低温胁迫后增温对‘罗宾娜’百合生理特性的影响
2020-04-23葛蓓蕾张瑞军
葛蓓蕾,张 萍,雅 蓉,张瑞军,靳 磊
(宁夏大学 农学院,宁夏 银川 750021)
【研究意义】‘罗宾娜’百合(‘Robina’)是宁夏地区温室中常见的切花品种,因花大色艳、观赏性高、抗病性强等优点备受消费者青睐。‘罗宾娜’百合的生长发育需要充足的热量,白天生长最适温度为20~25 ℃,夜间最适温度为14~15 ℃;温度过低会导致其生长停滞、花期延迟,而温度过高则会引起花蕾败育、开花率下降等症状[1]。目前由于宁夏百合栽培地温室保温效果不理想,常使冬季生长的‘罗宾娜’百合遭受低温胁迫,进而导致百合切花质量差、切花率低[2-3]。种植过程中,生产者常依据百合株型或花朵的变异等指标来判断温室温度是否过低,但此时百合大多已遭受低温胁迫,因此在何种受损程度下采取增温措施从而抑制其继续恶化(或恢复正常生长)将显得尤为重要。【前人研究进展】近几年,国内外针对百合低温响应的研究大多集中在低温胁迫后鳞茎的生理、表型及品质变化等方面,主要通过测定低温下百合的萌芽率、电导率、抗氧化酶活性、丙二醛以及可溶性糖含量等指标评价不同种质的抗寒差异及生理机制[4-9],对百合低温胁迫后增温修复的研究还鲜有报道,目前仅有李智辉等[10]对新铁炮百合幼苗期叶片进行低温胁迫和增温处理的研究,该研究发现增温一段时间能够恢复低温胁迫后百合叶片的活性氧产生速率、SOD和POD活性,但涉及的切花百合生长期较单一、指标较少,重点亦放在低温胁迫后切花百合的生理变化上,关于增温对其生理和品质是否具有一定恢复作用并未见讨论。【本研究切入点】通过人工气候室在不同生长期对 ‘罗宾娜’切花百合进行低温胁迫后增温处理。【拟解决的关键问题】探讨增温对不同生长期百合生理特性的影响,以期为冬季日光温室的调温及切花百合的种植管理提供技术参考,进而促进切花百合的冬季优质、高效种植。
1 材料与方法
1.1 供试材料
供试的百合品种为‘罗宾娜’,鳞茎周径为18~20 cm,购自昆明红日花卉种植有限公司。
1.2 试验方法
1.2.1 材料培养与处理 试验于2017年3-7月在宁夏大学林学实验室进行。在栽培箱(60 cm×40 cm)底部铺3~4 cm厚草炭与珍珠岩混合基质(比例为3∶1),3月31日种植60颗种球,分别在3个时期(10叶期、20叶期、现蕾期)各取20株用于温度处理,覆盖基质10 cm,基质pH值为6.5,根据基质水分状况每周浇水1~2次。
2个温度处理分别为:①室温处理(昼/夜温度22 ℃/14 ℃),作为对照(CK);②低温处理(昼/夜温度8 ℃/2 ℃),每种温度处理各10株。低温处理的百合首先在昼夜温度22 ℃/14 ℃条件下预处理1 d,测定各生理作为初始值,第2 天分别移入光照强度为400 μmol·m-2·s-1的光照培养箱内进行低温处理,光周期均设为白天15 h,黑夜9 h。低温处理10 d,分别在胁迫第3、7和10天取样测定各项生理指标,随后立即进行增温处理(温度增至室温),分别测定增温第3、7和10天的各项生理指标。试验期间各处理的环境条件及管理措施均保持一致。
1.2.2 生理指标测定 在低温和增温处理过程中,分别选取百合10叶期(2017年4月29日采样,每株10~12片叶)、20叶期(2017年5月19日采样,每株18~20片叶)和现蕾期(2017年6月24日采样)的中部叶用于其生理指标的测定,各重复测定3次。
