长江上游中华金沙鳅和短身金沙鳅线粒体遗传多样性研究
2020-06-06龙安雨田辉伍汪登强陈大庆周湖海段辛斌
龙安雨,田辉伍,汪登强,陈大庆,周湖海,3,段辛斌
(1.上海海洋大学,水产科学国家级实验教学示范中心,上海 201306; 2.中国水产科学研究院长江水产研究所,农业农村部长江中上游渔业资源环境科学观测实验站,武汉 430223;3.华中农业大学水产学院,武汉 430070)
长江上游指宜昌以上江段,该江段地形地貌复杂、河流水系众多,鱼类区系与物种组成具有多样性、特有性特征,有鱼类200多种,其中特有鱼类100多种[1]。近年来,随着经济和社会的发展,该江段大型水利工程相继建设,导致水文等环境条件发生改变,鱼类适宜生境不断破碎、萎缩,种群数量急剧下降[1],鱼类资源和遗传多样性保护已成为长江上游水电开发过程中受关注的热点之一。
中华金沙鳅(Jinshaiasinensis)和短身金沙鳅(J.abbreviata)是长江上游的特有鱼类,是金沙鳅属仅有的两个种[2],其中中华金沙鳅也是“长江上游珍稀特有鱼类国家级自然保护区”的保护对象之一[3]。两种金沙鳅均为底栖、杂食性、喜激流、产漂流性卵鱼类,其生活类型是长江上游最主要的生态类群,具有代表性。两种金沙鳅不仅有相似的形态特征和生活习性,而且分布区域基本重叠,主要分布在长江上游干流、金沙江、雅砻江、嘉陵江和岷江[2,4]等水域。但是在资源量上,中华金沙鳅和短身金沙鳅有明显差别,根据鱼类早期资源调查,中华金沙鳅规模大于短身金沙鳅[4],表明这两种鱼对环境的适应力存在差异。因此研究群体遗传多样性有助于阐明近缘同域分布种类的适应性进化。本研究采用线粒体DNA (mtDNA)控制区序列对长江上游两种金沙鳅遗传多样性进行分析,旨在了解两种金沙鳅的遗传多样性现状,为长江上游金沙鳅属鱼类的资源保护提供科学依据。
1 材料与方法
1.1 材料
2016-2018年在长江上游8个样点采集中华金沙鳅136尾,3个样点采集短身金沙鳅66尾。样本采集地见图1,样本量见表1。所有样本经形态鉴定[2]后剪取少量鳍条并保存于无水乙醇中备用。

