三种流速对卵形鲳鲹幼鱼耗氧率和排氨率的影响
2020-08-24苏家齐秦金华朱长波张博陈素文李俊伟
苏家齐 秦金华 2 朱长波 *张博 陈素文 李俊伟
三种流速对卵形鲳鲹幼鱼耗氧率和排氨率的影响
苏家齐1, 秦金华1, 2, 朱长波1, *,张博1, 陈素文1, 李俊伟1
1. 中国水产科学研究院南海水产研究所, 农业农村部南海渔业资源开发利用重点实验室, 广东省渔业生态环境重点实验室, 广州 510300 2. 上海海洋大学海洋科学学院, 上海 201306
采用实验生态学方法, 研究了饥饿态下平均体重分别为(2.54±1.09) g、(24.35±7.19) g、(233.79±26.00) g、(508.98±126.61) g的卵形鲳鲹幼鱼()在静水、0.1 m·s-1、0.2 m·s-1三种流速下的耗氧率和排氨率的变化。结果表明, 卵形鲳鲹体重对耗氧率和排氨率影响显著(< 0.05)。在同一流速下, 耗氧率和排氨率分别随体重的增加而下降, 体重与两者间的回归关系符合异速方程Y=aWb。卵形鲳鲹的耗氧率和排氨率随着流速的升高均出现升高的趋势, 流速对均重为24.35 g和233.79 g卵形鲳鲹的耗氧率和排氨率影响显著(< 0.05)。在静水条件下, 卵形鲳鲹代谢氧氮比的范围为31.36—46.32, 表明卵形鲳鲹主要以脂肪和碳水化合物为能源, 蛋白质其次。在试验流速范围内, 均重为24.35 g卵形鲳鲹幼鱼的氧氮比随着流速的升高而升高, 其他两组的氧氮比则随着流速的升高而降低。
卵形鲳鲹; 耗氧率; 排氨率; 流速
0 前言
卵形鲳鲹()又称金鲳、黄腊鲳, 为暖水性中上层洄游鱼类, 广泛分布于我国南海、日本沿海、地中海、印度洋等热带亚热带海域。卵形鲳鲹肉质细嫩、味道鲜美, 生长速度快, 养殖周期短, 且对环境具有较强的适应能力, 备受消费者喜爱, 是我国华南重要的海水养殖鱼类品种。在我国广东、海南已进行大规模的人工养殖, 特别是网箱深海养殖[1]。
呼吸和排泄是鱼类进行新陈代谢的基本生理活动, 它不仅可以反映鱼类的生理状态, 也通过呼吸耗氧及排泄氨氮影响周围的环境。水流可以增加鱼类代谢系统的负担, 是影响鱼类行为、生长、繁殖等重要环境因子之一[2]。在网箱养殖过程中, 箱内水体的流速与鱼类生长密切相关。流速大小决定了网箱内外水体的交换速度, 对箱内水质环境影响显著[3]。在大西洋鲑的网箱养殖中, 流速、网箱性状、鱼类的呼吸作用及行为共同影响了网箱内部的溶氧变化及分布情况, 间接限制了大西洋鲑的生长[4]。耗氧率和排氨率是反映水生生物代谢状态的重要指示因子, 还能为水生生物的能量收支模型的构建以及校正提供必要的参数[5–6]。近年来, 学者们对卵形鲳鲹的耗氧率和排氨率进行了一系列的研究报道, 这些报道主要集中于温度、盐度、光照、PH等环境因子对卵形鲳鲹仔、稚鱼的呼吸代谢的影响[7–9]。本研究运用封闭式流水实验方法以及闭合式呼吸运动测量法, 研究流速和体重对卵形鲳鲹幼鱼耗氧率和排氨率的影响, 为卵形鲳鲹的代谢研究提供基础资料, 为其养殖生产、运输、养殖水体水质调控等提供数据支持及科学依据
1 材料与方法
1.1 实验材料