采用氮蓝四唑(NBT)光化还原法测定超氧化物歧化酶(SOD)活性,愈创木酚法测定过氧化物酶(POD)活性,紫外吸收法测定过氧化氢酶(CAT)活性,酸性茚三酮比色法测定脯氨酸(Pro)含量[11];利用电导仪(DDS-307)测定相对电导率(REC);蒽酮比色法测定可溶性糖含量[12]。
1.3 数据处理
应用Microsoft Excel 2010和Origin 8.5软件分析并作图,数据以平均值±标准差表示。
2 结果与分析
2.1 增温对不同生长期低温胁迫百合叶片抗氧化酶活性的影响
2.1.1 超氧化物歧化酶(SOD) 在8 ℃/2 ℃(昼/夜温度)低温胁迫处理下,百合10叶期叶片SOD活性随胁迫时间的延长不断上升,胁迫10 d时达到183.92 U·g-1,是同期CK的2.19倍,20叶期叶片SOD活性随胁迫时间的延长呈上升-下降-上升趋势,胁迫3 d时出现最高值(138.75 U·g-1),是同期CK的1.80倍,现蕾期叶片SOD活性与同期CK相比无明显变化,胁迫10 d时为82.44 U·g-1,是同期CK的1.03倍。增至室温后,百合10叶期和20叶期叶片SOD活性均随处理时间的延长不断下降,但仍略高于各期CK,现蕾期叶片SOD活性随处理时间的延长呈先升后降趋势。增温10 d后10叶期、20叶期及现蕾期叶片SOD活性分别降到114.96、81.81、141.24 U·g-1,分别是各期CK的1.49、1.13、1.03倍。可见,百合各时期叶片SOD活性受温度影响的大小为:10叶期>20叶期>现蕾期,即现蕾期百合SOD活性在低温胁迫下波动较小,抗低温能力较强(图1,A~C)。综上,低温胁迫会诱导百合叶片SOD活性升高,而增温处理能有效缓解这种上升趋势,使叶片SOD活性下降,逐渐恢复到CK水平。
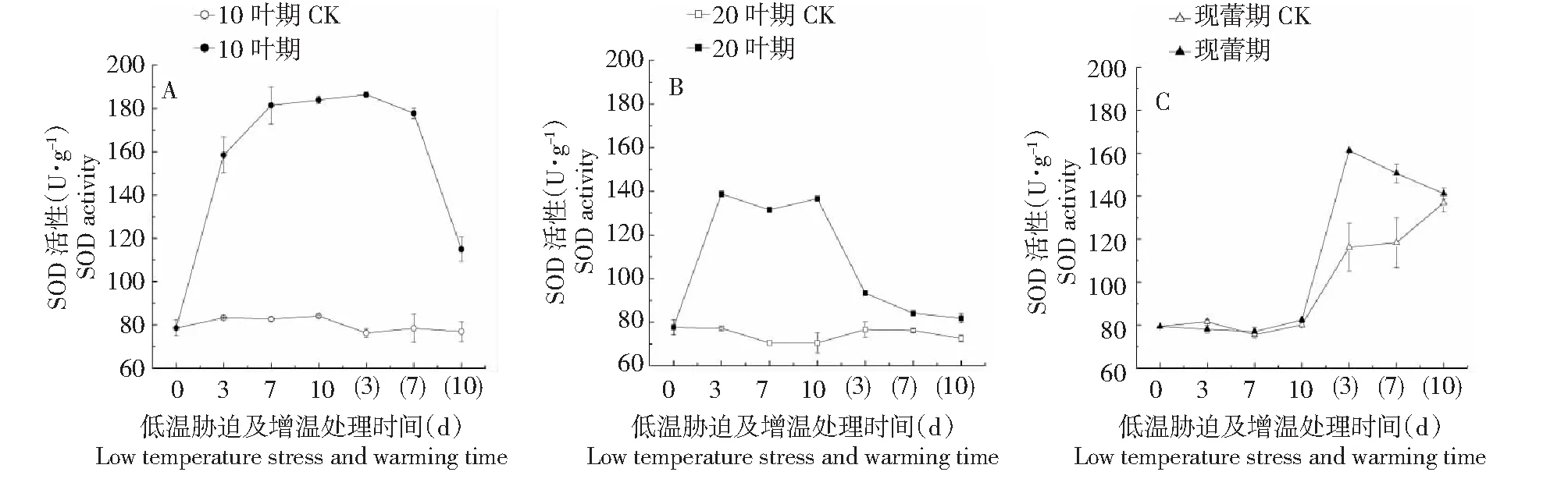
处理时间3、7、10代表低温胁迫第3、7、10天;(3)(7)(10)代表增温第3、7、10天,下同 Treatment time 3, 7, and 10 represent the third, seventh, and tenth days of low temperature stress; (3) (7) (10) represent the third, seventh, and tenth days of warming, the same as below
2.1.2 过氧化物酶(POD) 低温胁迫下,百合10叶期、20叶期及现蕾期叶片POD活性均随胁迫时间的延长不断升高,胁迫10 d时各期叶片POD活性分别升到49.70、25.69、30.38 U·g-1(FW)·min-1,分别是各期CK的5.48、2.96、3.29倍。增温后各时期叶片POD活性均随处理时间的延长不断下降,且逐渐趋于CK,增温10 d后各期叶片POD活性分别降到13.01、13.03、14.17 U·g-1(FW)·min-1,分别是各期CK的1.47、1.55、1.33倍。可见,百合各时期叶片POD活性受温度影响的大小为:10叶期>现蕾期>20叶期(图2,A~C)。综上,低温胁迫会诱导百合各期叶片POD活性升高,而增温能有效缓解这种上升趋势,使叶片POD活性下降,逐渐恢复到各期CK水平。
