甘蓝型油菜GPDH基因家族的全基因组鉴定
2020-05-11付三雄周小婴王璐璐杨克相戚存扣
张 超,付三雄,唐 容,周小婴,黄 莎,王璐璐,杨克相,戚存扣
(1.贵州省农业科学院 贵州省油料研究所,贵阳 550006;2.江苏省农科院 经济作物研究所,南京 210014)
3-磷酸甘油脱氢酶(glycerol-3-phosphate dehydrogenase,GPDH)的主要作用是将糖酵解的中间产物磷酸二羟丙酮(dihydroxyacetone phosphate, DHAP)加氢还原成甘油3磷酸(glycerol-3-phosphate,G3P),是植物体内G3P合成的限速酶之一[1-2]。在酵母和衣藻等微生物中的研究表明,GPDH在维持细胞内的甘油平衡及促进油脂,即三脂酰甘油(triacylglycerol, TAG)的合成方面具有重要作用[3-5];在高等植物甘蓝型油菜(Brassicanapus L.)中过量表达酵母胞质型GPD1基因,发现转基因油菜种子中TAG的合成量增加了40%,且糖酵解的中间产物DHAP迅速减少[6],将种子特异表达的AtDGAT1和酵母胞质型GPD1共表达载体转到亚麻芥中,结果显示转基因株系种子含油量比野生型提高13%[7]。此外,近年来的研究还发现GPDH在调节渗透压[8]、防御病原菌侵害[9]及增强对盐分等逆境胁迫的适应[10]方面也具有重要的作用。
在模式植物拟南芥中有5个GPDH基因,根据亚细胞定位分为3类,其中有2个基因为质体型(plastidic glycerol-3-phosphate dehydrogenase,GPDHp),2个基因为细胞质型(cytosolic glycerol-3-phosphate dehydrogenase,GPDHc),另外1个则为线粒体型(mitochondrial glycerol-3-phosphate dehydrogenase,GPDHm)[11-14]。AtGPDHm主要在线粒体膜上将G3P氧化成DHAP,与AtGPDHc协同作用形成G3P穿梭机制,维持细胞内的G3P动态平衡[12-13],AtGPDHm在种子萌发时表达较高,并受ABA及逆境胁迫的影响[12],在盐生杜氏藻中发现盐胁迫能够促进GPDHm的表达,而缺氧和低温能够抑制其表达[15],推测GPDHm可以通过调控G3P氧化成DHAP这一释放能量的代谢过程来应对不同的胁迫条件。AtGPDHp的N端序列具有典型的叶绿体信号肽,且AtGPDHp与AtGPDHc的氨基酸序列差异较大[11],在水稻中过量表达质体型基因AtGLY1显著提高了水稻质体脂类的含量,而对内质网途径合成的脂类含量没有明显影响[16],可见GPDHp与GPDHc可能存在功能分化;另外,由于TAG的合成是在细胞质的内质网上进行的,相关研究已经证实过量表达GPDHc能够显著促进TAG合成[6-7],表明GPDHc在参与种子存储物质TAG的合成中的作用更关键。
