雅砻江冬麻豆(Salweenia bouffordiana)种群结构与动态特征
2020-04-16张全建段晨松张远彬
杨 彪,张全建,龚 旭,段晨松,张远彬
1 西南野生动植物资源保护教育部重点实验室,西华师范大学,南充 637002 2 中国科学院、水利部成都山地灾害与环境研究所,成都 610041 3 中国科学院大学,北京 100049
种群应系指一个地区内同属于一个种的个体组成的群体[1],种群结构是种群最基本的特征,是种群生态学研究的核心问题之一[2- 3]。极小种群是指地理分布狭小的种群,长期受到外界因素的干扰和压力,其数量小于防止灭绝所需的最低限度[4],极小种群具有剩余种群少、栖息地受限、人为干扰严重和绝种风险极高的特征[5]。目前,我国仅有120个极小种群被关注[4- 5],众多处于濒危状态的物种并未获得保护地位,而其作为生物多样性的重要组成,一旦灭绝将给人类带来不可估量的损失。当前,物种灭绝已是全球最严重的生态问题之一,直接威胁着人类社会的可持续发展[6],而保护濒危物种已成为一个紧迫的保护生物学课题[7]。中国作为世界上最大的生物多样性国家之一,保护中国最受威胁的野生植物物种比以往任何时候都更加迫切。
雅砻江冬麻豆(Salweeniabouffordiana)是豆科冬麻豆属(Salweenia)植物新种[8],为我国特有种,主要分布于四川省新龙县境内的雅砻江河谷干燥灌丛和砾石生境中[8]。由于本种的分布范围极其狭窄,种群数量稀少,根据世界自然保护联盟条例[9],有学者们将其保护等级定为极度濒危(Critically Endangered, CR)[8],《中国生物多样性红色名录——高等植物卷》将其定为濒危(Endangered, EN)物种[10]。由于缺乏对该物种的种群数量、种群结构等方面的研究,不利于后续关于该物种的相关保护策略的制定和行动的开展,需要对其现有种群数量、分布范围等进行调查与研究。
1 材料与方法
1.1 研究区概况
雅砻江冬麻豆为常绿丛生灌木,高约0.5—2 m,茎直立,被绒毛,与冬麻豆(S.wardii)的差别主要在于本种的小叶更大、瓣膜呈波浪状,主要分布于四川省新龙县境内的雅砻江河谷。本研究的研究区位于四川省甘孜藏族自治州新龙县,地理位置介于东经99°37′—100°54′E、30°23′—31°32′N之间,海拔2760—3232 m,地处雅砻江中游高山峡谷地带。该区属青藏高原型季风气候,具有独特的大陆性高原季风气候的特点。根据新龙县气象站多年统计资料可知,全县平均气温为7.5℃,极端最高温32.7℃,极端最低温-19.2℃,1月均温-2.3℃,7月均温15.1℃,≥10℃活动积温1887℃,相对湿度在40%左右,全年无霜期114 d。研究区水量分布不均匀,降雨主要集中于每年6月至9月。植被群落主要有河谷灌丛、亚高山暗针叶林、高山松林、高山柏林、栎类林、桦木林、亚高山草甸、高山草甸等类型。
1.2 调查方法