实验用鱼为深圳市龙岐庄实业发展有限公司培育, 挑选健康、活力强的四种规格的卵形鲳鲹幼鱼于中国水产科学研究院南海水产研究所深圳试验基地用圆形水桶(高0.9 m, 直径1 m)暂养7 d, 实验用水为沙滤海水, 暂养及实验水温为(30.86±0.58) °C, 盐度为31.65±0.98。四种规格卵形鲳鲹的体质量由小到大分别为(2.54±1.09) g、(24.35±7.19) g、(233.79± 26.00) g以及(508.98±126.61) g。体长分别为(4.34± 0.61) cm、(9.32±0.97) cm、(20.94±1.41) cm以及(26.12±2.07) cm。实验用鱼于实验开始前24 h停止投饵。
1.2 实验方法及实验设计
采用沙滤海水, 设置静水、0.1 m·s-1、0.2 m·s-1三个流速实验组。采用封闭式流水实验方法测定静水条件下卵形鲳鲹耗氧率与排氨率。采用闭合式呼吸运动测量法测量流速为0.1 m·s-1、0.2 m·s-1条件下卵形鲳鲹耗氧率与排氨率。每组实验设置4个平行组及1个空白对照组。其中, 四种规格由小到大卵形鲳鲹在平行组每个呼吸室中放置的数量分别为30尾、6尾、1尾以及1尾。空白对照组与平行组实验装置相同, 但呼吸室内不放置卵形鲳鲹。实验装置的呼吸室大小为0.2×0.25×0.5 m。
在封闭流水式实验中, 实验鱼放入呼吸室后, 密封呼吸室并排尽室内空气, 海水由贮水箱经进、出水口流入及流出呼吸室。实验期间呼吸室流速保持稳定, 四次实验流量分别为61.02、87.07、44.24以及66.44 L·h-1。于实验开始后2 h和3 h分别在进出水口取样, 测定进出水口的水温、溶氧量和氨氮浓度。
在闭合式呼吸运动测量法测量实验中, 使用的鱼类游泳运动装置实验装置如图1所示。装置总容量为121 L, 密封后该区域内的溶解氧不与外界交换。鱼类游泳区规格同呼吸室, 装置通过变频水泵控制游泳区水流的大小。在装置密封前和鱼类游泳30 min后分别取样, 测量装置内水样的水温、溶氧量和氨氮浓度。
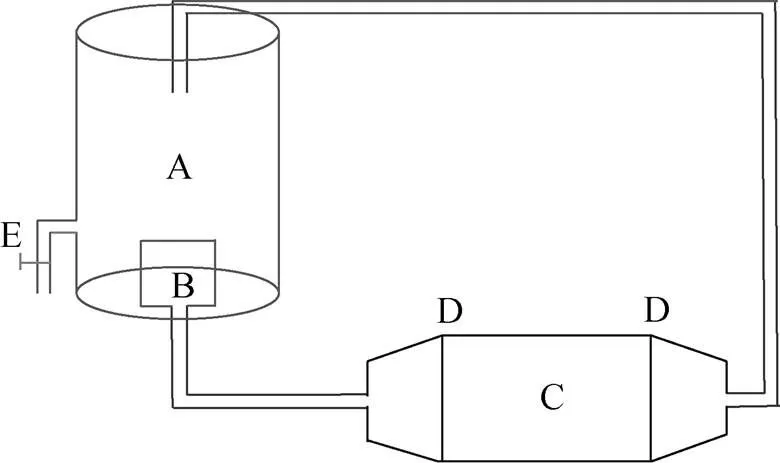
A: 储水槽; B: 变频水泵; C: 鱼类游泳区; D: 导流筛板 E取样口。
Figure 1 Schematic diagram of experimental device
1.3 测定方法及数据处理
采用次溴酸钠氧化法测定水中的氨氮, 溶解氧采用碘量法测定。每组实验结束后, 用毛巾吸干鱼体表的水分, 使用精度为0.01 g的电子天平称量鱼的体质量。封闭流水式实验下耗氧率(R)和排氨率(R)的计算公式如下:
R=(C- CDO20-C+C) ×/
R=(C- C- C+C) ×/
闭合式呼吸运动测量法测量实验下耗氧率和排氨率的计算公式如下:
R=(k–k) ××/
R=(K–K) ××/
氧氮比: O:N=R×14/R×16。
上述各式中,C、C分别为流入实验组以及对照组呼吸室的海水溶氧量,C、C分别为流出实验组以及对照组呼吸室的海水溶氧浓度(mg·L-1)。Q为单位时间水流量(L·h-1), W实验鱼体质量(kg)。C、C分别为流入实验组以及对照组呼吸室的海水氨氮浓度,C、C分别为流出实验组以及对照组呼吸室的海水氨氮浓度(mg·L-1)。k、k分别为测定期间实验组及对照组水中溶解氧曲线斜率[mg·(L·min)-1],K、K分别为测定期间实验组及对照组水中氨氮曲线斜率[mg·(L·min)-1]。为装置总容量(L)。
实验结果数据使用SPSS19.0以及Excel进行分析, 运用单因素方差分析进行显著性检验。不满足方差齐性时, 对数据进行自然对数或平方根转换。