图1 中华金沙鳅和短身金沙鳅采样点及中华金沙鳅线粒体DNA控制区谱系分布Fig.1 Sampling sites and mtDNA clad distribution of J.sinensis and J.abbreviate
1.2 基因组DNA 提取、PCR 扩增及测序
采用高盐法提取基因组DNA[5]。线粒体控制区PCR扩增采用申绍祎等[6]研究所用引物。PCR的扩增体系为50 μL,包含2×PCR Mix(北京擎科新业生物技术有限公司)25 μL,上下游引物(10 pmol/L)各2 μL,DNA模板2 μL,灭菌去离子水加至50 μL。PCR扩增条件:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸60 s,35个循环;72 ℃再延伸8 min。PCR结束后,取2.5 μL产物,用1.0%琼脂糖凝胶电泳检测PCR,将扩增效果良好的PCR产物送武汉天一辉远生物科技有限公司进行双向测序,测序引物与PCR引物相同。
1.3 数据处理和分析
使用Lasergene v7.1软件包中的SeqMan对双向测序结果进行序列拼接,Clustal X[7]软件进行对位排列。用 MEGA v6.0[8]分析软件统计序列的平均碱基组成、计算转换/颠换比率。在DnaSP v5.0[9]软件中获得单倍型数(H)、变异位点数(S)、简约信息位点数,计算单倍型多样性指数(Hd)和核苷酸多样性指数(Pi)等参数,并计算群体间的K2P遗传距离,。
应用Arlequin v3.11[10]软件计算群体间的遗传分化指数(FST)值,进行分子方差分析(AMOVA),设1 000次重复随机抽样作显著性检验。用公式Nm=(0.5/FST)-0.5计算群体间基因流[11]。在MEGA v6.0[7]分析软件中构建单倍型邻接(NJ)发育树,bootstrap验证各分支支持率,设置为1 000。同时采用MrBayes v 3.1.2[12]软件对单倍型之间的系统发生树进行了贝叶斯推理(BI),最佳碱基替换模型由在线软件IQtree (http://iqtree.cibiv.univie.ac.at/)获得。使用Network5.0程序[13]构建单倍型网络图,对单倍型进化网络关系进行分析。
在DnaSP v5.0软件中计算中性检验值Tajima′s D和Fu′s Fs。用Beast 2.4.3.0[14]进行Bayesian Skyline Plots (BSP)分析种群历史,MCMC设为600 000 000,所得结果由TRACER ver.1.4 读取,各项参数的ESS均大于200。
2 结果与分析
2.1 序列变异
序列比对后共获得中华金沙鳅136条控制区序列,有效序列长度为915 bp,A、T、C、G的平均含量为35.63%、30.19%、20.02%和14.15%,具有明显的反G含量倾向,Ts/Tv =23.63,转换数明显高于颠换数。共获得66尾短身金沙鳅控制区序列,比对后得到有效长度为908 bp,A、T、C、G的平均含量为35.82%、31.39%、19.49%和13.30%,Ts/Tv =12.87,转换数也明显高于颠换数。
中华金沙鳅序列中共发现132个变异位点,其中39个为单一突变位点,93个为简约信息位点。136条序列共定义了131个单倍型,其中频率在2以上的单倍型只有5个,其余单倍型的频率均为1。总样本核苷酸多样性(Pi)和单倍型多样性指数(Hd)分别为0.018 2和0.999(表1),各群体之间的遗传多样性差别较小。短身金沙鳅序列仅检测到2个变异位点,定义了3个单倍型,Pi和Hd分别为0.000 5和0.411(表1)。

表1 中华金沙鳅和短身金沙鳅采样点、样本量、遗传多样性及中性检验值Tab.1 Sampling sites,sample size,genetic diversity parameters and neutral test of J.sinensis and J.abbreviate populations
注:*表示显著性检验P<0.05,**表示显著性检验P<0.01。
2.2 遗传结构
基于K2P模型计算,中华金沙鳅种内单倍型之间的遗传距离为0.001 1~0.039 6,短身金沙鳅种内单倍型之间的遗传距离为0.001 1~0.002 2,这两种鱼种间的遗传距离为0.035 7~0.051 5。
中华金沙鳅群体间分子变异方差分析(AMOVA)结果显示,来自群体间遗传变异组成占-0.75%,来自群体内的变异组成占100.75%,群体总的遗传分化指数(FST)为-0.007 53,说明中华金沙鳅群体遗传变异主要来自群体内,群体间未出现显著遗传分化(表2)。
短身金沙鳅群体间AMOVA分析结果也显示群体间未发生显著遗传分化,其变异主要来源于群体内(98.72%),且FST< 0.05 (FST=0.012 78)(表2)。
分别计算两两群体间的FST,结果显示,中华金沙鳅除NN及QJ与部分群体间的FST值大于0.05外,其余群体间FST均小于0.05(表3)。短身金沙鳅显示,除BN与HJ群体间FST大于0.05外,其余群体间FST均小于0.05(表4)。

表2 中华金沙鳅和短身金沙鳅群体间分子变异分析Tab.2 Analysis of molecular variance (AMOVA)among populations of J.sinensis and J.abbreviata
中华金沙鳅和短身金沙鳅群体间基因流分别见表3和表4。这两种鱼群体间的Nm值(或绝对值)均大于4(除JJ与NN、BN与HJ外)表明群体间基因交流比较频繁。
利用Median Joining方法构建中华金沙鳅单倍型网络结构图(图2),可见单倍型关系复杂,无明显的中心单倍型。根据连接单倍型之间的突变步长,可将单倍型划分为3个谱系:Clad I、Clad II、