由于雅砻江冬麻豆主要生长在雅砻江河谷地带,故沿河岸1 km范围随机布设调查样线8条,沿每条样线记录发现的雅砻江冬麻豆、干扰信息和生境信息。在有雅砻江冬麻豆种群分布的地段设置5 m×5 m样方做常规的种群学调查,共计调查样方55个。样方调查时记录样方的经纬度、海拔、坡向、坡度等因子,乔木层、灌木层和草本层的植物种类、盖度和高度等植被状况,并对雅砻江冬麻豆的种群及生长状况进行常规种群学调查,记录样方内雅砻江冬麻豆的幼苗数量、盖度、高度、冠幅和地径等参数。
1.3 计算方法
1.3.1径级划分标准
对于木本植物尤其濒危物种而言,大多数学者认为在数量较少,较难获取其年龄的情况下,可用径级结构代替其年龄结构[11]。本研究选择生物指标特征显著的地径结构来代替年龄结构,为分析其种群结构的精细动态,采用李清河等对荒漠珍稀灌木半日花种群的径级划分方法[12]进行地径划分,即采用较小极差,每1 mm划分1个径级。根据地径(D)的分布状况,将地径划分为25个等级:Ⅰ级(3 mm≤D<4 mm)、Ⅱ级(4 mm≤D<5 mm)、Ⅲ级(5 mm≤D<6 mm)、……、XXIV级(26 mm≤D<27 mm)和XXV级(D≥27 mm)。以上述划分为标准,统计每个龄级雅砻江冬麻豆个体数,并以此进行种群动态量化分析和建立种群静态生命表。
1.3.2种群动态量化分析方法
种群动态量化采用陈晓德[13]的量化分析方法定量描述雅砻江冬麻豆种群动态,以Leak[14]的划分理论作为确定种群结构类型的依据,公式如下:
(1)
(2)
式中,Vn为种群从n到n+1级的个体数量变化;式(2)中Vpi为整个种群结构的数量变化动态指数(忽略外部干扰);Sn、Sn+1分别为第n和n+1级种群个体数;k为种群大小级数量。而当考虑未来的外部干扰时,Vpi还与年龄级数量(k)以及各龄级内的个体数(Sn)相关,可进一步将式(2)修正为:
(3)
(4)
式中,Vn、Vpi、Vpi′取正、负、零值时分别反映种群个体数量的增长、衰退和稳定的结构动态关系。其中,Vpi值越大,说明种群的增长趋势越大,而Vpi′值越大,说明种群抗干扰能力越大,种群稳定性越好。仅当P(种群对外界干扰所承担的风险概率)取值为最大时才会对种群动态Vpi′构成最大的影响。
1.3.3种群静态生命表和生存分析方法
静态生命表又称特定时间生命表,多用于对长寿命的木本植物种群的统计研究,主要包括以下参数:Ax是在x龄级内现有个体数;lx是在x龄级开始时标准化存活个体数;dx是从x到x+1龄级间隔期内标准化死亡数;qx是从x到x+1龄级间隔期间死亡率;Lx是从x到x+1龄级间隔期间还存活的个体数;Tx是从x龄级到超过x龄级的个体总数;ex是进入x龄级个体的生命期望或平均期望寿命;Kx为消失率(损失度)。它们之间的计算关系如下:
lx=(ax/a0)×1000
(5)
dx=lx-lx+1
(6)
qx=(dx/lx)×100%
(7)
Lx=(lx+lx+1)/2
(8)
(9)
ex=Tx/lx
(10)
Kx=lnlx-lnlx+1
(11)
