了PCD过程[39]。在番茄花和叶脱落后期的离层细胞中观察到PCD的一些典型特征,包括细胞活力丧失,核形态改变,DNA片段化,活性氧和酶活性水平升高,以及PCD相关基因的表达量升高,这表明PCD与脱落密切相关。但二者之间潜在的调节机制还存在争议,需进一步研究。活性氧(reactive oxygen species,ROS)是植物PCD的关键影响因子,与发育过程和环境应激反应有关[40],相比于正常细胞,NADPH氧化酶基因RBOH1在离层细胞表达显著升高,ROS含量显著提高[41]。较低剂量的ROS可以提高植物的抗逆性,而较高浓度则可能导致细胞程序性死亡。研究表明,ROS可以介导碳水化合物胁迫诱导的龙眼果实的脱落[30]。不同的胁迫因子在逆境胁迫条件下通过其他信号和激素的作用,直接或间接促进ROS的产生和PCD的开始,在极端情况下甚至导致组织坏死。并且ROS含量的变化会影响PCD进程,从而影响离区发育。
2 果树器官脱落相关基因的研究
进行果树器官脱落的分子机制研究,可为进一步了解器官脱落发生过程和对脱落进行调控提供更可靠的理论依据。目前,在模式植物拟南芥中已报道的引起脱落的基因中,在桃、柑橘、苹果等11种果树中研究了其中14类,包括7类与花器官脱落相关和7类与果实脱落相关(表2)。
目前J(Jointless)是离区发育相关的研究较多的基因之一[33,59-61],J和J-2(Jointless-2)是非等位的2个MADS-box基因,且J是J-2的上位基因[62]。J的突变可以抑制花序分生组织中合轴的发育和离区的产生,维持花序分生组织的正常生长,并且对于胚珠和种子发育、果实形状也有重要作用[59,63]。番茄jointless突变体不能形成离区,因此不发生果实或花的脱落[59]。将梨的J基因转入到番茄植株中,发现该基因促进花柄脱落,花柄变细,离层细胞数目增加、细胞变小、破裂、排列无序,花柄离区纤维酶的活性升高,细胞壁水解酶的基因和诱导编码脱落相关的转录因子表达量上升[64]。IDA(Inflorescence Deficient In Abscission)是模式植物拟南芥中促进花器官脱落的主要基因。最新的研究表明,在IDA与受体激酶HAE/HSL2(Haesa/Haesa-Like2)结合后,SERKs(Somatic Embryognesis Receptor-Like Kinase)能够与HAE/HSL2形成复合物,从而调节花器官脱落[65](图1)。目前,IDA基因在梨、柑橘和荔枝中均有报道研究。PsIDA可以促进梨萼片脱落[64]。在拟南芥过表达荔枝或柑橘的IDA基因引起拟南芥的早期落花[66]。柑橘中的CitIDA3还可以明显抑制拟南芥突变体ida-2花的脱落[57],说明该基因功能比较保守。此外,St等在杨树(双子叶树)和棕榈(单子叶树)研究了IDA-HAE/HSL2脱落信号通路,发现IDA-HSL在多个物种的不同离区中均有表达,为该通路的功能保守性提供进一步的证据[67]。这些结果表明IDA-HAE/HSL2脱落信号通路在不同植物类群的高等植物中均起着重要作用。其他几类基因如:KNAT1/BP(Knotted-Like From Arabidopsis thaliana/Bervipedicellus)、STK(Seedstick)等基因[68-70]如何参与影响果树生长发育过程,如何影响器官脱落还需进一步研究。FUL(Fruitfull)基因不仅可以抑制苹果和桃离区发育,还具有促進果实成熟和提高花器官数量的功能[71]。此外,与激素相关的部分基因也调节了果树器官的脱落。杜艺在研究荔枝生理落果问题时,发现了13个IAA和12个ABA相关基因分别参与荔枝幼果和成熟果脱落过程[72]。Li等进行了荔枝果实全基因组数字转录丰度分析挖掘了控制脱落2 730个候选基因,其中195个基因与激素相关[25]。干旱胁迫诱导柑橘ABA生物合成基因CcNCED3(9-Cis-Epoxycarotenoid Dioxygenase)的表达,促进根中ABA合成,进而使叶片产生乙烯,导致叶片脱落。