目前蓖麻[17]、水稻[18]、玉米[10, 19]的GPDH已有研究报道,在甘蓝型油菜中也有部分GPDH被克隆和鉴定[20-21],但针对BnGPDH家族的全基因组鉴定和分析尚未见报道,尤其是BnGPDHc与油菜种子TAG合成的关系尚不明确,因此,本研究利用已公布的甘蓝型油菜参考基因组对BnGPDH进行了全基因组鉴定,全面分析BnGPDH基因结构、氨基酸保守结构域、系统进化树、染色体分布,采用实时荧光定量PCR(quantitative real-time PCR, qRT-PCR)的方法分析BnGPDHc在甘蓝型油菜不同组织器官中的表达模式,并比较BnGPDHc在高、低含油量自交系材料中的表达差异,为研究BnGPDH在油菜种子TAG积累中的作用奠定基础。
1 材料与方法
1.1 材 料
供试的甘蓝型油菜高含油量自交系‘IL130’‘IL474’‘IL594’及低含油量自交系‘IL134’‘IL202’‘IL525’由贵州省农业科学院贵州省油料研究所提供,2017-2018年度种植于贵阳试验地,于盛花期标记开放花朵,并取盛花期的根(Root)、茎(Shoot)、叶(Leaf)、花(Flower)、蕾(Bud)以及7、14、21、30、40DAF(day after flower)角果皮(Pericarp)和种子(Seed),液氮速冻后,于-80 ℃保存,用于基因组织表达模式分析和差异表达分析。
1.2 甘蓝型油菜、白菜和甘蓝中GPDH基因的鉴定
在拟南芥TAIR(http://www.arabidopsis.org/)网站下载AtGPDH蛋白序列,利用BlastP[22]比对Brassicadatabase (BRAD, http://brassicadb.org/brad/index.php)[23]中公布的甘蓝型油菜、白菜(BrassicarapaL.)、甘蓝(BrassicaoleraceaL.)基因组测序数据,获得BnGPDH、BrGPDH及BoGPDH蛋白序列,利用Pfam数据库(http://pfam.xfam.org/)中蛋白质结构域分析程序核对确认;基于在线网站ExPASY(https://web.expasy.org/compute_pi/)[24]预测蛋白质氨基酸数量、相对分子量和等电点。
1.3 BnGPDH的系统进化树构建、基因结构、保守结构域分析及染色体定位