由于静态生命表是用同一时期收集的种群所有个体编制而成,雅砻江冬麻豆种群为天然种群,分析中利用不同龄级结构在空间上的差异来研究特定时间段的种群动态,难免存在系统抽样误差产生与数学假设技术不符的现象,但仍能提供有用的生态学记录[15]。对于这种情况,在编制静态生命表时常采用匀滑技术对统计数据进行处理,本研究采用方程拟合方法进行匀滑处理[15- 17]。由于第Ⅵ龄级前的植株数存在明显异常,所以拟合采用第Ⅵ龄级及其之后的数据,以年龄级为自变量,以龄级对应的现存个体数Ax为因变量,经R3.3.3软件拟合得到方程:y=1473.506x-2.1263(R2=0.7517,P<0.001)。经此拟合方程匀滑修正后得到修正后的ax,然后据此编制出雅砻江冬麻豆种群的静态生命表。
为了检验种群存活状况是符合Deevey-Ⅱ型还是Deevey-Ⅲ型曲线,本文采用Hett和Loucks提出的数学模型对雅砻江冬麻豆种群存活曲线进行检验,即用指数方程Nx=N0e-bx和幂函数方程Nx=N0x-b分别描述Deevey-Ⅱ和Deevey-Ⅲ型存活曲线[18]。式中Nx、N0分别代表x龄级内存活数和种群形成初期的个体数,即Nx、N0为经匀滑后的个体存活数,b为死亡率。
为了更好地分析雅砻江冬麻豆种群动态,阐明其生存规律,采用种群生存率函数(S(i))、累计死亡率函数(F(i))、死亡率密度函数(f(ti))和危险率函数(λ(ti))进行分析[19-20]。计算公式如下:
S(i)=S1×S2×…Si
(12)
F(i)=1-S(i)
(13)
(14)
(15)
式中,Si为存活率,其计算公式为:Si=lx+1/lx,hi为龄级宽度。依据这4 个生存函数的估算值,绘制生存率曲线、累计死亡率曲线、死亡密度曲线与危险率曲线。
1.3.4种群数量动态的时间序列预测方法
本研究采用时间序列分析中的一次移动平均法[21]对雅砻江冬麻豆种群的年龄结构进行预测。
(16)
式中,n表示需要预测的时间(本研究为龄级时间),t为龄级,Xk为k龄级内的个体数量,Mt(1)表示经过未来n个龄级时间后t龄级的种群大小。本文对未来经过2、4、9、6、8、12、20和24龄级时间后的种群各龄级的个体数量进行预测。
2 结果与分析
2.1 雅砻江冬麻豆种群分布现状和结构特征

图1 雅砻江冬麻豆种群地径结构Fig.1 Size structure of S. bouffordiana population
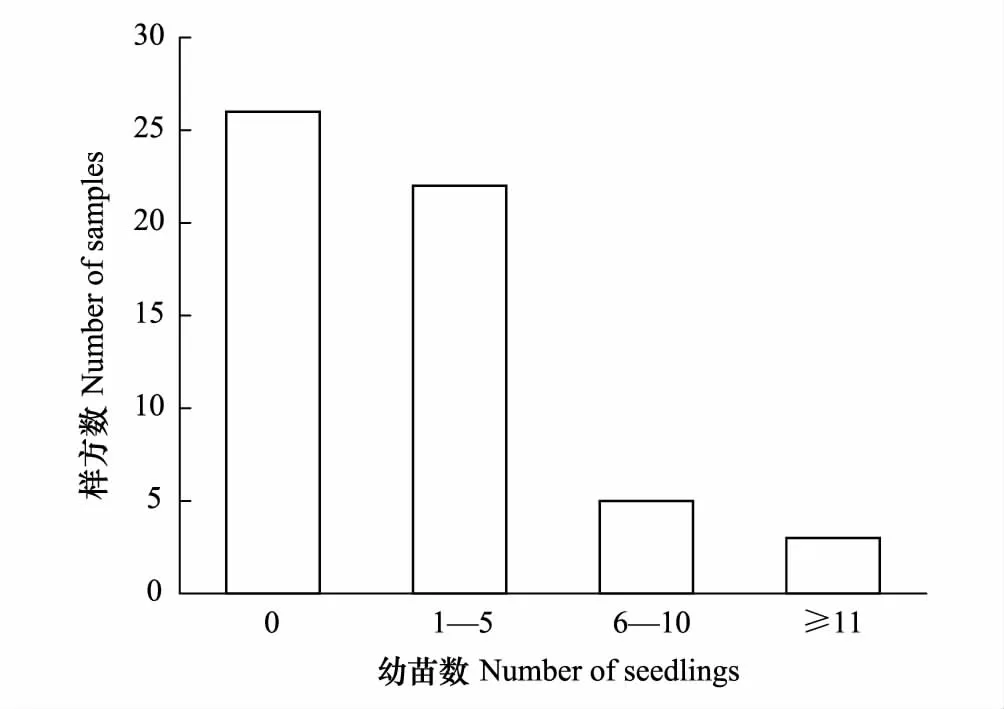
图2 调查样方内雅砻江冬麻豆幼苗数量分布Fig.2 Seedling distribution of S. bouffordiana in samples
雅砻江冬麻豆种群年龄结构呈纺锤形(图1)。种群植株个体主要集中在第Ⅲ—XIV龄级,即地径5—17 mm之间,占总植株数的80.71%。其中,地径8—15 mm的植株数占总植株数的56.85%,说明第Ⅵ—Ⅻ龄级的植株数量充足,而地径3—8 mm和>17 mm之间的植株数分别占总植株数的18.27%和14.72%,说明雅砻江冬麻豆种群缺乏低龄级和高龄级植株,且第Ⅰ—Ⅲ龄级的植株个体数占比之和不足9%,第XXIII—XXV级的植株个体数占比之和不足3%,表明种群有一定的更新能力,但更新后劲不足,甚为缺乏龄级两端处的植株,种群呈现出衰退状态。