3 器官脱落的工作假说
植物器官脱落大致分为4个阶段(图1)。第1阶段为离区的分化,第2阶段为离区根据外界刺激或发育信号作出反应同时启动脱落进程,第3阶段和第4阶段部分重叠,为离层细胞的激活,开始器官的脱落和保护层的形成。目前对模式植物拟南芥和番茄的器官脱落研究较多,已形成简单的基因调控网络。
第1阶段中,SHP1(Shatterproof1)、SHP2(Shatterproof2)、IND(Indehiscent)和ALC(Alcatraz)在拟南芥角果开裂的离区发育过程中起着重要功能[90]。AP1(Apetala1)的同源基因FUL(Fruitfull),以及RPL(Replumless)也参与角果开裂发育。FIL(Filamentous)、JAG(Jagged)、YAB3(Yabby3)、AS1/AS2(Asymmetric Leaves)和KNAT1/BP等部分基因形成一个复杂的调控网络共同控制角果离层的发育。根据已报道的FUL基因在草莓、木瓜、苹果、桃和RPL基因在龙眼中的功能,推测这2个基因可能通过抑制IND、SHP和ALC等下游基因的表达,进而抑制果实离区发育。在荔枝和桃中报道了基因FIL、YAB3及KNAT1/BP参与离区发育。这些基因功能较为保守,很可能在其他果树中也存在相似的基因调控网络参与离区发育。
落粒性是粮食作物的栽培育种的一个重要农艺性状。在脱落发生的第1阶段,促进离区分化的SH4/SHA1(Shattering1)、qSH1(Seed Shattering in Chromosome 1)、SHAT1(Shattering Abortion 1)基因,抑制离区形成的OsCPL1(C-Terminal Domain Phosphatase-Like 1)基因在作物落粒性的研究中取得了一定的進展。水稻qSH1基因与拟南芥RPL和FUL序列相似性较高,与长角果的离区的分化相关。SHAT1是拟南芥AP2蛋白的水稻直系同源物,参与花发育的调控[57]。这些结果表明,控制拟南芥角果发育的基因在其他物种中也可能具有同样的功能。MACROCALYX、LeSPL3(Squamosa Promoter Binding Protein-Like)可以促进花柄离区的分化。但二者调控途径不一样,MACROCALYX与J形成复合物[58]发挥功能,而LeSPL3在J的上游发挥作用[91]。此外,研究表明LS(Lateral Suppressor)、CEL2促进脱落,而BL(Blind)、TAPG2(Tomato Abscission-Related Polygalac-Turonases 2)、TAPG4可以抑制脱落[91-92]。Roldan等研究发现,LS、BL、LeWUS(Wushel)和GOB(Goblet)与离区形成相关[62]。因此,对于果树离区分化阶段,可参照其他作物中脱落相关基因进行研究。
根据发育进程与环境的刺激,离区发育进入第2阶段。病虫害、温度胁迫、水分胁迫等逆境环境以及激素的刺激会引发植物开始脱落。乙烯、脱落酸、细胞分裂素、茉莉酸促进脱落,生长素、赤霉素、油菜素内酯、多胺抑制脱落发生。调控各激素的基因在此阶段变得活跃,发挥重要作用,直接或间接地引起乙烯的变化而影响脱落。如水分胁迫引起柑橘ABA生物合成基因CcNCED3表达,促进乙烯产生,导致叶片脱落。而沉默THyPRP(Tomato Hybrid Proline-rich Protein)可以减少乙烯的生物合成[93]。使用2,4-D和脱落诱导剂可以促进乙烯前体ACC的产生,进而促进器官脱落。