运用MEGA 7.0软件[25]的ClustalW进行序列比对并采用邻接法(neighbour-joining, NJ)构建系统进化树,参数设置bootstrap重复1 000次;在BRAD数据库中获得BnGPDH、BrGPDH及BoGPDH的基因组DNA序列、编码区(coding sequenc,CDS)序列、内含子、外显子等物理位置信息,用Gene Structure Display Server (http://gsds.cbi.pku.edu.cn/)[26]分析BnGPDH基因结构;利用MEME(http://meme-suite.org/)[27]进行BnGPDH蛋白序列的保守元件分析,最大基序检索值设置为10,其他设置采用默认;使用Mapchart 2.2软件[28]对BnGPDH的染色体位置进行展示。
1.4 RNA提取与qRT-PCR分析
采用生工生物工程(上海)股份有限公司EZ-10 DNA away RNA提取试剂盒提取样品RNA,浓度测定和质量检测后用大连宝生物工程有限公司的PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒合成第一链cDNA,并按照SYBR○RPremix Ex TaqTMⅡ试剂盒说明在CFX96荧光定量检测系统(Bio-Rad,美国)上进行qRT-PCR分析,反应条件为:95 ℃预变性 90 s,接着95 ℃变性10 s,60 ℃退火延伸35 s,然后采集荧光信号,共40个循环,循环结束后从 60 ℃升温到95 ℃进行融解曲线分析;运用CT法计算相对表达量[29];内参基因采用BnUBC21(EV086936);每个反应3个生物学重复,3次技术重复;qRT-PCR所用引物见表1。

表1 qRT-PCR引物Table 1 Primers used in qRT-PCR
2 结果和分析
2.1 GPDH基因的全基因组鉴定和系统进化树构建
GPDH基因在甘蓝型油菜、甘蓝和白菜中分别鉴定出18、10和9个成员。18个BnGPDH基因中,gDNA长度从2 058 bp(BnGPDHp1-3)到 3 893 bp(BnGPDHm-4)不等,对应的CDS序列长度也是从1 071 bp变化到1 896 bp;相对分子质量最低的是BnGPDHp1-1的38.88 ku,最高的为BnGPDHm-2的67.93 ku;等电点从6.08(BnGPDHp1-1)到9.03(BnGPDHm-1)(表2)。
对甘蓝型油菜、白菜、甘蓝和拟南芥4个物种的GPDH蛋白进行序列分析后构建系统进化树显示,GPDH蛋白被分为3个亚家族(图1):GPDHp亚家族、GPDHc亚家族和GPDHm亚家族。3个亚家族中均包含6个BnGPDH和3个BrGPDH,其中GPDHp亚家族和GPDHc亚家族均有2个AtGPDH和3个BoGPDH,而GPDHm亚家族有1个AtGPDH和4个BoGPDH。
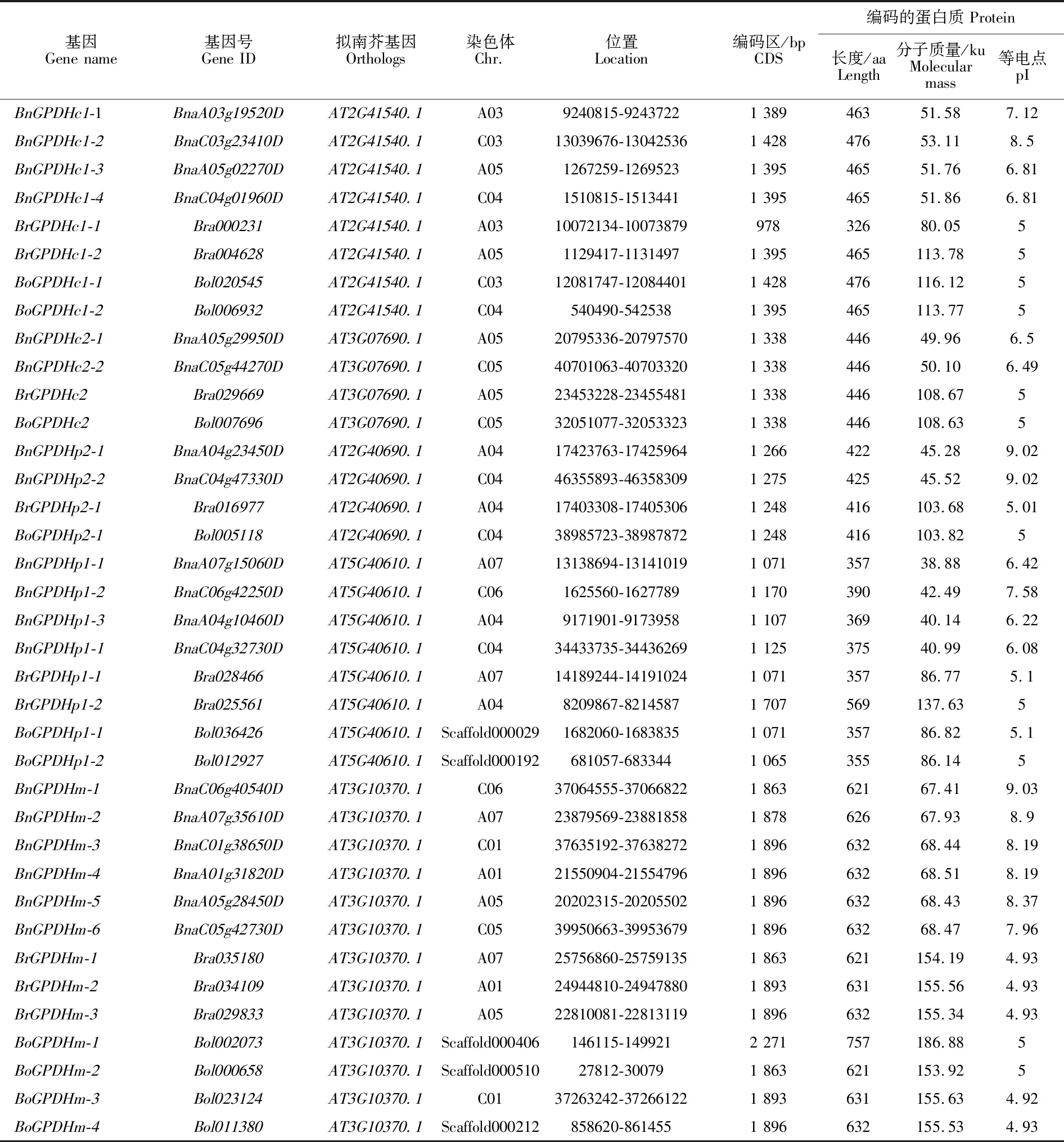
表2 甘蓝型油菜、白菜、甘蓝GPDH基因Table 2 GPDH genes in Brassica napus L., Brassica rapa L., Brassica oleracea L.