根据调查样方幼苗数统计显示,约53.57%的样方中有雅砻江冬麻豆幼苗,其中以1—5株幼苗的居多,占总样方数的39.29%(图2),表明研究区内幼苗虽有分布,但其个体数量仍相当缺乏,种群更新能力低下。雅砻江冬麻豆种群整体呈现出中龄数量丰富,低龄数量缺乏,高龄数量严重不足的纺锤形结构,种群更新不良,种群年龄结构为衰退型。
由表1可知,雅砻江冬麻豆种群V6、V17、V19、V23均等于0,说明第Ⅵ—Ⅶ、XVII—XVIII、XIX—XX、XXIII—XXIV龄级间的个体数量没有变化,V4、V8、V10、V12、V14、V15、V16、V18、V20均大于0,说明第Ⅳ—Ⅴ、Ⅷ—Ⅸ、Ⅹ—Ⅺ、Ⅻ—XIII、XIV—XVII、XVIII—XIX、XX—XXI龄级间的个体数量呈现增长的结构动态关系,而剩余的龄级间均呈现出个体数量减少的结构动态关系,表明种群总体呈现出“衰退—增长—衰退—增长”的往复波动性结构动态关系。进一步分析其数量动态变化指数Vpi(不考虑外部干扰)和Vpi′(考虑随机干扰)的结果显示,Vpi=0.0799,Vpi′=0.0032,Vpi>Vpi′,虽然种群总体动态变化指数Vpi大于0,但仅为0.0799,且Vpi′值仅为0.0032,均趋向于0,表明种群波动大,种群稳定性差,抗干扰能力低。随机干扰风险极大值P极大=0.04,即种群结构对随机干扰的敏感性指数为0.04,说明雅砻江冬麻豆种群对外界随机干扰具有较高的敏感性,表明其对环境的适应力低,而且由于种群结构低龄个体更新不良,因此雅砻江冬麻豆种群应属于衰退型种群。
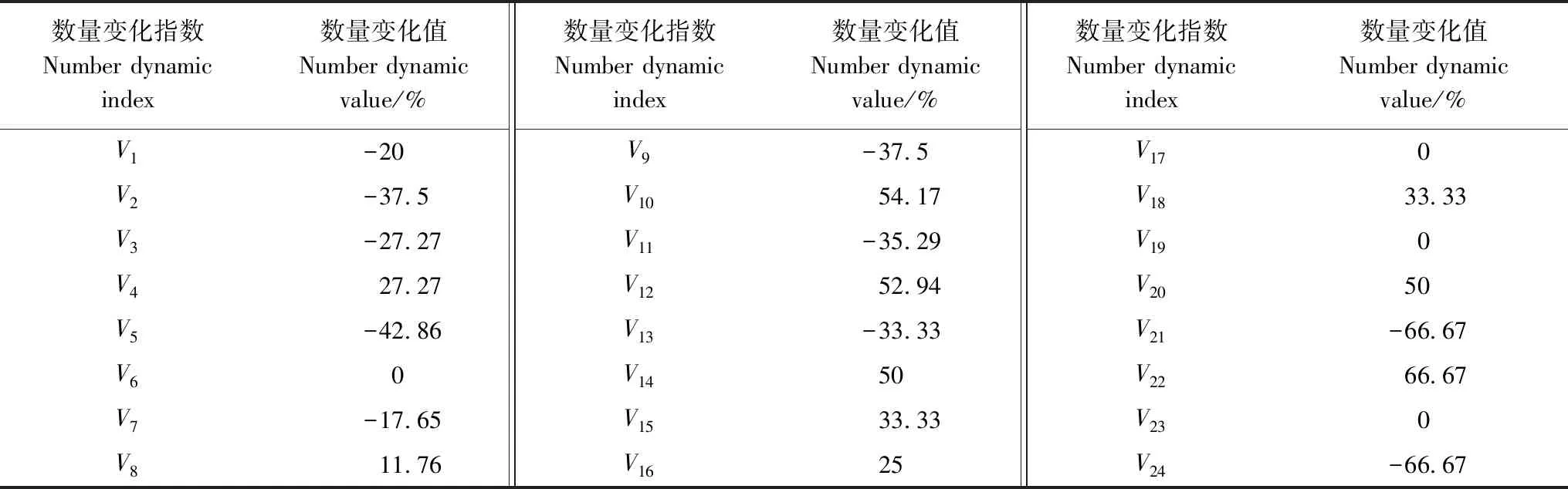
表1 雅砻江冬麻豆种群个体数量变化
Vn:种群从第n到n+1龄级的个体数量变化,V1为种群从第Ⅰ到Ⅱ龄级的个体数量变化,V2、……、V24以此类推 the individual number change of the population from age classnto age classn+1,V1means the individual number change of the population from age classⅠ to age class Ⅱ, theV2toV24are calculated in the same way
2.2 雅砻江冬麻豆种群静态生命表
由雅砻江冬麻豆种群静态生命表可知,随着年龄的增大,雅砻江冬麻豆种群存活数(lx)逐渐减小(表2)。植株个体期望寿命(ex)随着龄级的增加呈现出先增后减的状态,其反应了植株个体的生命期望寿命,其最小期望寿命在第XXV龄级处(ex=1),最大期望寿命在第XIV龄级处(ex=7.667),说明雅砻江冬麻豆种群在较高龄级阶段仍然具有较高的生命期望。种群死亡率(qx)与消失率(Kx)反映了种群数量随着龄级增加的动态变化,雅砻江冬麻豆种群死亡率与消失率在第Ⅻ龄级前均随龄级增加而呈增加的状态,但在第Ⅻ龄级及其后出现剧烈波动,表明种群处于极不稳定状态。因此,雅砻江冬麻豆种群整体呈现出极不稳定的状态,种群发展状况差。
经R3.3.3软件拟合,雅砻江冬麻豆种群存活曲线的拟合方程为:
Nx=186.3488e-0.21878x(R2=0.8424,P<0.001)
Nx=1418.521x-2.10069(R2=0.998,P<0.001)