2.1.3 过氧化氢酶(CAT) 低温胁迫后,百合10叶期、20叶期和现蕾期叶片CAT活性均随胁迫时间的延长不断升高,胁迫10 d时达到峰值,分别为243.45、239.69、251.81 U·g-1(FW)·min-1,分别是各期CK的1.97、1.94、2.15倍。增温后各时期叶片CAT活性均随处理时间的延长不断下降,逐渐恢复CK水平。增温10 d后百合10叶期、20叶期和现蕾期叶片CAT活性分别降到134.89、137.00、137.05 U·g-1(FW)·min-1,分别是各期CK的1.14、1.11、1.13倍。可见,百合各时期叶片CAT活性受温度影响的大小为:现蕾期>10叶期>20叶期(图3,A~C)。综上,低温胁迫会诱导百合叶片CAT活性升高,而增温能有效缓解这种上升趋势,使叶片CAT活性下降,逐渐恢复CK水平。
2.2 增温对不同生长期低温胁迫百合叶片相对电导率(REC)的影响
百合10叶期叶片REC在低温胁迫前7 d大幅上升,随后开始下降,但始终高于同期CK,胁迫7 d时出现最高值(26.35 %),是同期CK的2.91倍;20叶期和现蕾期叶片REC均随胁迫时间的延长不断上升,胁迫10 d时达到峰值,分别为20.02 %和20.85 %,分别是各期CK的2.38和2.43倍。增温后各时期百合REC均随处理时间的延长不断下降,且逐渐趋于各期CK,增温10 d后百合各期REC分别降到12.45 %、10.95 %、11.03 %,分别是各期CK的1.24、1.39、1.25倍。可见,百合各时期叶片REC受温度影响的大小为:10叶期>20叶期>现蕾期(图4,A~C)。综上,低温胁迫会诱导百合叶片REC升高,而增温能有效缓解这种上升趋势,使REC下降逐渐趋于CK。

图2 低温胁迫后增温处理对切花百合‘罗宾娜’POD活性的影响

图3 低温胁迫后增温处理对切花百合‘罗宾娜’CAT活性的影响

图4 低温胁迫后增温处理对切花百合‘罗宾娜’相对电导率的影响
2.3 增温对不同生长期低温胁迫百合叶片脯氨酸(Pro)含量的影响
低温胁迫下,百合10叶期叶片Pro含量随胁迫时间的延长大幅上升,胁迫10 d时达到3.92 mg·g-1,是同期CK的1.81倍,20叶期和现蕾期叶片Pro含量趋于稳定,无大幅度波动,胁迫10 d时分别达到1.04和1.94 mg·g-1,是各期CK的0.72和0.82倍。增温后百合10叶期叶片Pro含量随处理时间的延长不断下降,20叶期叶片Pro含量仍趋于稳定,现蕾期叶片Pro含量随处理时间的延长呈小幅先升后降趋势。胁迫10 d后10叶期、20叶期及现蕾期百合叶片Pro含量分别为2.24、1.03、2.04 mg·g-1,分别是各期CK的1.05、0.70、0.94倍。可见,百合各时期叶片Pro含量受温度影响的大小为:10叶期>现蕾期>20叶期(图5,A~C)。综上,整个处理过程对20叶期和现蕾期百合叶片Pro含量影响不大,但对10叶期百合叶片Pro含量影响较大,即低温胁迫会诱导百合10时期叶片Pro含量大幅上升,且明显高于同期CK,而增温能有效缓解这种升高趋势,使Pro含量逐渐减少并趋于同期CK。
2.4 增温对不同生长期低温胁迫百合叶片可溶性糖含量的影响