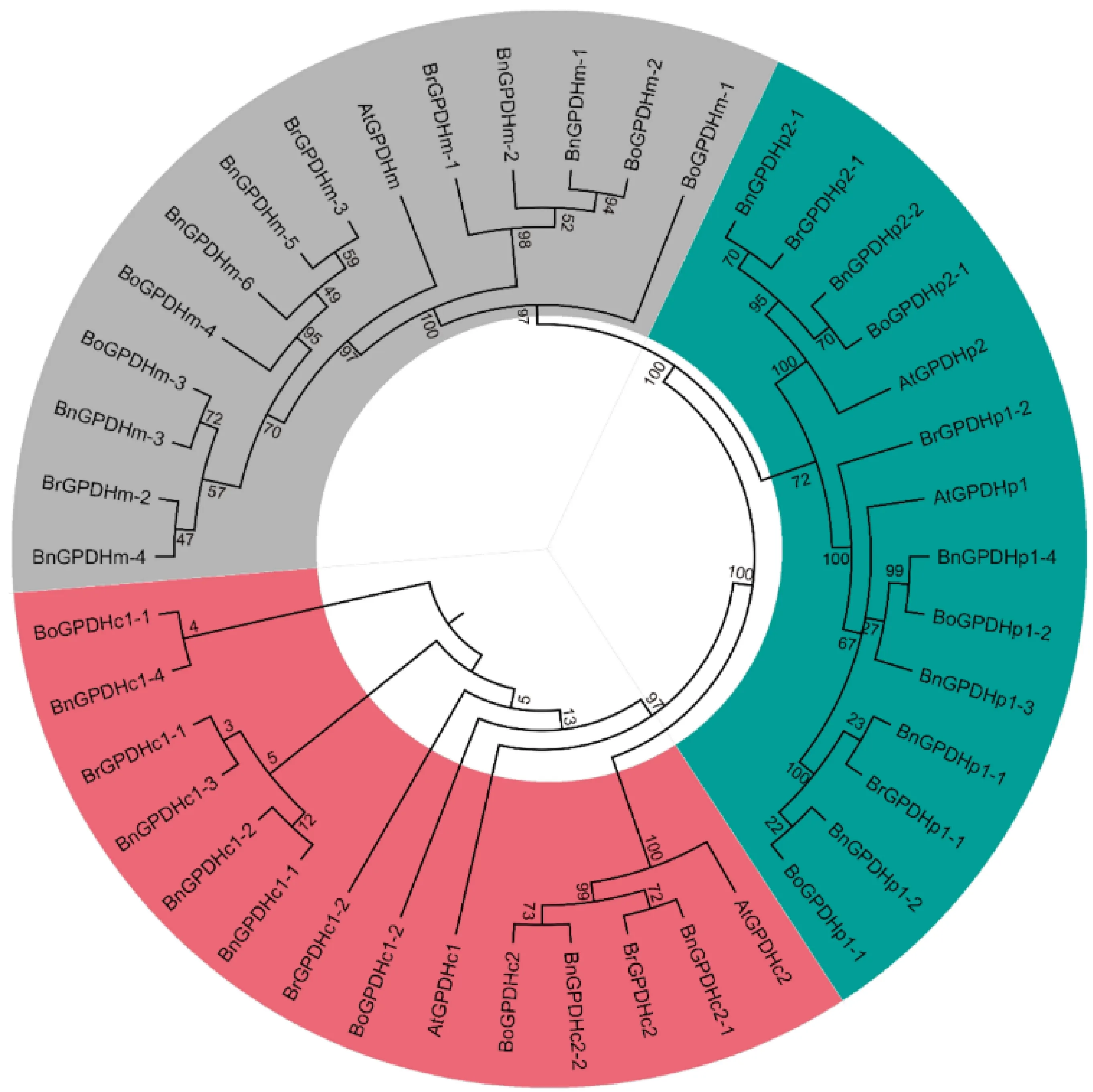
图1 甘蓝型油菜、白菜、甘蓝及拟南芥GPDH系统进化树分析Fig.1 Analysis of phylogenetic tree of B.napus, B.rape, B.oleracea, and Arabidopsis GPDH proteins
2.2 BnGPDH的染色体位置、基因结构和保守结构域分析
染色体位置分布显示,18个BnGPDH分布在A、C基因组的10条染色体上,其中,A01、A03和C01、C03染色体上只分布有1个BnGPDH基因,A04、A07、C05、C06染色体上均分布有2个BnGPDH基因,而A05和C04上各分布3个BnGPDH基因(图2)。
基因的结构分析表明,BnGPDHp亚家族中的外显子数目明显多于BnGPDHm和BnGPDHc两个亚家族,BnGPDHp亚家族中BnGPDHp2-1和BnGPDHp2-2具有8个外显子,另外4个成员外显子数目都为10个,而BnGPDHc亚家族中除了BnGPDHc1-2有6个外显子,其余5个成员都有5个外显子,BnGPDHm家族中BnGPDHm-1和BnGPDHm-2具有5个外显子,另外4个成员有6个外显子(图3-A),同一个亚家族内的基因成员外显子和内含子数量相对 一致。
保守结构域分析总共在BnGPDH蛋白中预测出20个保守元件,其中,只有元件2和元件7在所有BnGPDH中都存在,元件6、10、13、15、17只存在于BnGPDHm亚家族成员中;除了BnGPDHc2中的两个成员不包含元件20外,BnGPDHc亚家族中的6个成员都具有元件2、3、4、5、7、8、9、11、12、14、16和20;BnGPDHp亚家族成员具有较少的元件,其中BnGPDHp1中的4个成员基因都具有相同的元件,即元件1、2、4、7、11、12、14、18、19和20,BnGPDHp2中的两个成员只有元件2、7、18与BnGPDHp1成员相同;而BnGPDHm亚家族中的6个成员都包含16个同样的元件(图3-B),可见亚家族内的成员具有比较一致的元件,而亚家族间的保守元件差异较大。