第3和第4阶段表明发生的脱落现象,已经明确的花器官脱落信号传导途径是BOPl/BOP2(Blade On Petiole)→IDA→HAE/HSL2→MPK3/6→MKK4/MKK5→KNAT→花器官脱落,AtDOF4.7在IDA上游发挥作用。IDA基因在梨、柑橘和荔枝果树中均可以促进花器官脱落,该基因参与的花器官脱落信号传导途径在多个物种均有报道,在果树中很可能存在着相似的调控网络。另外,干旱或病害的发生会促进水杨酸的产生,诱导HAE/HSL2的表达。与生长素相关的ARF和SlPIN1等基因可以调控番茄脱落。ARF基因参与番茄离区发育和完成脱落进程[75],抑制SlPIN1的表达,加速番茄花瓣的脱落[23]。ARF基因在葡萄和苹果中参与离区发育已经被报道,但在果树中未发现SlPIN1及其同源物。因此SlPIN1基因在果树中可能不参与离区过程。此外,乙烯相关基因在拟南芥和番茄的脱落调控中存在差异,如ETR1、ETR2和ETR3的突变延迟了拟南芥角果的脱落,但在番茄中只有ETR1的同源基因neverripe突变延迟果实脱落[94-95]。根据该结果推测在果树器官脱落的研究中,乙烯相关基因的作用会依据物种的不同存在差异。
4 展望
植物器官脱落在植物基础理论研究和农业生产方面都具有重要意义。拟南芥、番茄等模式植物遗传背景清楚,繁殖快,容易转化,为器官脱落研究提供了很多便利条件。果树遗传转化体系比较困难,通过克隆果树相关基因转入到模式植物中,进行异源转化对果树器官脱落研究有重要的参考意义。果树植物中仅梨、柑橘、苹果、桃和荔枝等主要果树中的单个脱落相关基因功能研究较多,但是还缺乏完整的分子调控网络,需要进一步扩大果树种类,深入挖掘离区发育遗传机制。
在果树生产实践过程中,叶片脱落是一个重要问题。育种家和分子生物学家应共同努力,了解每个果树物种的发展规律,使用分子手段结合传统育种,以及形态学和生理学研究,提高对果树不同时期不同器官脱落的理解,应用更恰当的管理技术,提高果品的质量和产量,防止不必要的脱落。例如开发保叶剂、乙烯抑制剂、生长素合成剂以及人工控制下的适度脱落。目前在生产中疏花疏果剂已应用较多,但仍然需进一步细化其激素浓度配比以及喷施时间。在大多情况下,早期芽或果实的适度脱落会改善根的发育,使植物更有活力,中期适当疏花疏果能够防止“大小年”现象,获得更高品质的果品。国内外学者虽已在植物脱落机制方面进行了大量的研究,但仍有许多科学问题需要进一步解决。例如在植物激素对脱落的调控中,只有乙烯调控机制研究得较为深入,生长素、脱落酸等其他植物激素的作用部位和分子调控机制还不清楚[96]。另外,研究者虽然对脱落过程中细胞壁水解酶进行了大量的研究,但其他酶类包括过氧化物酶、几丁质酶、苯丙氨酸解氨酶等在脱落过程中的具体作用方式并不明确,不同果树对脱落起主要作用的酶也并不一致。今后对每个果树激素的调控作用、激素之间的调控网络以及酶在不同植物中的具体作用机制和它们之间的协同作用需要做进一步深入研究。
另外有关离区发育和器官脱落的分子机制应当在果树中进行更加广泛、深入的研究。例如目前柑橘落花落果机制尚不清楚,梨早期落叶的分子机制还未完全解析[51,97]。PsJ增强了脱落相关基因的表达,但这些基因之间的相互作用并不清晰,具体的基因调控网络需进一步确定。
在柑橘[98]、梨[99]、苹果[100]、草莓等[101]部分果树中基因编辑和遗传转化技术的日益完善以及基因组学与蛋白质组学的发展,为研究果树器官脱落的分子机制提供了强有力的工具,可以进一步阐明果树器官的脱落机理,解析果树器官脱落的特异性机制。这不仅对基础研究具有重要意義,在新品种培育和产业技术研发方面也将有重要的推动作用。
参考文献:
[1]张全军,李文贵,钟必凤,等. 南方砂梨返花返青发生特性及其与早期落叶的关系[J]. 西南农业学报,2013,26(3):1160-1163.