模型检验结果表明,两种模型的拟合结果在雅砻江冬麻豆种群中均达到了极显著水平,由于幂函数模型的R2值比指数函数模型大,故认为雅砻江冬麻豆种群存活曲线更趋近于Deevey-Ⅲ型。依据Deevey的划分[22]:Ⅰ型呈凸曲线,多数个体都能活到该物种的生理年龄,但当达到一定生理年龄时则大量死亡;Ⅱ型呈直线,说明各年龄的死亡率相等;Ⅲ型是凹曲线,早期死亡率极高。因此雅砻江冬麻豆种群具有早期死亡率极高的特点。
以龄级为横坐标,以存活数的对数为纵坐标,根据雅砻江冬麻豆种群静态生命表绘制存活曲线(图3)。由图3可知,雅砻江冬麻豆种群存活曲线在第Ⅰ到第Ⅲ龄级几乎成直线下降,进入第Ⅳ龄级后,逐渐趋缓,且变化率在第XV、XVI、XIX龄级处和第XXII龄级及其之后均为0,同时自第XXI龄级之后存活数的对数值也均为0,表明雅砻江冬麻豆种群低龄级死亡率高,中龄级植株存活率相对较高,当种群进入第XV、XVI、XIX龄级及第XXI龄级之后大部分植株会较快死亡。在存活曲线方面,雅砻江冬麻豆种群与濒危物种鹅掌楸、矮牡丹、裂叶沙参、木根麦冬、长喙毛茛泽泻种群的研究结果一致[23],均为Deevey-Ⅲ型,高死亡率集中在低龄阶段,具有生殖能力的成年个体少。
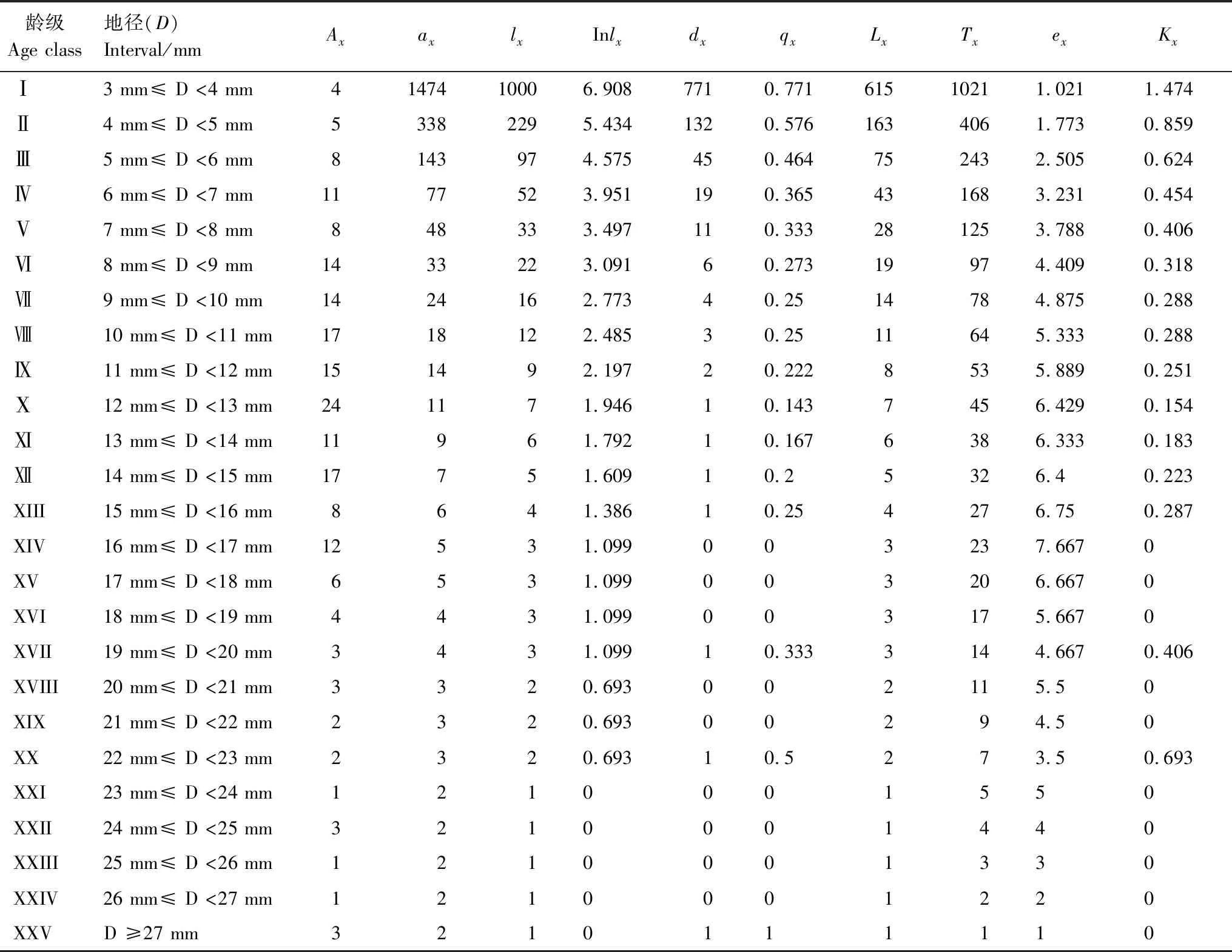
表2 雅砻江冬麻豆种群静态生命表
Ax:x龄级内现有个体数 individual number of age classx;ax:Ax匀滑后x龄内的存活个体数 the revised data ofAx;lx:x龄级开始时标准化存活个体数 the standardized number of surviving individuals of age classx;Inlx:lx的自然对数 the natural logarithm oflx;dx:从x到x+1龄级间隔期内标准化死亡数 the standardized number of death individuals from age classxto age classx+1;qx:从x到x+1龄级间隔期间死亡率 mortality from age classxto age classx+1;Lx:从x到x+1龄级间隔期间还存活个体数 the number of surviving individuals from age classxto age classx+1;Tx:从x龄级到超过x龄级的个体总数 the total individuals number of age classxand age classes elder thanx;ex:进入x龄级个体的生命期望或平均期望寿命 life expectancy or average life expectancy of individuals in the age classx;Kx:消失率 disappearance rate
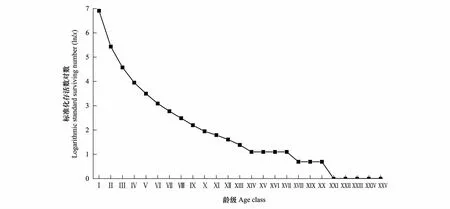
图3 雅砻江冬麻豆种群的存活曲线Fig.3 Survival curve of S. bouffordiana population