2.3 BnGPDHc的表达模式分析
为了明确BnGPDHc组织表达特征,采用qRT-PCR检测了BnGPDHc在高含油量自交系IL130盛花期的根、茎、叶、花、蕾,以及7、14、21、30、40DAF的角果皮和种子中的表达情况,结果显示,BnGPDHc亚家族中的不同成员在不同组织器官中的表达量各不相同;BnGPDHc1-1、BnGPDHc1-2和BnGPDHc2-1、BnGPDHc2-2在各个组织器官中均有表达,且这4个基因在不同发育时期种子中的表达趋势一致:在7DAF中相对表达量较高,在14DAF略有下降,到21DAF、30DAF达到表达高峰,而40DAF则开始下降;此外,这4个基因在角果发育前期(7DAF、14DAF、21DAF)角果皮中的相对表达量要略高于后期(30DAF、40DAF)。BnGPDHc1-3和BnGPDHc1-4具有明显的组织表达特异性,BnGPDHc1-3在蕾、花中相对表达量较高,其次是21DAF、30DAF和40DAF的种子中,而BnGPDHc1-4在30DAF、40DAF的种子和根中的相对表达量较高,在其余的组织部位中几乎不表达(图4)。总体而言,BnGPDHc的6个成员在21DAF、30DAF、40DAF的种子中的表达更为活跃。

图2 BnGPDH在甘蓝型油菜染色体上的位置Fig.2 Chromosomes location of BnGPDH genes
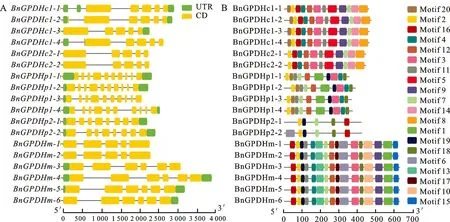
图3 BnGPDH基因结构(A)和BnGPDH蛋白保守元件(B)Fig.3 Gene structures of BnGPDH genes (A) and conserved motifs of BnGPDH(B)
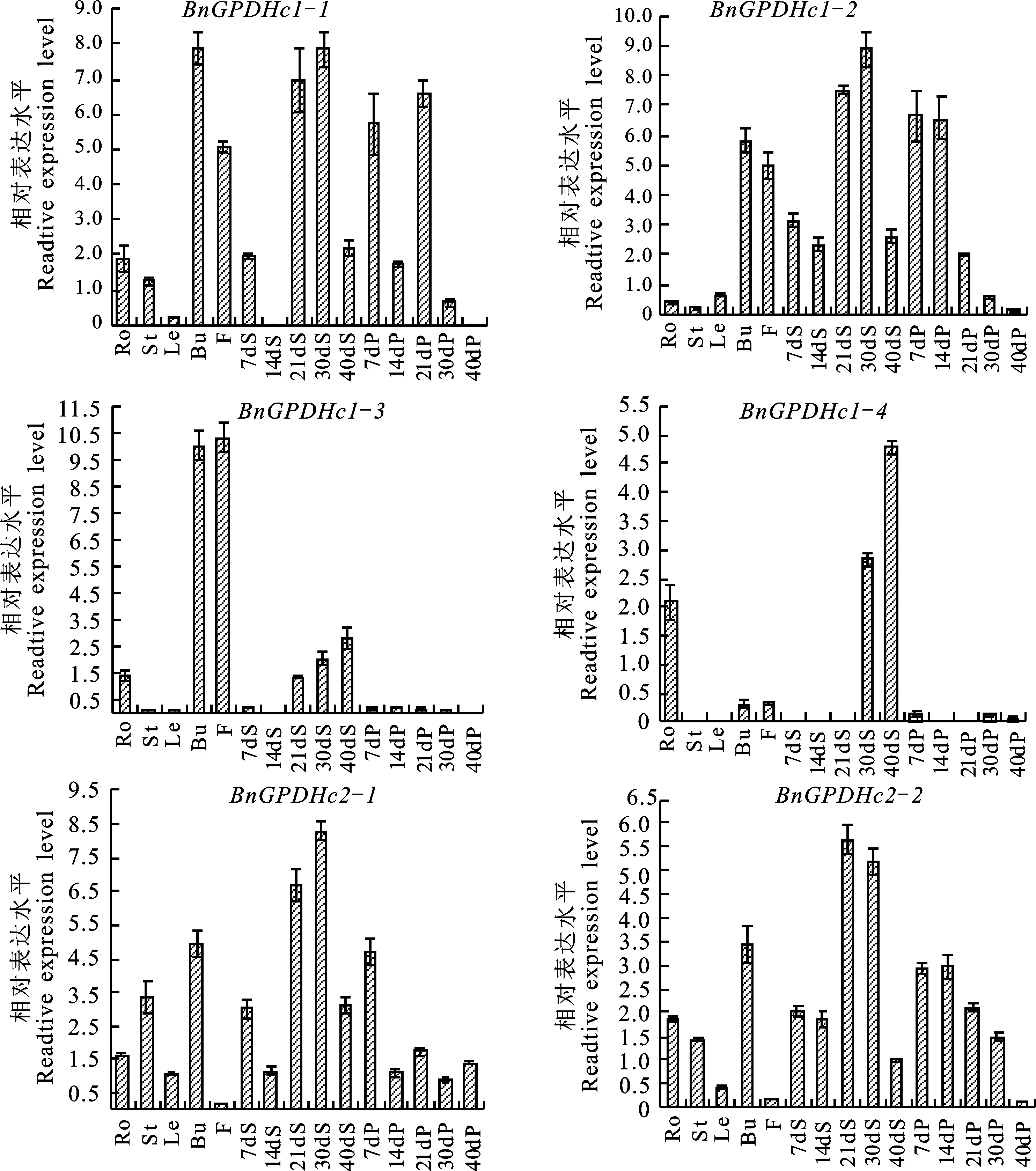
Ro.根;St.茎;Le.叶;Bu.蕾;F.花;7dS.7DAF种子;14dS.14DAF种子;21dS.21DAF种子;30dS.30DAF种子;40dS.40DAF种子;7dP.7DAF角果皮;14dP.14DAF角果皮;21dP.21DAF角果皮;30dP.30DAF角果皮;40dP.40DAF角果皮(下同)