表3 中华金沙鳅群体间的遗传分化(对角线下方)和基因流Nm值(对角线上方)Tab.3 Pairwise FST (below diagonal)and Nm values (above diagonal)among populations of J.sinensis

表4 短身金沙鳅种群间的遗传分化(对角线下方)和基因流Nm值(对角线上方)Tab.4 Pairwise FST (below diagonal)and Nm values (above diagonal)among populations of J.abbreviata
Clad III谱系间的步长为10和11,谱系内相邻单倍型之间突变步长都低于10(除了Clad II内的一个单倍型)。以犁头鳅(Lepturichthysfimbriata,GenBank登录号:DQ105283.1)作为外类群,构建两种金沙鳅单倍型NJ树和BI树(图3)。两棵树显示相似的拓扑结构,中华金沙鳅和短身金沙鳅的单倍型分为两个独立分支,形成两个单系群。中华金沙鳅的单倍型又分为3个具有较高支持率的支系。这3个支系包含的单倍型与中华金沙鳅的Network网络结构一致。
2.3 种群历史
通过中性检验Tajima′s D和Fu′s Fs等方法分析种群历史[15],中华金沙鳅结果显示,总群体以及3个谱系群体的Tajima′s D值均不显著,而Fu′s Fs检验中均为极显著负值(表1)。Bayesian skyline plot (BSP)进一步分析表明(图4),中华金沙鳅总群体和3个谱系群体的扩张时间分别约在距今0.12~0.17、0.1~0.12和0.12~0.17百万年(Ma)。短身金沙鳅群体由于序列变异位点和单倍型数量少,不作种群历史分析。
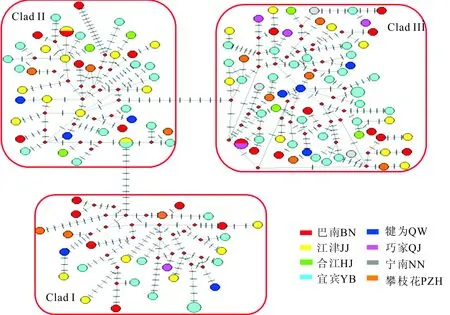
图2 基于线粒体DNA控制区序列构建的中华金沙鳅单倍型网络结构图(短线代表增加一个突变步骤)Fig.2 Haplotype network of J.sinensis based on mtDNA control region sequences

图3 基于线粒体DNA控制区序列的中华金沙鳅和短身金沙鳅单倍型NJ树(A)和BI树(B)(以犁头鳅作外类群)Fig.3 Haplotype NJ phylogenetic tree (A)and BI phylogenetic tree (B)based on mtDNA control region sequence of J.sinensis and J.abbreviata (L.fimbriata was used as outgroup)