2.3 雅砻江冬麻豆种群生存
由图4可知,雅砻江冬麻豆种群的生存率随龄级增大表现出先急剧减小而后趋缓的状态,累计死亡率呈现出先快速递增而后趋缓的态势,二者互补,但种群生存率远小于累计死亡率,表明种群处于衰退状态。生存率曲线和累计死亡率曲线在前3个龄级呈现出急剧升降变化,在第Ⅴ龄级后逐渐趋缓,这可能和种群在低龄级的高死亡率有关,大量低龄级个体死亡,种群呈现衰退特征。在第Ⅵ龄级处,种群生存率为0.016,累计死亡率为0.984,种群已进入衰退期,表明雅砻江冬麻豆种群在生存早期已处于衰退状态。
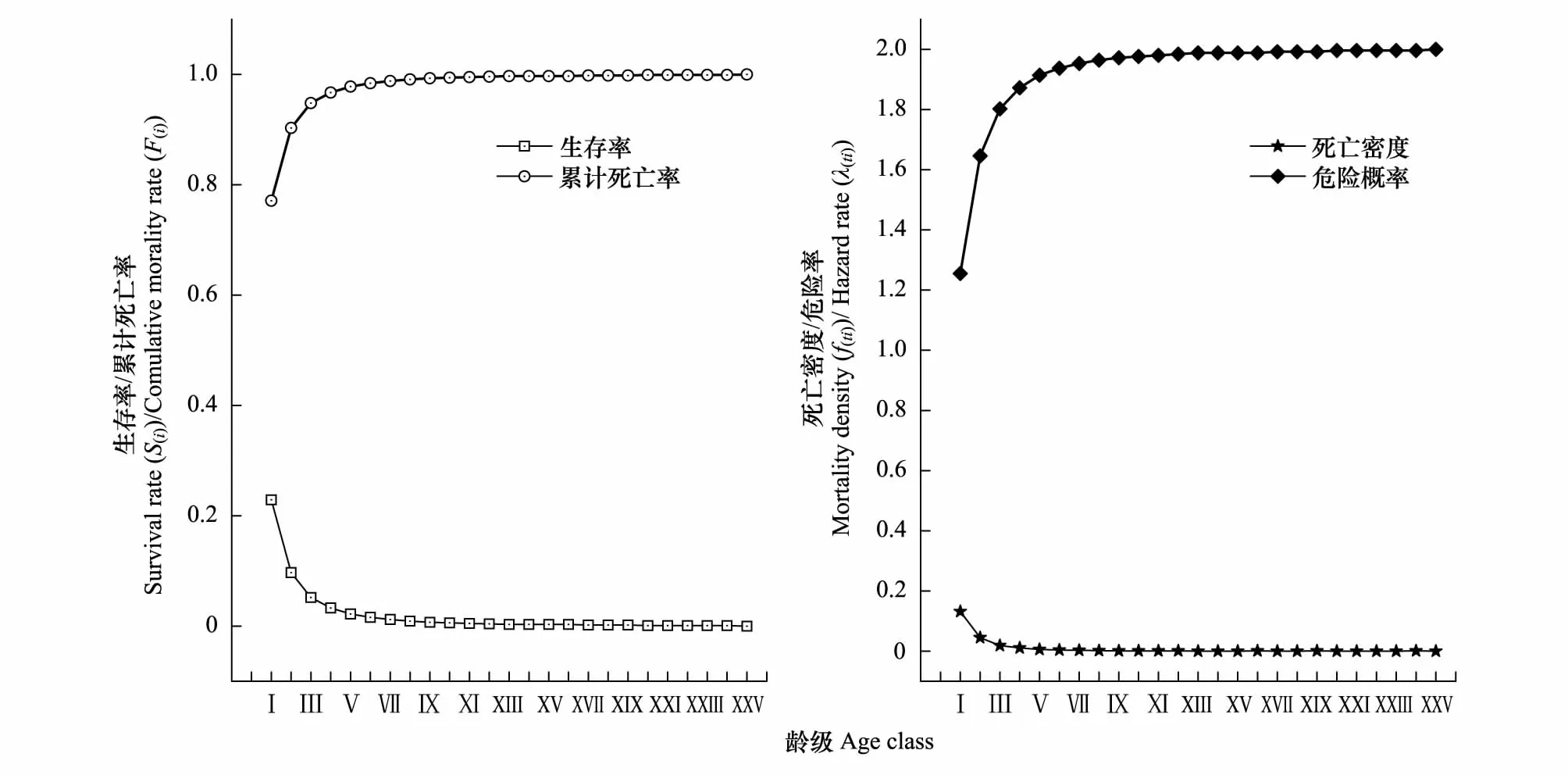
图4 雅砻江冬麻豆种群生存率、累计死亡率、死亡密度和危险率函数曲线Fig.4 Survival rate, cumulative mortality rate, mortality density and hazard rate functional curves of S. bouffordiana population
种群死亡密度曲线和危险概率曲线与其生存率和累计死亡率变化趋势相似,均在前3个龄级急剧变化,但种群危险概率曲线一直显著大于死亡密度曲线,在进入第Ⅳ龄级后均变化趋缓,其中危险概率曲线的整体变化更显著,死亡密度曲线在第XXV龄级达到最小值(0),危险概率曲线在第XXV龄级达到最大值(2)。由4 个生存函数曲线表明,雅砻江冬麻豆种群具有前期薄弱,中期、后期衰退显著的特点,种群生存力较低,处于衰退状态。
2.4 雅砻江冬麻豆种群数量的时间序列
雅砻江冬麻豆种群数量动态变化的时间序列预测是以其种群各龄级株数为原始数据,按照一次移动平均法预测出各龄级在未来2、4、9、6、8、12、20和24个龄级时间后的个体数。从表3分析可知,经历2、4、6个龄级时间,种群从第Ⅻ龄级开始之后均有不同程度的增加,经过8和12个龄级时间,从第Ⅻ龄级开始减少,而后趋于稳定,说明一旦过了一定的生理年龄,雅砻江冬麻豆种群就开始逐渐走向衰退。种群前期,在第Ⅵ龄级经过2、4和6个龄级时间后个体数分别下降了21.43%、28.57%和42.86%;种群前中期,在第Ⅺ龄级经过2、4、6和8个龄级时间后个体数下降了16.67%、25.00%、37.50%和41.67%;而种群中期,在第Ⅻ龄级经过12个龄级时间后个体数下降了29.41%。表明雅砻江冬麻豆种群严重缺乏低龄个体。综合其整个发展趋势来看,雅砻江冬麻豆种群正常更新难以维持,未来仍然会继续衰退。
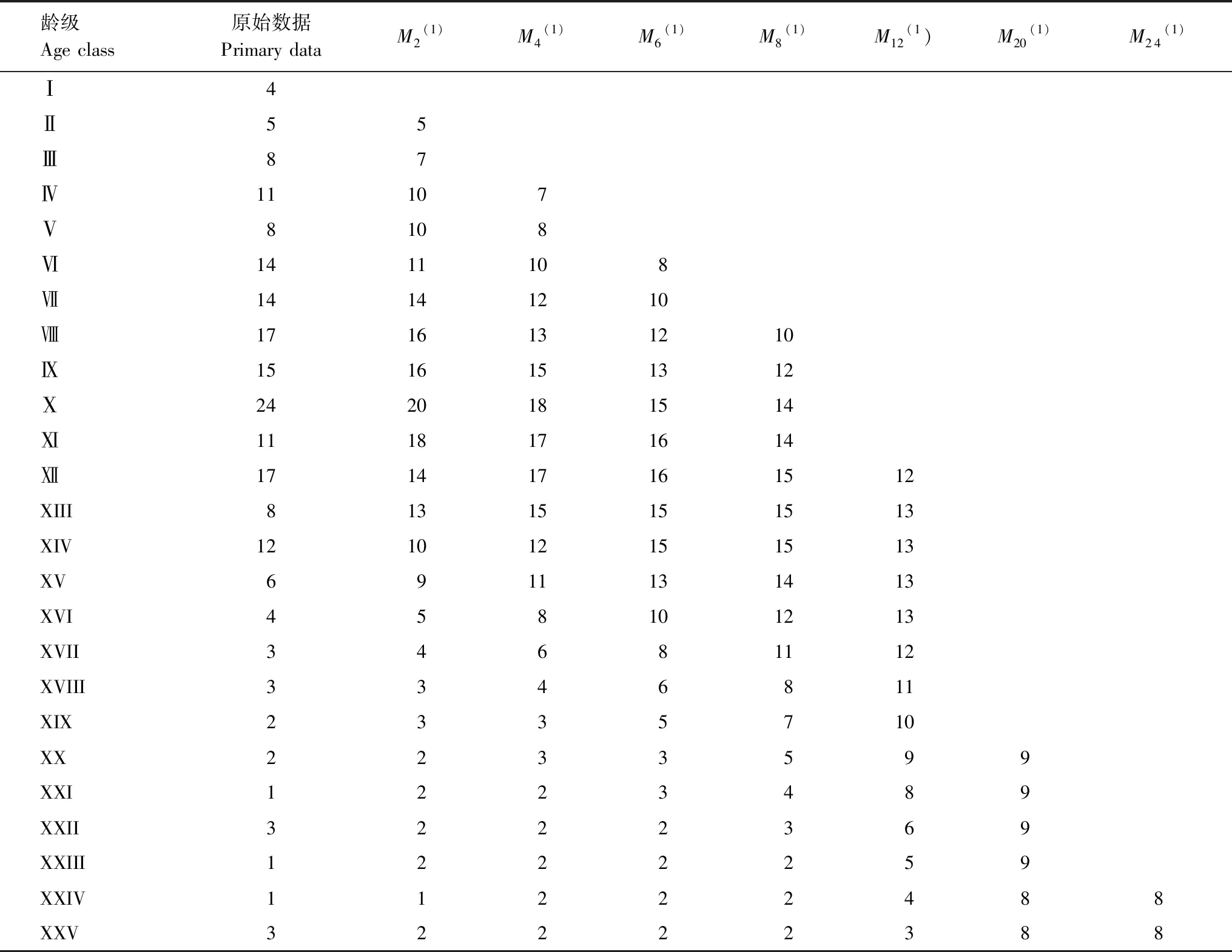
表3 雅砻江冬麻豆种群动态变化的时间序列分析
Mt(1):经过2、4、6、8、12、20和24龄级时间后t龄级的种群大小the population size of age class t after the time of 2, 4, 6, 8, 12, 20, 24 age class;(1):移动平均法中的一次移动平均法标识the mark of one-time moving average in moving average method
3 讨论