Ro.Root;St.Stem;Le.Leaf;Bu.Bud;F.Flower;7dS.Seed of 7DAF;14dS.Seed of 14DAF;21dS.Seed of 21DAF;30dS.Seed of 30DAF;40dS.Seed of 40DAF;7dP.Silique pericarp of 7DAF;14dP.Silique pericarp of 14DAF;21dP.Silique pericarp of 21DAF;30dP.Silique pericarp of 30DAF;40dP.Silique pericarp of 40DAF(the same below)
图4BnGPDHc在自交系IL130中的组织表达模式分析
Fig.4 Tissue-specific expression patterns ofBnGPDHcgenes in inbred line IL130
2.4 BnGPDHc在高、低含油量自交系中的差异表达分析
对BnGPDHc在3对高、低含油量自交系的不同组织器官中的相对表达量进行分析发现:在叶片和21DAF、30DAF、40DAF的角果皮中BnGPDHc的相对表达量较低,而在花和蕾中的相对表达量较高,但这些组织部位中BnGPDHc的相对表达量在高、低含油量材料间差异不显著;在21DAF的种子中BnGPDHc1-2和BnGPDHc2-1的相对表达量在高、低含油量材料间存在显著差异,30DAF的种子中BnGPDHc2-1和BnGPDHc2-2在高含油量材料中的相对表达量要显著高于低含油量材料,40DAF种子中BnGPDHc的相对表达水平在高、低含油量材料间差异不显著(图5)。这些结果说明,BnGPDHc1-2、BnGPDHc2-1和BnGPDHc2-2可能与种子TAG的积累相关。
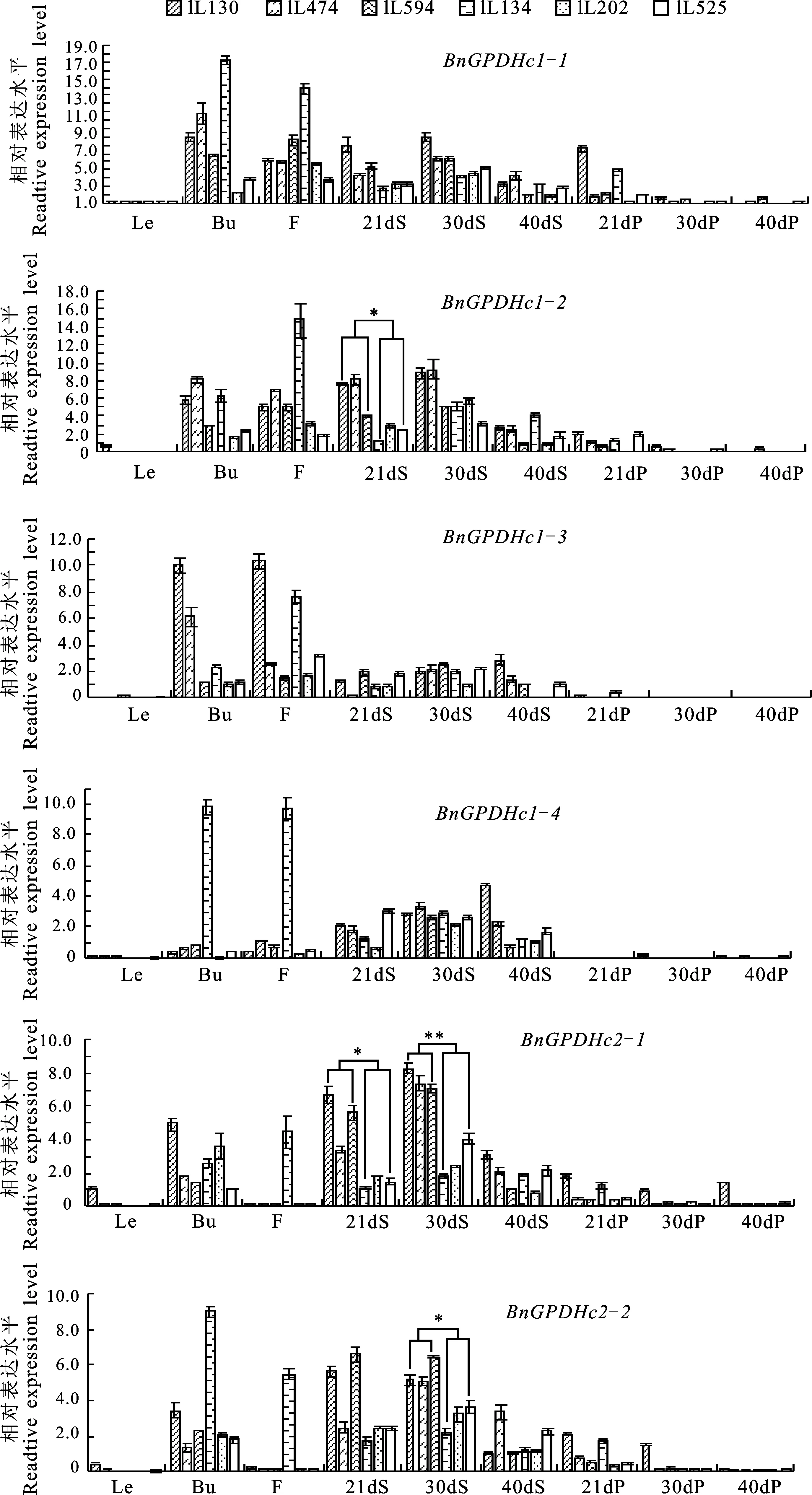
*表示差异显著(P<0.05);**表示差异极显著(P<0.01)
* Indicates significant difference atP<0.05;** Indicates significant difference atP<0.01