图4 中华金沙鳅的BSP分析Fig.4 BSP analysis of J.sinensis population
3 讨论
3.1 群体遗传多样性
本研究显示,中华金沙鳅具有很高水平的线粒体遗传多样性,与Duan等[16]采用微卫星标记研究的结果一致,但该研究仅对雅砻江里庄31尾样本进行了分析,本研究的采样点和样本量更为丰富,包含Duan等[16]研究的江段。长江上游已开展的一些鳅科鱼类同样显示出高水平的遗传多样性,如中华沙鳅(Hd=0.986,Pi=0.003 65)[17]、小眼薄鳅(Hd=0.958,Pi=0.004 20)[18]、红唇薄鳅(Hd=0.907,Pi=0.003 15)[6]、长薄鳅(Hd=0.916,Pi=0.004 50)[19]等,与长江鲤科、鲿科等鱼类遗传多样性高低不同,高遗传多样性可能是长江上游鳅科鱼类线粒体DNA共有特征,更快积累变异。但是短身金沙鳅遗传多样性水平却明显较低,低水平的遗传多样性通常是由于近亲繁殖或小种群导致[20]。鱼类早期资源调查发现,金沙江干流、长江上游干流、雅砻江和岷江等均存在中华金沙鳅和短身金沙鳅产卵场,表明它们是同域分布的,但所有产卵场的中华金沙鳅产卵规模均大于短身金沙鳅[4],说明短身金沙鳅的种群规模比中华金沙鳅小。种群小容易近交,发生遗传漂变,导致遗传多样性降低。
3.2 种间及种群分化
长江大多数鱼类的种类鉴别可以通过DNA条形码(mtDNACOI基因)成功解决,但中华金沙鳅和短身金沙鳅例外[21]。这两种鱼之间Cytb基因的遗传距离也很低(Cytb:0.008±0.001),系统发育关系上出现种间单倍型嵌套,无法获得确定的种间区分[22]。本研究采用线粒体控制区分析的结果显示,两种金沙鳅的种间单倍型遗传距离达到0.035以上,虽然与中华金沙鳅种内单倍型之间遗传距离存在部分重叠,但NJ系统发育树可以将它们分开(图3)。这说明线粒体控制区比COI和Cytb基因对金沙鳅种类鉴别具有更高的区分度,但遗传距离上仍然没有明显种间间隔,也说明了这两种鱼物种分化时间比较晚。
AMOVA分析显示,中华金沙鳅和短身金沙鳅群体均未发生地理遗传分化,可能与其产卵特征有关。这两种鱼均产漂流性卵,受精卵随水漂流较长的距离而发育,成体向各栖息地扩散,促进了不同群体间的基因交流。长江上游一些产漂流性卵鱼类的遗传结构也表现出类似的同质性,如长鳍吻鮈[23]和异鳔鳅鮀[24]等。
尽管未检测到地理遗传分化,但中华金沙鳅单倍型系统发育树(图2)显示形成了三个具有较高支持率的谱系,表明中华金沙鳅发生群体内遗传分化。中华金沙鳅这一遗传结构特征符合Avise等[25]提出的线粒体系统地理学的第四类模式,即具有弱的同域单倍型谱系分化,但没有显著地理分化,是由于缺少长期亚群体隔离和高水平基因流,或群体扩张和单倍型广泛分布引起。长江上游一些鱼类就具有相似的遗传结构,如小眼薄鳅[18]和异鳔鳅鮀[24]等。短身金沙鳅单倍型没有出现谱系分化,可能与其单倍型数少,单倍型间序列差异小有关。
3.3 种群历史
BSP分析结果显示中华金沙鳅群体在10万~17万年前发生扩张,这一时期刚好是在第四纪冰期的庐山亚冰期(20万~23万年前)与大理亚冰期(1万~11万年前)之间的间冰期,间冰期相对于冰期温暖[26],有利于物种生存,也有利于物种从冰期的“避难所”向外扩散,发生种群扩张。研究表明,长江上游的一些鱼类像圆筒吻鮈[27]和铜鱼[28]等,也受到冰期影响,其种群在历史上也发生过瓶颈和扩张现象。中华金沙鳅在历史上经历了快速扩张,群体内可能积累了大量变异,与其较高的遗传多样性结果相吻合。
3.4 保护建议
本研究发现两种金沙鳅均没有发生地理遗传分化,为单一的进化显著单元,建议就地保护。目前长江上游已经建成向家坝和溪洛渡等大型水电工程,正在建设或规划建设更多梯级水电站,这将使中华金沙鳅和短身金沙鳅等的生境压缩和碎片化,阻碍群体基因交流。因此,今后还需加强对长江上游两种金沙鳅等鱼类产卵场和遗传多样性的监测,特别是对群体小、遗传多样性较低的短身金沙鳅的监测和保护。