2 结果与分析
2.1 体重和流速对卵形鲳鲹耗氧率的影响
三种流速下的卵形鲳鲹耗氧率与体重之间的关系及回归方程如表1所示, 在相同流速下, 卵形鲳鲹的耗氧率随着体质量的增加而减少。体质量2.54 g组卵形鲳鲹幼鱼在静水条件下耗氧率最高并和其他体质量组差异极显著(< 0.01), 且该规格的幼鱼无法在0.1 m·s-1及以上的水流中长期游泳。在同一流速下, 体质量24.35 g组卵形鲳鲹的耗氧率与其他组别差异极显著(< 0.01)。而233.7 g组与508.98 g组两组之间耗氧率差异不显著(> 0.05)。在静水、0.1 m·s-1、0.2 m·s-1流速条件下卵形鲳鲹耗氧率与体质量的回归方程分别为y=1547.5x-0.267、y=1561.9x-0.193和y =8418.1x-0.437。同时, 卵形鲳鲹耗氧率随着水流速度的加快而增加。其中, 均重为24.35 g组和均重为233.7 g组的卵形鲳鲹在三种流速下的耗氧率差异极显著(P < 0.01)。均重为508.98 g组在静水条件下的耗氧率与水流速度为0.1 m·s-1条件下的耗氧率差异显著(< 0.05)与0.2 m·s-1条件下的耗氧率差异极显著(< 0.01)。508.98 g组卵形鲳鲹在另外两组水流下的耗氧率差异不显著(> 0.05)(图2)。
2.2 体重和流速对卵形鲳鲹排氨率的影响
三种流速下的卵形鲳鲹排氨率与体重之间的关系及回归方程如表2所示, 在相同流速下, 卵形鲳鲹的排氨率随着体质量的增加而减少。在静水条件下, 均重为2.54 g组卵形鲳鲹幼鱼的排氨率高于其他三组且差异极显著(< 0.01)。均重为24.35 g组的卵形鲳鲹的排氨率高于233.7 g组但无显著差异(> 0.05), 同时高于508.98 g组且差异显著(< 0.05)。233.7 g组与508.98 g组两组之间差异不显著(> 0.05)。在0.1 m·s-1的水流速度下, 卵形鲳鲹各体质量组间差异极显著(< 0.01)。在0.2 m·s-1的水流速度下, 均重为24.35 g组卵形鲳鲹的排氨率与其他两组差异极显著(< 0.01), 233.7 g组与508.98 g组间的排氨率差异显著(< 0.05)。在静水条件、0.1 m·s-1、0.2 m·s-1流速条件下卵形鲳鲹排氨率与体质量的回归方程分别为y=27.81x-0.213、y=56.847x-0.271和y=456.04x-0.607。同时, 卵形鲳鲹排氨率随着水流速度的加快而增加。其中, 均重为24.35 g组在三种流速下的排氨率差异极显著(< 0.01)。均重为233.7 g组的卵形鲳鲹在静水条件下的排氨率低于其他两种流速下的排氨率且差异极显著(< 0.01), 0.1 m·s-1、0.2 m·s-1流速下的排氨率差异显著(< 0.05)。均重为508.98 g组在静水条件下的排氨率与其他两种流速下的排氨率差异显著(< 0.05), 其他两种流速下的排氨率差异不显著(> 0.05)。(图3)。

表1 不同流速下卵形鲳鲹体重与耗氧率的关系及回归方程
注: 上标无相同字母者表示两两比较差异显著(< 0.05), 下同

图2 流速对卵形鲳鲹耗氧率的影响
Figure 2 The influence of water velocity on oxygen consumption rate in.