图5BnGPDHc在高、低含油量自交系不同组织器官中的差异表达
Fig.5 Differential expressions ofBnGPDHcgenes between high and low seed oil content inbred lines in different tissues
3 讨 论
GPDH在促进TAG的合成[5-7]、渗透压的调节[8,10]、防御病原菌侵害[9]等方面具有重要作用,在蓖麻[17]、水稻[18]、玉米[10,19]及甘蓝型油菜[20-21]中已有部分GPDH被克隆和鉴定,但针对BnGPDH家族的全基因组鉴定尚未见报道。甘蓝型油菜(AACC,2n=4x=38)是白菜(AA, 2n=2x=20)和甘蓝(CC, 2n=2x=18)2个二倍体祖先种通过天然远缘杂交形成的异源四倍体作物,理论上拟南芥的单个基因在白菜和甘蓝基因组上应该有3个拷贝,在甘蓝型油菜基因组上应该有6个拷贝[30],本研究在甘蓝型油菜、甘蓝、白菜和拟南芥中分别鉴定出18、10、9及5个GPDH基因,白菜和甘蓝进化成甘蓝型油菜的过程中仅有1个GPDH基因丢失,而拟南芥在进化成白菜和甘蓝的过程中却出现了更多的GPDH基因丢失,这一现象与高堃等[31]发现的甘蓝型油菜中PIN1s基因数量远少于甘蓝和白菜中PIN1s基因总数的现象有所不同,可见不同的基因在进化和遗传上具有一定的差异性。
起源于同一拷贝的基因拥有类似的基因结构和保守元件[32],例如对BnCKX家族的分析[33],本研究基因结构分析发现同一个亚家族内起源于同一个拟南芥基因的BnGPDH外显子和内含子数量相对一致;保守元件分析发现BnGPDHm的6个成员具有完全相同的保守元件,BnGPDHc的6个成员保守元件也高度相似,暗示BnGPDHm和BnGPDHc这2个亚家族内的基因功能分化较小,但是否存在功能冗余则需要进一步的试验证明;BnGPDHm亚家族又分为BnGPDHp1和BnGPDHp2两个子集,且这两个子集间保守元件存在较大差异,推测BnGPDHp1和BnGPDHp2可能分别由AtGPDHp1和AtGPDHp2独立进化而来,且存在一定的功能分化。