低温胁迫后,百合10叶期叶片可溶性糖含量随胁迫时间的延长呈先升后降趋势,但仍高于同期CK,20叶期和现蕾期叶片可溶性糖含量均随胁迫时间的延长呈先降后升趋势,胁迫10 d时10叶期、20叶期及现蕾期百合叶片可溶性糖含量均高于各期CK,分别为1.02 %、1.52 %、1.36 %,是各期CK的1.35、1.82、2.59倍。增温后百合10叶期叶片可溶性糖含量随处理时间的延长呈先升后降趋势,20叶期和现蕾期叶片可溶性糖含量均随处理时间的延长大幅降低,增温10 d后各期叶片可溶性糖含量分别降到0.85 %、0.30 %、0.42 %,分别是各期CK的1.10、0.38、0.55倍。可见,百合各时期叶片可溶性糖含量受温度影响的大小为: 20叶期>现蕾期>10叶期(图6,A~C)。综上,低温胁迫会诱导百合各时期叶片可溶性糖含量上升,且明显高于各期CK,而增温能有效缓解这种升高趋势,使其不断减少并逐渐趋于各期CK。

图5 低温胁迫后增温处理对切花百合脯氨酸含量的影响

图6 低温胁迫后增温处理对切花百合可溶性糖含量的影响
3 讨 论
温度是影响百合生长发育的重要因子,尤其在低温胁迫下,百合的细胞膜和抗氧化等系统产生一系列变化,使其抗低温能力下降、生理机能发生改变,最终导致百合优质生产严重受限[13]。本研究中,低温胁迫下百合各时期叶片抗氧化酶活性、REC、Pro及可溶性糖含量均有所升高,增温后各指标不断下降逐渐趋于各期CK,这表明低温胁迫后增至室温对不同生长期‘罗宾娜’切花百合的生理活性有一定修复作用。
抗氧化酶活性是判断植物在低温胁迫环境中生理机能的重要指标[14-16]。本研究中,百合在不同生长期经低温胁迫后,其SOD、POD和CAT活性均大幅上升,这与罗萍等[17]研究结果一致,这些抗氧化酶活性的上升加强了清除H2O2的能力,从而使膜脂过氧化物减少,百合抗寒能力增强,增温后其叶片SOD、POD和CAT活性基本恢复至CK水平,这表明增温对低温胁迫后‘罗宾娜’切花百合的抗氧化酶活性有一定修复作用。
植物相对电导率(REC)是反映植物细胞膜透性的一项基本指标,低温胁迫会影响植物的REC。百合在低温胁迫下产生大量自由基与活性氧,从而引起细胞膜透性增大,电解质和小分子有机物外渗较多,REC升高,膜系统受到伤害[18]。本研究中,百合在不同生长期经低温处理后REC升高,这与王玲丽等[19]研究结果一致,增温后REC下降,百合叶片膜透性减小,电解质和小分子有机物外渗减少,这表明增温对低温胁迫后‘罗宾娜’切花百合的REC有一定修复作用。
脯氨酸(Pro)和可溶性糖是植物体内重要的有机渗透调节物质[20]。通常低温胁迫下植物为保持自身正常的生理代谢,常把淀粉转化成可溶性糖,然后主动的积累可溶性糖和Pro去调节细胞渗透势,保护体内所含的酶类以及细胞膜结构免受冻害[21]。本研究中,百合在不同生长期经低温处理后,其叶片Pro和可溶性糖含量均升高,这与金鑫等[22]研究结果一致,渗透调节物质的增加会引起细胞液浓度增大,渗透势降低,从而减小低温胁迫伤害,增温后百合叶片Pro和可溶性糖含量下降,逐渐趋于CK,这说明增温对低温胁迫后‘罗宾娜’切花百合的Pro和可溶性糖含量有一定修复作用。其中,温度变化对10叶期百合Pro含量影响最大,20叶期和现蕾期百合Pro含量与CK相比无显著变化,可能是因为幼龄时期百合生长发育不完全,易受低温影响,随着植株生长,其自身抗性不断增强,Pro含量不易受低温影响,但具体原因还有待进一步深究。
综上所述,增温对低温胁迫后不同生长期‘罗宾娜’百合的生理活性具有一定恢复作用,其中,百合叶片的SOD、POD、CAT、REC及可溶性糖含量在3个时期均对温度变化均较为敏感,可作为主要生理指标用于今后切花百合的抗寒研究,本研究结果为宁夏地区‘罗宾娜’切花百合的优质生产提供了技术参考,为增温恢复低温胁迫后切花百合的生理活性奠定了理论基础。