2.3 体重和流速对卵形鲳鲹氧氮比的影响
三种流速下不同质量的卵形鲳鲹的氧氮比如表3所示, 均重为24.35 g组的卵形鲳鲹氧氮比范围为27.84—43.15且随着流速的增加而降低。其他两组的氧氮比则随流速的增加而增加。在静水条件下, 在均重为2.54—233.79 g范围之内卵形鲳鲹的氧氮比随体质量的增加而减小, 从46.32降到31.36。而在均重为508.98 g组中, 氧氮比为37.88, 高于233.79 g组。
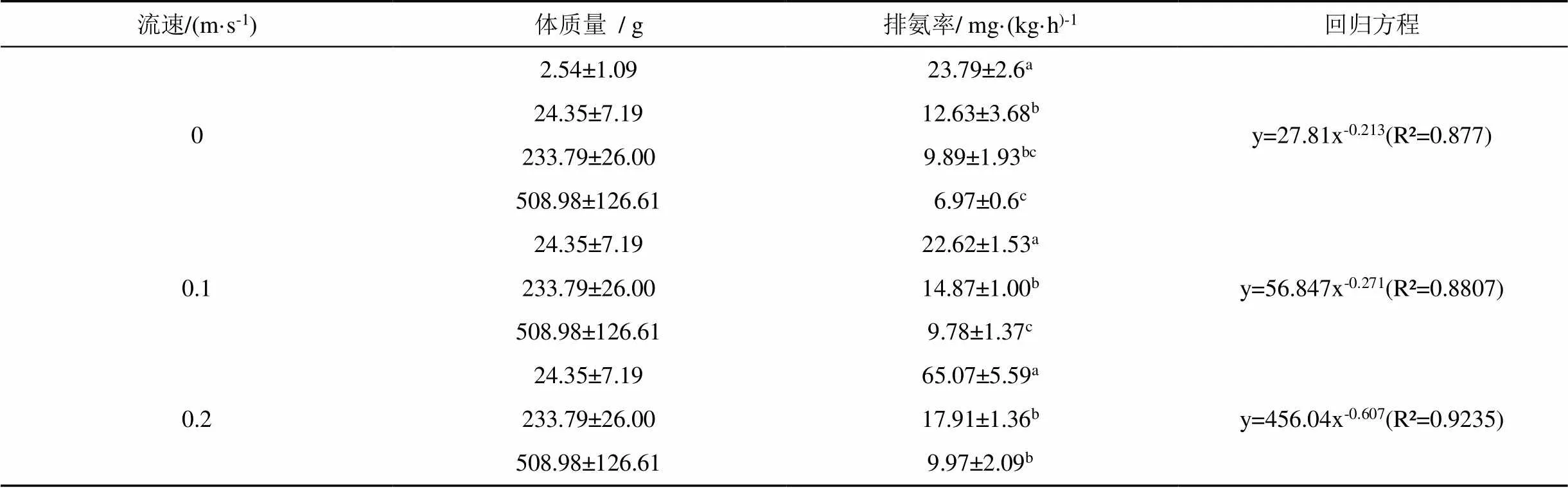
表2 不同流速下卵形鲳鲹体重与排氨率的关系及回归方程
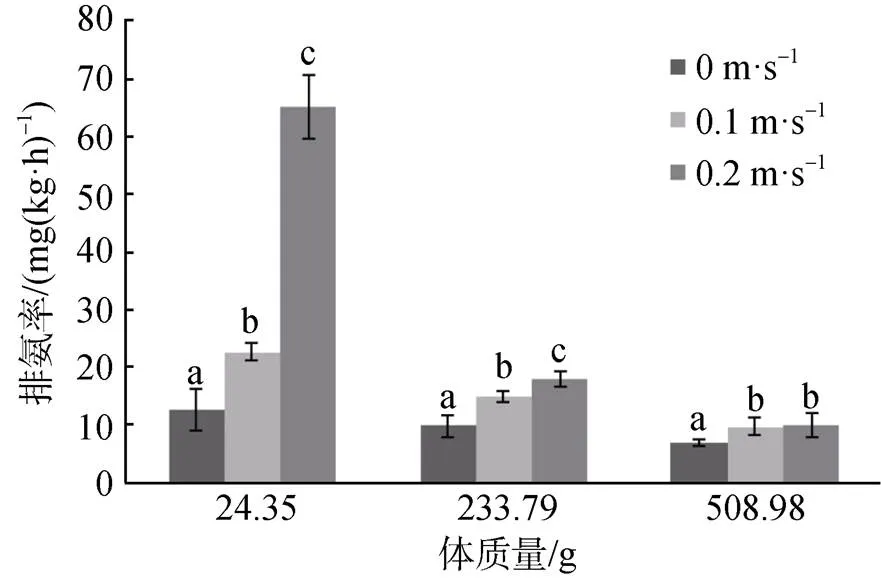
图3 流速对卵形鲳鲹排氨率的影响
Figure 3 The influence of water velocity on ammonia excretion rate in.