[2]张全军. 砂梨二次花发生的生理特征及基因表达谱分析[D]. 南京:南京农业大学,2015:2-14.
[3]韩 静,王幼群,王晓理. 植物器官脱落的机制及其研究进展[J]. 植物学通报,1999,16(4):405-410.
[4]刘 春. 月季花瓣脱落转录组分析及RhIAA16基因响应脱落的功能分析[D]. 北京:中国农业大学,2016:1-5.
[5]郭凯斌,李 升,乔 光. 大樱桃生理落花落果特性及调控机制研究进展[J]. 种子,2018,37(10):50-56.
[6]Li X,Kitajima A,Kataoka K,et al. Anatomical observations of the citrus fruit abscission zone and morphological changes of the cells during secondary physiological fruit drop[J]. The Horticulture Journal,2017,86(4):447-455.
[7]蔡庆生. 植物生理学[M]. 北京:中国农业大学出版社,2011:275-277.
[8]Simons R K. Tissue response of young developing apple fruits to freeze injury[J]. Journal of the American Society for Horticultural Science,1969,94:376-382.
[9]Stsser R,Anvari S. On the senescence of ovules in cherries[J]. Scientia Horticulturae,1982,16(1):29-38.
[10]Cerovic R,Ruzic D,Micic N. Viability of plum ovules at different temperatures[J]. Annals of Applied Biology,2000,137(1):53-59.
[11]Hedhly A,Hormaza J I,Herrero M. Warm temperatures at bloom reduce fruit set in sweet cherry[J]. Journal of Applied Botany and Food Quality,2007,81(2),158-164.
[12]高欣欣,刘少春,张跃彬,等. 植物器官脱落相关激素和酶的研究进展[J]. 中国农学通报,2013,29(33):17-21.
[13]Taesakul P,Siriphanich J,van Doorn W G. Two abscission zones proximal to Lansium domesticum fruit:one more sensitive to exogenous ethylene than the other[J]. Frontiers in Plant Science,2015,6(3):264.
[14]Jin X. The role of auxin in abscission of organs and tissues[D]. Sweden:Acta Universitatis Agriculturae Sueciae,2015:11-20.
[15]Domingos S,Scafidi P,Cardoso V,et al. Flower abscission in Vitis vinifera L. triggered by gibberellic acid and shade discloses differences in the underlying metabolic pathways[J]. Frontiers in Plant Science,2015,6:457.
[16]Gao Y R,Liu C,Li X D,et al. Transcriptome profiling of petal abscission zone and functional analysis of an Aux/IAA family gene RhIAA16 involved in petal shedding in rose[J]. Frontiers in Plant Science,2016,7(R106):1375.
[17]王高奇. 乙烯依赖和非乙烯依赖的花器官脱落途径调控AtDOF4.7的研究[D]. 北京:中国农业大学,2015:4-6.
[18]Xie R J,Ge T,Zhang J,et al. The molecular events of IAA inhibiting citrus fruitlet abscission revealed by digital gene expression profiling[J]. Plant Physiology and Biochemistry,2018,130:192-204.
[19]Liu J P,Zhang Z Q,Li H M,et al. Alleviation of effects of exogenous ethylene on cut ‘Master carnation flowers with nano silver and silver thiosulfate[J]. Postharvest Biology and Technology,2018,143:86-91.
[20]Jones M L,Woodson W R. Differential expression of three members of the 1-aminocyclopropane-1-carboxylate synthase gene family in carnation[J]. Plant Physiology,1999,119(2):755-764.
[21]Wang H,Chang X X,Lin J,et al. Transcriptome profiling reveals regulatory mechanisms underlying corolla senescence in petunia[J]. Horticulture Research,2018,5(1):16.
[22]王艳玲. MicroRNA1917调控番茄花柄脱落的分子机制研究[D]. 沈阳:沈阳农业大学,2017:7-10.