植物种群结构特征能够体现种群内部个体的发展过程,其径级结构能很好地反应种群动态变化[24]。生命表是判断植物种群发展趋势的重要指标之一,对植物种群的生命表和存活曲线的分析可以反映种群现实状况、展现植物种群与环境的竞争关系,对珍稀濒危植物的保护和利用具有重要意义[25]。本研究得出雅砻江冬麻豆种群年龄结构为纺锤形衰退型,动态指数显示种群波动大、稳定性差、抗干扰能力低,且其对生存环境适应力低、具有较高的敏感性。静态生命表显示雅砻江冬麻豆种群整体呈现出极不稳定的状态,种群发展状况差。其存活曲线为Deevey-Ⅲ型,早期死亡率极高。生存函数曲线表明雅砻江冬麻豆种群具有前期薄弱,中期、后期衰退显著的特点,种群生存力较低,处于衰退状态。时间序列分析显示雅砻江冬麻豆种群严重缺乏低龄个体,种群正常更新难以维持,未来仍然会继续衰退。因此,雅砻江冬麻豆种群为衰退种群,急需进一步弄清雅砻江冬麻豆种群的衰退原因和衰退程度,也需对雅砻江冬麻豆种群进行生物化学与分子生物学、生态学和地理学等方面的综合研究。
通过对极小种群梓叶槭(Acercatalpifolium)[26]、多脉铁木(Ostryamultinervis)[27]、广西青梅(Vaticaguangxiensis)[28]、海南风吹楠(Horsfieldiahainanensis)[29]、圆叶玉兰(Magnoliasinensis)[30]、馨香木兰(Magnoliaodoratissma)[31]、大树杜鹃(Rhododendronprotistumvar.giganteum)[32]、单性木兰(Kmeriaseptentrionalis)[33]、伯乐树(Bretschneiderasinensis)[34]和雅砻江冬麻豆种群的种群结构与动态特征比较发现(表4):(1)从龄级结构上看,梓叶槭、大树杜鹃、伯乐树与雅砻江冬麻豆种群均呈现出衰退型;(2)从存活曲线上看,多脉铁木、广西青梅、大树杜鹃、单性木兰与雅砻江冬麻豆种群均为Deevey-Ⅲ型,而梓叶槭、海南风吹楠种群为Deevey-Ⅱ型(除馨香木兰种群为Deevey-Ⅰ型外),这与张文辉等对十种濒危植物种群总结出来的存活曲线结论基本一致[23];(3)从种群动态指数上看,梓叶槭和多脉铁木为增长型,而雅砻江冬麻豆种群均为衰退型;(4)对种群关键威胁因素进行分析,表明种群均与种子问题或者幼苗问题有关,而外因与干扰、立地环境和竞争有关。所以,这些极小种群和雅砻江冬麻豆种群均表现出脆弱性高的共性。
表4 极小种群和雅砻江冬麻豆的种群结构与动态特征比较分析
Table 4 The comparison in population structures and dynamic characteristics between extremely small populations andS.bouffordianapopulation
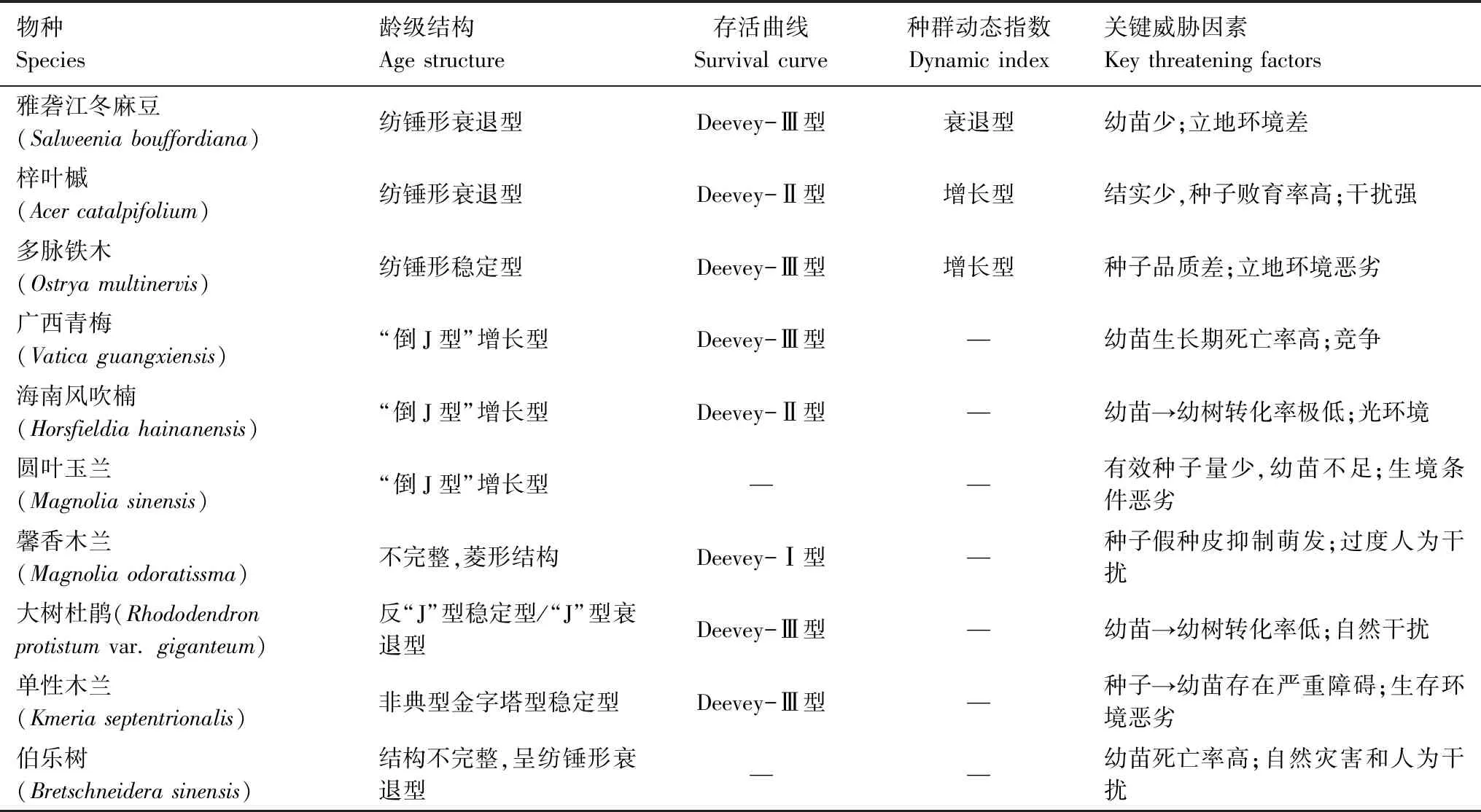