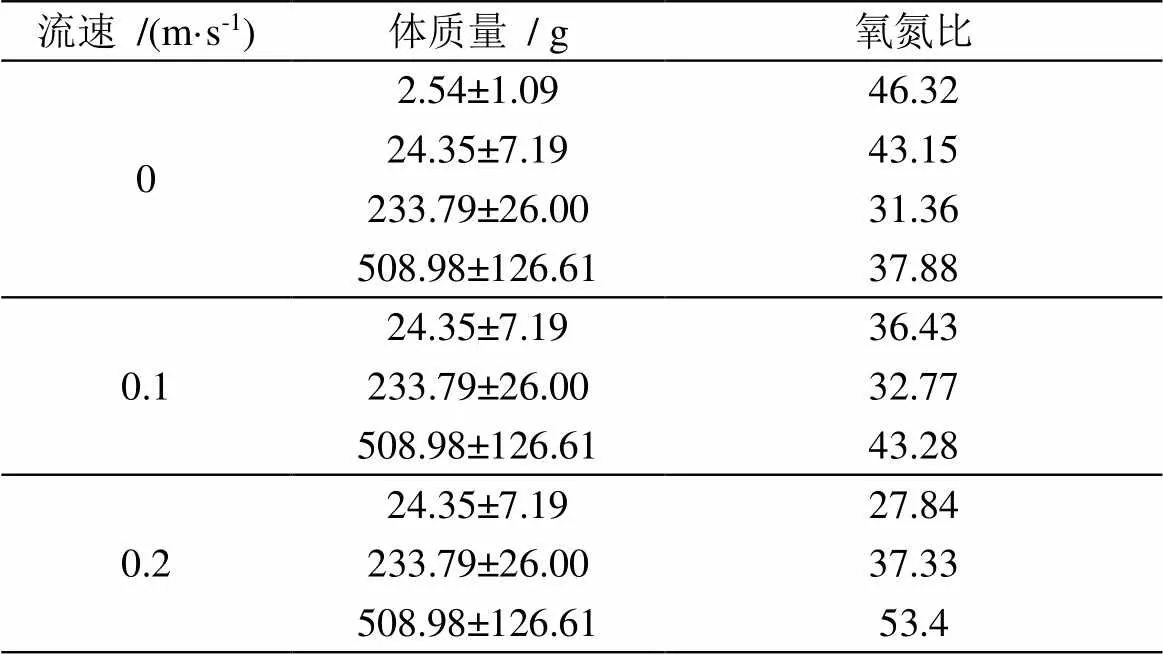
表3 不同条件下卵形鲳鲹的氧氮比
3 讨论
3.1 体重对卵形鲳鲹耗氧率和排氨率的影响
鱼类的呼吸代谢不仅受到环境因素的影响, 还与其个体的生理特性及体质量相关。在一定的环境条件下, 鱼类的耗氧率与排氨率通常随着体重的增加而下降。研究表明, 斑石鲷(50.94—194.56 g)的耗氧率在20—30 ℃条件下、排氨率在25和30℃条件下随着体重的增大而降低[10]。在同一温度条件下(15 °C、20 ℃、25 ℃), 体重对红鳍东方鲀()幼鱼(23.9—43.8 g)的耗氧率和排氨率呈负相关且影响显著[11]。在自然光和遮光条件下, 卵形鲳鲹幼鱼的耗氧率与排氨率与鱼体重(0.30—2.81 g, 0.30—1.49 g)呈负相关[7]。在本研究中, 同一流速条件下的卵形鲳鲹随着规格的增加, 其耗氧率与排氨率逐渐降低, 结果与前述报道较为接近。学者们认为鱼类在小个体时期时生长发育相对较快, 单位体重的代谢水平较高, 因此体重较小的幼鱼耗氧率和排氨率较高。随着鱼类的生长, 直接维持生命的肝、肾等器官的脏体比逐渐降低, 导致成体的耗氧率和排氨率和幼体相比均有所降低[12–13]。
3.2 流速对卵形鲳鲹耗氧率和排氨率的影响
鱼类对水流变化造成的刺激会产生相应的行为上的反应, 随着流速的增加, 鱼类会产生趋流行为, 且随着游泳速度的加快, 其耗氧率也相应增加[14]。对白斑红点鲑()、鳊鱼()幼鱼和宽鳍鱲()幼鱼等鱼类的呼吸代谢研究结果表明鱼类在流水条件下的耗氧率均高于静水, 且耗氧率随着流速的提高而上升[15–16]。本研究的结果与上述研究结果一致。
关于流速对鱼类排氨率的影响, 国内外在此方面的报道不多。Alsop等研究了尼罗罗非鱼()在不同流速下的能量代谢, 结果表明在流速为15—45 cm·s-1的条件下, 罗非鱼的排氨率与随着流速的加快而上升, 在55 cm·s-1的水流下排氨率低于其他各组[17]。王刚等100—250 mL·min-1流量范围内测定卵形鲳鲹幼鱼的呼吸代谢, 结果显示卵形鲳鲹幼鱼的排氨率随着流速的增大先增大后减小[7]。本研究结果表明, 在0—0.2 m·s-1低流速下, 体质量为24.35—508.98 g的卵形鲳鲹排氨率随流速的增大而上升。李加儿等在封闭流水式实验下通过调节进水口流速(0.15 m·s-1、0.30 m·s-1、0.45 m·s-1、0.60 m·s-1、0.75 m·s-1。)研究了水流对鲻()幼鱼的呼吸代谢的影响。结果表明鲻幼鱼在0.30 m·s-1时的耗氧率和排氨率最低, 之后排氨率随着水交换速度上升而增高[18]。