[23]Shi Z H,Jiang Y,Han X Q,et al. SlPIN1 regulates auxin efflux to affect flower abscission process[J]. Scientific Reports,2017,7:14919.
[24]Hagemann M H,Winterhagen P,Hegele M,et al. Ethephon induced abscission in mango:physiological fruitlet responses[J]. Frontiers in Plant Science,2015,6:706.
[25]Li C Q,Wang Y,Ying P Y,et al. Genome wide digital transcript analysis of putative fruitlet abscission related genes regulated by ethephon in litchi[J]. Frontiers in Plant Science,2015,6(12):502.
[26]Hauser F,Li Z,Waadt R,et al. Snapshot:abscisic acid signaling[J]. Cell,2017,171(7):1708-1708.
[27]張全军,钟必凤,李文贵,等. ‘丰水梨二次开花过程中枝芽多胺含量的变化[J]. 西南农业学报,2016,29(9):2221-2224.
[28]Roberts J A,Elliott K A,Gonzalezcarranza Z H. Abscission,dehiscence,and other cell separation processes[J]. Annual Review of Plant Biology,2002,53(1):131-158.
[29]Qi X X,Hu S,Zhou H S,et al. A MADS box transcription factor of ‘Kuerlexiangli(Pyrus sinkiangensis Yu) PsJOINTLESS gene functions in floral organ abscission[J]. Gene,2018,642:163-171.
[30]Yang Z Q,Zhong X M,Fan Y,et al. Burst of reactive oxygen species in pedicel mediated fruit abscission after carbohydrate supply was cut off in longan (Dimocarpus longan)[J]. Frontiers in Plant Science,2015,6:360.
[31]Goldental C S,Burstein C,Biton I,et al. Ethephon induced oxidative stress in the olive leaf abscission zone enables development of a selective abscission compound[J]. BMC Plant Biology,2017,17(1):87.
[32]Chersicola M,Kladnik A,Tusek Z M,et al. 1-aminocyclopropane-1-carboxylate oxidase induction in tomato flower pedicel phloem and abscission related processes are differentially sensitive to ethylene[J]. Frontiers in Plant Science,2017,8:464.
[33]Srivignesh S,Sonia P H,Joseph R,et al. De novo transcriptome sequencing and development of abscission zone-specific microarray as a new molecular tool for analysis of tomato organ abscission[J]. Frontiers in Plant Science,2016,6:1258.
[34]Tabuchi T,Arai N. Changes in esterase activity in the abscission zone of jointless tomato fruits[J]. Journal of the Japanese Society for Horticultural Science,1999,68(6):1152-1154.
[35]Ratner A,Goren R,Monselise S P. Activity of pectin esterase and cellulase in the abscission zone of citrus leaf explants[J]. Plant Physiology,1969,44(12):1717-1723.
[36]楊子琴. 饥饿胁迫诱导龙眼果实脱落的信号发生与调控机理研究[D]. 广州:华南农业大学,2011:1-18.
[37]齐明芳,李天来,许 涛,等. 园艺作物器官脱落相关酶的研究进展[J]. 北方园艺,2007(6):62-65.
[38]Patharkar O R,Walker J C. Advances in abscission signaling[J]. Journal of Experimental Botany,2017,69(4):733-740.
[39]Bar-Dror,Dermastia M,Kladnik A,et al. Programmed cell deathoccurs asymmetrically during abscission in tomato[J]. Plant Cell,2011,23(11):4146-4163.
[40]Ma N,Ma C,Liu Y,et al. Petal senescence:a hormone view[J]. Journal of Experimental Botany,2018,69(4):719-732.
[41]丁坤峰,谭晓荣. 植物NADPH氧化酶研究进展[J]. 生命科学,2010(8):723-728.
[42]Garciatejero I,Romerovicente R,Jimenezbocanegra J A,et al. Response of citrus trees to deficit irrigation during different phenological periods in relation to yield,fruit quality,and water productivity[J]. Agricultural Water Management,2010,97(5):689-699.