研究认为GPDHm主要与GPDHc协同作用维持细胞内的G3P动态平衡并参与线粒体的能量代谢过程[12-13,15],GPDHp则主要参与质体脂类的合成,对种子含油量影响不大[11,16,20],而GPDHc在细胞质TAG合成过程中作用更为关键[6-7]。因此,本研究对BnGPDHc进行qRT-PCR分析,结果发现BnGPDHc的6个成员在21、30、40DAF的种子中的表达更为活跃,与刘少锋等[34]的研究结果一致,油菜种子21DAF-40DAF是油分积累的关键时期,BnGPDHc的表达量提高可以促进G3P的合成,为TAG的合成提供充足的底物;研究还发现尽管BnGPDHc在光合器官叶片中的相对表达量较低,但BnGPDHc1-1、BnGPDHc1-2、BnGPDHc2-1、BnGPDHc2-2这4个成员在角果发育前期的角果皮中的相对表达量却较高,在水稻中的研究也发现OsGPDH1在生殖发育时期的剑叶中的表达量较高[18],推测GPDH可能参与了角果皮、剑叶等光合作用(源)器官的形态建成;高、低含油量自交系材料不同组织器官qRT-PCR分析显示21DAF 、30DAF种子中BnGPDHc1-2、BnGPDHc2-1、BnGPDHc2-2的相对表达量在高、低含油量自交系中存在不同程度的差异,与Lu等[35]对高含油品系‘ZS11’和低含油量品系‘WH5557’ 34DAF种子子叶进行转录组测序的结果相吻合,表明这些基因可能在油菜种子TAG合成中具有关键作用,可以作为油菜含油量改良的候选基因研究 利用。
本研究采用生物信息学分析的方法在甘蓝型油菜基因组中共鉴定出18个BnGPDH基因,分为BnGPDHp、BnGPDHm和BnGPDHc3个亚家族,基因结构及保守元件分析显示亚家族内的基因成员具有一定的保守性,亚家族间的成员具有较大差异,暗示亚家族间的基因存在功能分化,qRT-PCR分析发现BnGPDHc1-2、BnGPDHc2-1、BnGPDHc2-2可能在油菜种子TAG合成中的作用更关键,这些结果为进一步利用这些基因进行油菜含油量改良奠定了基础。