Skov等对三种流速条件下长期饲养虹鳟鱼()的能量代谢开展研究, 结果表明虹鳟的耗氧率随水流升高而增大, 排氨率则在0.5 BL·s-1流速环境下低于静水条件下, 在1 BL·s-1下的排氨率则高于其他两组[19]。李丹发现体长为21.08 cm的杂交鲟在静水组和0.1 m·s-1流速组的趋流率、摆尾频率在大部分时段无显著性差异[19]。上述结果表明鱼类的排氨率可能与鱼体的逆流游泳运动行为正相关, 水流通过对鱼体造成的逆流游泳运动增加鱼类的排氨率。鱼类在感应流速以下的逆流游泳运动行为不明显, 其排氨率与水流之间的关系也存在一定的波动。
3.3 体重和流速对卵形鲳鲹代谢氧氮比的影响
氧氮比(O/N)常被作为估计生物体呼吸和排泄过程中供能物质的指标, 氧氮比值能够随着鱼体中脂类和碳水化合物被利用比例逐渐增加而逐渐升高, 进而能够反映出为生物体提供能量的蛋白质、脂肪与碳水化合物的比值关系[12, 20]。在本实验条件下, 随着流速的上升, 均重为24.35 g的卵形鲳鲹幼鱼的氧氮比逐渐下降, 表明其供能物质中蛋白质所占的比例逐渐上升。同时体质量较大的两组氧氮比呈上升趋势。糖原是糖类在鱼体内的储存形式, 葡萄糖氧化分解是供给鱼类能量的重要途径, 血糖(葡萄糖)则是糖类在体内的主要运输形式[22]。晁帅等报道美国红鱼()在游泳过程中, 血糖浓度会显著提高, 肌糖原少量减少, 肝糖原显著降低, 鱼类游泳接近疲劳时, 肝糖原浓度亦接近耗尽[23]。在本实验中, 鱼类在游泳运动中对碳水化合物的利用程度增加, 进而引起氧氮比的上升, 而随着糖类的消耗, 导致代谢底物成分的变化, 可能会引起氧氮比的下降。在同等流速下, 体型较小的卵形鲳鲹更易疲劳, 其对碳水化合物的消耗较快, 进而导致其氧氮比随着流速的上升而逐渐下降。
在静水条件下, 卵形鲳鲹的氧氮比值则随体重的增大呈现先降低后上升的趋势。该结果与大弹涂鱼)的基本代谢规律一致[24]。总体上, 卵形鲳鲹在(30.86±0.58) ℃的条件下主要依靠脂肪和碳水化合物供能, 其次为蛋白质。不同的外界环境条件及内在生理生态特性都会影响到鱼类的氧氮比, 如温度、盐度、鱼类不同发育时期等。仇登高等研究了5种水温对3种规格的珍珠龙胆石斑鱼()呼吸和排泄的影响, 结果表明在(15.8±0.1)℃、(21.0±0.1)℃和(36.5±0.1)℃下, 珍珠龙胆石斑鱼的氧氮比值随个体的增大而减少, 而在(26.6±0.1)℃、(30.9±0.1)℃下, 氧氮比值则随个体的增大呈现先降低后上升的趋势[25]。卵形鲳鲹当前氧氮比是否还与组织内部贮存物质的构成和温度等环境因子存在关联, 还需进一步的研究。
[1] 林川, 何永姑, 王小兵. 卵形鲳鲹深海网箱养殖渔获模式的研究[J]. 热带生物学报, 2018, 9(04): 4–10.