[43]伏健民,束怀瑞. 春季干旱对金冠苹果不同部位叶片衰老和脱落的影响[J]. 果树科学,1993(2):65-68.
[44]Rodrigo J,Herrero M. Effects of pre-blossom temperatures on flower development and fruit set in apricot[J]. Scientia Horticulturae,2002,92(2):125-135.
[45]Higuchi H,Utsunomiya N,Sakuratani T. High temperature effects on cherimoya fruit set,growth and development under greenhouse conditions[J]. Scientia Horticulturae,1998,77(1/2):23-31.
[46]Tromp J,Borsboom O. The effect of autumn and spring temperature on fruit set and on the effective pollination period in apple and pear[J]. Scientia Horticulturae,1994,60(1/2):23-30.
[47]Ebadi A,Coombe B G,May P. Fruit-set on small Chardonnay and Shiraz vines grown under varying temperature regimes between budburst and flowering[J]. Australian Journal of Grape and Wine Research,1995,1(1):3-10.
[48]Ferree D C,Mcartney S J,Scurlock D M. Influence of irradiance and period of exposure on fruit set of French-American hybrid grapes[J]. Journal of the American Society for Horticultural Science,2001,126(3):283-290.
[49]Zhu H,Dardick C D,Beers E P,et al. Transcriptomics of shading induced and NAA induced abscission in apple(Malus domestica)reveals a shared pathway involving reduced photosynthesis,alterations in carbohydrate transport and signaling and hormone crosstalk[J]. BMC Plant Biology,2011,11(1):138.
[50]Einhorn T C A M,Shading I. Pear abscission and inhibit photosynthesis but are not additive[J]. Journal of Plant Growth Regulation,2018,37(1):300-308.
[51]刘 坤. “汉水砂梨”早期落叶的诱因调查及初步分析[J]. 中国果树,2016(2):91-93,103.
[52]李忠才,黄新忠,林洪龙,等. 梨早期落叶原因与对策[J]. 河北果树,2006(5):44-45.
[53]Duchêne E,Jaegli N,Salber R,et al. Effects of ripening conditions on the following seasons growth and yield components for Pinot noir and Gewurztraminer grapevines(Vitis vinifera L.)in a controlled environment[J]. OENO One,2003,37(1):39-49.
[55]Iwanami H,Moriyat Y,Honda C,et al. Relationships among apple fruit abscission,source strength,and cultivar[J]. Scientia Horticulturae,2012,146,39-44.
[55]Sawicki M,Ait B E,Clement C,et al. Cross talk between environmental stresses and plant metabolism during reproductive organ abscission[J]. Journal of Experimental Botany,2015,66(7):1707-1719.
[56]Zhu H,Yuan R,Greene D W,et al. Effects of 1-methylcyclopropene and naphthaleneacetic acid on fruit set and expression of genes related to ethylene biosynthesis and perception and cell wall degradation in apple[J]. Journal of the American Society for Horticultural Science,2010,135(5):402-409.
[57]Estornell L H,AgustaJ,Merelo P,et al. Elucidating mechanisms underlying organ abscission[J]. Plant Science,2013,199(3):48-60.
[58]Deng Y,Wu Y,Li Y F. Effects of high CO2 and low O2 atmospheres on the berry drop of ‘Kyoho grapes[J]. Food Chemistry,2007,99(2):768-773.
[59]Mao L,Begum D,Chuang H W,et al. JOINTLESS is a MADS box gene controlling tomato flower abscission zone development[J]. Nature,2000,406(6798):910-913.
[60]Guan X,Xu T,Gao S,et al. Temporal and spatial distribution of auxin response factor genes during tomato flower abscission[J]. Journal of Plant Growth Regulation,2014,33(2):317-327.
[61]Ito Y,Nakano T. Development and regulation of pedicel abscission in tomato[J]. Frontiers in Plant Science,2015,6:442.
[62]Roldan M,Perilleux C,Morin H,et al. Natural and induced loss of function mutations in SlMBP21 MADS box gene led to jointless2 phenotype in tomato[J]. Scientific Reports,2017,7(1):4402.