物种Species龄级结构Age structure存活曲线Survival curve种群动态指数Dynamic index关键威胁因素Key threatening factors雅砻江冬麻豆(Salweenia bouffordiana)纺锤形衰退型Deevey-Ⅲ型衰退型幼苗少;立地环境差梓叶槭(Acer catalpifolium)纺锤形衰退型Deevey-Ⅱ型增长型结实少,种子败育率高;干扰强多脉铁木(Ostrya multinervis)纺锤形稳定型Deevey-Ⅲ型增长型种子品质差;立地环境恶劣广西青梅(Vatica guangxiensis)“倒J型”增长型Deevey-Ⅲ型—幼苗生长期死亡率高;竞争海南风吹楠(Horsfieldia hainanensis)“倒J型”增长型Deevey-Ⅱ型—幼苗→幼树转化率极低;光环境圆叶玉兰(Magnolia sinensis)“倒J型”增长型——有效种子量少,幼苗不足;生境条件恶劣馨香木兰(Magnolia odoratissma)不完整,菱形结构Deevey-Ⅰ型—种子假种皮抑制萌发;过度人为干扰大树杜鹃(Rhododendron protistum var. giganteum)反“J”型稳定型/“J”型衰退型Deevey-Ⅲ型—幼苗→幼树转化率低;自然干扰单性木兰(Kmeria septentrionalis)非典型金字塔型稳定型Deevey-Ⅲ型—种子→幼苗存在严重障碍;生存环境恶劣伯乐树(Bretschneidera sinensis)结构不完整,呈纺锤形衰退型——幼苗死亡率高;自然灾害和人为干扰
根据种群结构与动态特征比较分析表明,雅砻江冬麻豆种群与极小种群存在多方面的相似,但其生物学地位却存在差距。确定濒危植物种群的生物学地位是植物保护工作的第一步[35],而雅砻江冬麻豆种群的生物学地位还未引起足够的重视。任何特有种都可能受到其有限分布的威胁[36],种群灭绝的概率与其种群大小呈负相关[35]。雅砻江冬麻豆属于小种群,为衰退型结构,种群更新后劲乏力,生存力较低,且已处于濒危状态,面临灭绝的可能。研究发现,人类活动是造成物种濒危的首要因素[35],而人类活动无处不在,随着人类活动的不断扩张,雅砻江冬麻豆种群的生存将面临着更为严峻的形势。因此,需要相关部门加快对雅砻江冬麻豆的研究步伐和尽快确定其生物学地位并对其进行定级,确保其得到有效保护。
就地保护是濒危植物解危的主要措施[23],其能使保护植物避免诸多外界干扰。本文建议对雅砻江冬麻豆种群进行就地保护,在雅砻江冬麻豆集中分布的区域设立野外保护小区,使其拥有一个修养生息的生存空间,每个保护小区必须针对所保护的物种特点制定相应的保护管理办法,加强保护点原生境的保护管理力度,加强巡护、监管,严禁盗伐、开荒、放牧、火烧、种植等改变原生境生态的行为,严格保护原生生境,防止人为干扰、自然灾害、兽类侵害的发生,保障原生种群的稳定和逐步繁衍壮大。
雅砻江冬麻豆种群年龄结构呈纺锤形衰退型结构、存活曲线为Deevey-Ⅲ型、动态指数显示为衰退型、对外界干扰具有较高的敏感性,属于亟需进行生物学定位的濒危物种。使物种出现敏感性的不稳定环境因素是造成物种陷入危机的主要原因,甚至可能会引起濒危物种的灭绝,所以保护雅砻江冬麻豆幼苗定居和存活以及为其营造稳定的生存环境是维持雅砻江冬麻豆种群存续和稳定的重要措施。