[2] 袁喜, 涂志英, 韩京成, 等. 流速对细鳞裂腹鱼游泳行为及能量消耗影响的研究[J]. 水生生物学报, 2012, 36(2): 270–275.
[3] 赵云鹏, 李玉成, 董国海. 深水抗风浪网箱水动力学特性研究[J]. 渔业现代化, 2011, 38(2): 10–16.
[4] SOLSTORM D, OLDMAN T, SOLSTORM F, et al. Dissolved oxygen variability in a commercial sea-cage exposes farmed Atlantic salmon to growth limiting conditions[J]. Aquaculture, 2017, 486: 112–119.
[5] NIE, Hongtao, ZUO, Shuqi, LI, Long, et al. Physiological and biochemical responses ofto different thermal and salinity stressors[J]. Journal of Experimental Zoology Part A: Ecological and Integrative Physiology, 2018, 329(1): 15–22.
[6] CASAS S M, LAVAUD R, PEYRE M K L, et al. Quantifying salinity and season effects on eastern oyster clearance and oxygen consumption rates[J]. Marine Biology, 2018, 165(5): 90–103.
[7] 王刚, 李加儿, 区又君, 等. 环境因子对卵形鲳鲹幼鱼耗氧率和排氨率的影响[J]. 动物学杂志, 2011, 46(6): 80– 87.
[8] 王刚, 李加儿, 区又君, 等. 卵形鲳鲹幼鱼耗氧率和排氨率的初步研究[J]. 动物学杂志, 2010, 45(3): 116–121.
[9] 李金兰, 陈刚, 张健东, 等. 温度、盐度对卵形鲳鲹呼吸代谢的影响[J]. 广东海洋大学学报, 2014, 34(1): 30–36.
[10] 宋协法, 程亚伟, 邢道超, 等. 温度、体重对斑石鲷耗氧率、排氨率的影响及昼夜节律变化[J]. 中国海洋大学学报:自然科学版, 2017, 47(9): 21–29.
[11] 何亚, 王华, 王伟, 等. 温度对不同体质量红鳍东方鲀幼鱼耗氧率和排氨率的影响[J]. 大连海洋大学学报, 2014, 29(5): 481–485.
[12] 冯雪, 周艳波, 范江涛, 等. 鲻鱼在饥饿状态下的耗氧率和排氮率与其体质量及水温变化关系研究[J]. 南方水产科学, 2018, 14(1): 114–120.
[13] 李加儿, 刘士瑞, 区又君, 等. 花尾胡椒鲷幼鱼的呼吸和排泄代谢[J]. 南方水产, 2009, 5(2): 34–39.
[14] 房敏, 蔡露, 高勇, 等. 温度对鲢幼鱼游泳能力及耗氧率的影响[J]. 水生态学杂志, 2013, 34(3): 49–53.
[15] 吴青怡, 曹振东, 付世建. 鳊鱼和宽鳍鱲幼鱼流速选择与运动能量代谢特征的关联[J]. 生态学报, 2016, 26(13): 4187–4194.
[16] 张永泉, 尹家胜, 徐革峰, 等. 水温和体重对白斑红点鲑临界游泳速度和游动耗氧率的影响[J]. 水生生物学报, 2015, (04): 19–26.
[17] ALSOP D H, KIEFFER J D, WOOD C M . The effects of temperature and swimming speed on instantaneous fuel use and nitrogenous waste excretion of the[J]. Physiological and Biochemical Zoology, 1999, 72(4): 474–483.