[63]王 翔. 番茄花柄離区发育的分子生物学研究[D]. 北京:中国农业科学院,2008:5-7.
[64]齐笑笑. 梨果实萼片宿存与脱落过程基因表达谱分析及PsIDA、PsJOINTLESS基因功能的初步研究[D]. 南京:南京農业大学,2014:7-18.
[65]Lee Y,Yoon T H,Lee J,et al. A lignin molecular brace controls precision processing of cell walls critical for surface integrity in Arabidopsis[J]. Cell,2018,173(6):1468-1480.
[66]Ying P,Li C,Li X,et al. Identification and molecular characterization of an IDA-like gene from litchi,LcIDL1,whose ectopic expression promotes floral organ abscission in Arabidopsis[J]. Scientific Reports,2016,6:37135.
[67]St I M,Orr R J,Fooyontphanich K,et al. Conservation of the abscission signaling peptide IDA during angiosperm evolution:withstanding genome duplications and gain and loss of the receptors HAE/HSL2[J]. Frontiers in Plant Science,2015,6(634):931.
[68]韩亚萍. 桃体细胞胚发生相关基因SERK2的克隆与表达分析[D]. 郑州:河南农业大学,2017:4-7.
[69]Testone G,Bruno L,Condello E,et al. Peach[Prunus persica(L.)Batsch]KNOPE1,a class 1 KNOX orthologue to Arabidopsis BREVIPEDICELLUS/KNAT1,is misexpressed during hyperplasia of leaf curl disease[J]. Journal of Experimental Botany,2008,59(2):389-402.
[70]Yu Q,Steiger D,Kramer E M,et al. Floral mads box genes in trioecious papaya:characterization of AG and AP1 subfamily genes revealed a sex type specific gene[J]. Tropical Plant Biology,2008,1(2):97-107.
[71]褚婷婷,谢 华,徐 勇,等. 植物MADS-box基因FRUITFULL(FUL)研究进展[J]. 中国生物工程杂志,2010,30(9):98-104.
[72]杜 艺. 荔枝落果过程中IAA和ABA相关基因表达及其定量检测初探[D]. 广州:华南农业大学,2016:1-10.
[73]Aalen R B,Wildhagen M,St I M,et al. Ida:a peptide ligand regulating cell separation processes in Arabidopsis[J]. Journal of Experimental Botany,2013,64(17):5253-5261.
[74]Zhang Q,Zhang H,Sun L,et al. The genetic architecture of floral traits in the woody plant Prunus mume[J]. Nature Communications,2018,9(1):1702.
[75]关晓溪. 番茄花柄脱落相关生长素响应基因ARFs的筛选及其功能的初步鉴定[D]. 沈阳:沈阳农业大学,2015:1-18.
[76]Liu K,Feng S,Jiang Y,et al. Identification and expression analysis of seven MADS box genes from Annona squamosa[J]. Biologia Plantarum,2017,61(1):24-34.
[77]Jibran R,Tahir J,Cooney J,et al. Arabidopsis AGAMOUS regulates sepal senescence by driving jasmonate production[J]. Frontiers in Plant Science,2017,8:2101.
[78]Zhang Y,Zeng Z,Chen C,et al. Genome-wide characterization of the auxin response factor (ARF) gene family of litchi (Litchi chinensis Sonn.):phylogenetic analysis,miRNA regulation and expression changes during fruit abscission[J]. Peer J,2019,7:e6677.
[79]李慧峰,冉 昆,何 平,等. 苹果生长素响应因子(ARF)基因家族全基因组鉴定及表达分析[J]. 植物生理学报,2015,51(7):1045-1054.
[80]Xie R,Pang S,Ma Y,et al. The ARF,AUX/IAA and GH3 gene families in citrus:genome-wide identification and expression analysis during fruitlet drop from abscission zone A[J]. Molecular Genetics and Genomics,2015,290(6):2089-2105.