[18] 李加儿, 曹守花, 区又君, 等. 鲻幼鱼耗氧率、排氨率和窒息点的研究[J]. 四川动物, 2014, 33(3): 434–439.
[19] SKOV P V, LUND I, PARGANA M A. No evidence for a bioenergetic advantage from forced swimming in rainbow trout under a restrictive feeding regime[J]. Frontiers in Physiology, 2015, 6: 31–39.
[20] 李丹, 林小涛, 朱志明, 等. 不同流速下杂交鲟幼鱼游泳状态与活动代谢研究[J]. 水生生物学报, 2011, 35(4): 578–585.
[21] IKEDA T. Respiration and ammonia excretion of euphausiid crustaceans: synthesis toward a global- bathymetric model[J]. Marine Biology, 2013, 160(2): 251–262.
[22] 蔡春芳, 陈立侨. 鱼类对糖的代谢[J]. 水生生物学报, 2008, 32(4): 592-597.
[23] 晁帅, 王萍, 娄宇栋, 等. 美国红鱼游泳体能分配模型及其在海区选址中的应用[J]. 水产学报, 2017, (03): 75–83.
[24] 周银环, 曹伏君, 罗杰, 等. 不同温度梯度和规格体重对大弹涂鱼()耗氧率(Ro)和排氨率(RN)的影响[J]. 海洋与湖沼, 2017, (2): 392–397.
[25] 仇登高, 黄种持, 郑乐云, 等. 温度对不同规格珍珠龙胆石斑鱼(棕点石斑鱼♀×鞍带石斑鱼♂)呼吸和排泄的影响[J]. 中国水产科学, 2017, 24(2): 295–305.
Effects of the three current velocity on the oxygen consumption and ammonia excretion of
SU Jiaqi1, QIN Jinhua1, 2, ZHU Changbo1,*, ZHANG Bo1, CHEN Suwen1, LI Junwei1
1. Key Lab. of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture and Rural Affairs; South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangdong Provincial Key Lab. of Fishery Ecology and Environment, Guangzhou 510300, China 2. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China
The oxygen consumption and ammonia excretion rates of four weight groups (2.54g ±1.09 g, 24.35g ±7.19 g, 233.79g ±26.00 g, 508.98g ±126.61 g) of starved juvenile golden pompanowere investigated under experimental condition with three different current velocities (0 m·s-1, 0.1 m·s-1, 0.2 m·s-1). The results showed that body weight had significant influence on the oxygen consumption and ammonia excretion rate (<0.05) of the fish. Under the same current velocity, both oxygen consumption and ammonia excretion rate ofdecreased while the body weight increased, and the regressions fitted the allometric equation=abwell. The oxygen consumption and ammonia excretion rate ofincreased along with the rise of the current velocity, and significant effects were observed in the body weight groups 24.35 g and 233.79 g (<0.05). The metabolic O (n): N (n) ratio ofvaried from 31.36 to 46.32 in still water condition. It indicates thatuses fat and saccharide as its major metabolic energy source, and protein is the secondary energy source. Under the condition of this study, the metabolic O:N ratio of body weight group 24.35 g increased with current velocity, while the results of the two bigger body weight groups reversed.
; ammonia excretion rate; oxygen consumption rate; current velocity
10.14108/j.cnki.1008-8873.2020.04.008
苏家齐, 秦金华, 朱长波, 等. 三种流速对卵形鲳鲹幼鱼耗氧率和排氨率的影响[J]. 生态科学, 2020, 39(4): 60–65.
SU Jiaqi, QIN Jinhua, ZHU Changbo, et al. Effects of the three current velocity on the oxygen consumption and ammonia excretion of[J]. Ecological Science, 2020, 39(4): 60–65.
S917
A
1008-8873(2020)04-060-06
2019-11-16;
2019-12-10
世界自然基金WWF项目(10000889-102714-1.1.01.01); 中国水产科学研究院中央级公益性科研院所基本科研业务费项目(2019HY-XKQ01-05);中国水产科学研究院院级基本科研业务费专项 (2016HY-JC0103)
苏家齐(1987—), 男, 助理研究员,博士, 从事水产养殖生态学研究. E-mail: sujiaqi2005@126.com
朱长波(1978—), 男, 研究员, 博士, 从事水产养殖生态学与模式研究.E-mail:changbo@scsfri.ac.cn