[81]常 璟. 棉花離层相关基因GhBOP1的分离及功能鉴定[D]. 兰州:甘肃农业大学,2015:6-7.
[82]叶孋婵. 番石榴果实对乙烯敏感性与其受体cDNA选殖表现分析[D]. 台湾:台湾大学,2013.
[83]王 翔,陈晓博,李爱丽,等. 植物器官脱落分子生物学研究进展[J]. 作物学报,2009,35(3):381-387.
[84]Wu J Y,Zhang H N,Liu L,et al. Validation of reference genes for RT qPCR studies of gene expression in preharvest and postharvest longan fruits under different experimental conditions[J]. Frontiers in Plant Science,2016,7(439):780.
[85]Burko Y,Shleizer B S,Yanai O,et al. A role for APETALA1/FRUITFULL transcription factors in tomato leaf development[J]. Plant Cell,2013,25(6):2070-2083.
[86]Mouhu K,Hytnen T,Folta K,et al. Identification of flowering genes in strawberry,a perennial SD plant[J]. BMC Plant Biology,2009,9(1):122.
[87]Wang J,Zhang X M,Yan G H,et al. Over expression of the PaAP1,gene from sweet cherry(Prunus avium L.)causes early flowering in Arabidopsis thaliana[J]. Journal of Plant Physiology,2013,170(3):315-320.
[88]Chi Y J,Wang T T,Xu G L,et al. GmAGL1,a MADS box gene from soybean,is involved in floral organ identity and fruit dehiscence[J]. Frontiers in Plant Science,2017,8(242):175.
[89]Pathak A K,Singh S P,Gupta Y,et al. Transcriptional changes during ovule development in two genotypes of litchi (Litchi chinensis Sonn.) with contrast in seed size[J]. Scientific Reports,2016,6:36304.
[90]廖文彬,杨义伶,李雅韵,等. 植物离区发育及调控信号的多样性研究进展[J]. 热带农业科学,2014,34(12):42-48.
[91]陈晓博. 参与番茄花柄离区发育的转录因子SPL3的基因功能研究[D]. 北京:中国农业科学院,2010:1-13.
[92]Schumacher K,Schmitt T,Rossberg M,et al. The Lateral suppressor(Ls) gene of tomato encodes a new member of the VHIID protein family[J]. Proceedings of the National Academy of Sciences of the United States of America,1999,96(1):290-295.
[93]Sundaresan S,Philosoph H S,Ma C,et al. The tomato hybrid proline rich protein regulates the abscission zone competence to respond to ethylene signals[J]. Horticulture Research,2018,5(1):28.
[94]Lanahan M B,Yen H C,Giovannoni J J,et al. The never ripe mutation blocks ethylene perception in tomato[J]. Plant Cell,1994,6(4):521-530.
[95]Klee H J. Control of ethylene mediated processes in tomato at the level of receptors[J]. Journal of Experimental Botany,2002,53(377):2057-2063.
[96]王 鹏,王丹丹. 乙烯利、脱落酸对采收前艳红桃成熟期的调控[J]. 江苏农业科学,2018,46(23):110-114.
[97]王男麒. 柑橘花果脱落规律及其矿质养分损耗[D]. 重庆:西南大学,2014:20-45.
[98]Zhu C,Zheng X,Huang Y,et al. Genome sequencing and CRISPR/Cas9 gene editing of an early flowering mini citrus(Fortunella hindsii)[J]. Plant Biotechnology Journal,2019,17(11):2199-2210.
[99]Charrier A,Vergne E,Dousset N,et al. Efficient targeted mutagenesis in apple and first time edition of pear using the CRISPR-Cas9 system[J]. Frontiers in Plant Science,2019,10:40.
[100]Osakabe Y,Liang Z,Ren C,et al. CRISPR-Cas9-mediated genome editing in apple and grapevine[J]. Nature Protocols,2018,13(12):2844-2863.
[101]Zhou J H,Li D D,Wang G M,et al. Application and future perspective of CRISPR/Cas9 genome editing in fruit crops[J]. Journal of Integrative Plant Biology,2019,62(3):269-286.