外源亚精胺对渗透胁迫下烤烟幼苗生长及生理特性的影响
2019-10-15王艳芳申洪涛王丽君李丽华赵世民段卫东
李 冬,王艳芳,申洪涛,王丽君,李丽华,赵世民,刘 领*,段卫东*
1.河南科技大学农学院,河南省洛阳市开元大道263 号 471023
2.河南中烟工业有限责任公司技术中心,郑州经济技术开发区第3 大街9 号 450000
3.河南省烟草公司洛阳市公司技术中心,河南省洛阳市开元大道246 号 471023
近年来由于全球温室气体排放量的增加,全国范围内干旱危害和水资源匮乏现象日益加剧[1]。干旱已成为限制农作物产量的主要因素之一,所造成的损失相当于其他非生物胁迫造成的损失之和[2-3]。干旱胁迫通过影响植物光合作用、呼吸作用、渗透调节、蛋白质合成以及细胞膜的结构与功能等生理生化过程,进而影响到农作物和林木的生存、生长及产量[4-5]。烤烟是我国重要的经济作物,在整个生育期内对水分的变化极为敏感,尤其是在烟草苗期生长阶段,水分不足往往会导致烟苗移栽成活率低,严重影响到烟叶产量及品质的形成[6]。因此,研究烤烟在干旱胁迫下的适应性及保护机制,对优质烟叶生产具有重要意义。
多胺是生物体代谢活动中产生的具有强烈生理活性的含氮碱化合物,包括腐胺(Putrescine,Put)、精胺(Spermine,Spm)和亚精胺(Spermidine,Spd)等,在植物生长发育和生理调节中发挥着重要作用,其中Spd 由于其多聚阳离子特性,生理功能最强,可以直接作为胁迫保护物质和胁迫信号转导中的信号分子,协助逆境胁迫抗性机制的构建[7]。Spd 能通过影响核酸、蛋白质等生物大分子合成、清除活性氧自由基、稳定细胞膜结构等,参与到植物体内许多重要的代谢过程中,提高植物抗逆性[8-9]。近年来,越来越多的研究表明,Spd 能有效缓解多种非生物胁迫对植物体造成的损害,如干旱胁迫[10-12]、低温胁迫[13]、盐胁迫[14-15]、低氧胁迫[16]、淹水胁迫[17]、高温胁迫[18]和重金属胁迫[19]等。李州等[10]研究表明,外源Spd 能通过上调抗氧化酶相关基因和促进氨基酸代谢,减轻水分胁迫对白三叶的伤害;Yin 等[11]研究发现,干旱胁迫下外源Spd 和Spm 能有效提高樱桃体内多胺含量和抗氧化酶活性,增强光合作用能力;李丽杰等[12]研究发现,外源Spd 能显著促进干旱胁迫下玉米幼苗对光能的吸收利用,改善光合作用,减少玉米幼苗体内活性氧的产生量,减轻膜脂过氧化程度,增强根系活力,从而促进幼苗生长,缓解干旱胁迫对幼苗的损伤;Li 等[14]研究发现,外源Spd 可促进能量代谢、蛋白生物合成、抗氧化和解毒防御机制的蛋白高表达,从而提高黄瓜幼苗的耐盐性;李璟等[16]以黄瓜幼苗为材料,研究发现外源Spd 可有效提高低氧胁迫下根系中Spd 和Spm 含量,降低Put 含量,提高抗氧化酶活性以增强幼苗对低氧胁迫的耐受能力;僧珊珊等[17]研究表明,外源Spd 可通过有效改善淹水胁迫下玉米叶片光合功能,提高光系统Ⅱ电子传递速率和效率,提高根系的抗氧化酶活性,维持一定的有氧呼吸代谢,从而降低减产幅度;田婧等[18]研究发现,外源Spd 可显著降低高温胁迫下黄瓜幼苗叶片的膜脂过氧化程度,提高质膜和液泡膜质子泵活性,增强黄瓜幼苗的耐热性;王红霞等[19]研究发现,外源Spd 能显著提高铜胁迫下水鳖叶片的内源Spd 和Spm 含量,缓解矿质元素吸收平衡的紊乱,降低膜脂过氧化损伤。可见,目前有关外源Spd 对逆境胁迫下作物生长的研究已有大量报道。但在烟草上仅有韩锦峰等[20]研究发现,外源Spd 能有效提高烤烟根系活力,促进根系生长,同时多胺还可以明显增强烤烟光合作用能力,提高叶片烟碱含量,促进烟株生长发育,提高烟叶品质[21]。而在渗透胁迫条件下外源Spd 对烤烟幼苗生长、根系形态变化及光合荧光方面的研究则鲜见报道。鉴于此,采用营养液中加入聚乙二醇(PEG-6000)模拟渗透胁迫的方法,进行了喷施外源Spd 对渗透胁迫下烤烟幼苗生长、活性氧代谢、抗氧化酶活性、叶绿素含量及光合荧光特性的影响试验,旨在明确外源Spd缓解烤烟幼苗渗透胁迫的适宜浓度及作用机制,为烤烟的抗旱栽培及Spd 在烟草生产上的应用提供依据。
1 材料与方法
1.1 材料与试剂
供试烤烟品种为豫烟6 号,由河南省烟草公司洛阳市公司提供。外源亚精胺(Spd,纯度为99%)购于美国Sigma 公司,试验用PEG-6000 购于国药集团化学试剂有限公司。
1.2 试验设计
试验于2018 年5 月在洛阳市河南科技大学农学院旱作与节水生理生态重点实验室进行。试验种子用10%H2O2表面消毒10 min,蒸馏水冲洗后浸种8 h,然后在育苗盘中发芽,放入人工气候培养箱中培养,设置光照强度4 000 Lx,昼/夜温度(28/20±2)℃,光照周期14 h/10 h,当烟苗长至4片真叶时,选取生长均匀一致的幼苗,用蒸馏水洗净根部后移栽于盛有Hoagland 营养液[pH(5.7±0.2)]的不透明水培箱中(规格:41 cm×31 cm×14.5 cm),待烤烟幼苗培养至五叶一心时进行试验。渗透处理加入15%(W/V)的PEG-6000 模拟渗透胁迫(渗透势约为-0.3 MPa)。
试验共设置5 个处理:①对照(CK):Hoagland营养液+喷施蒸馏水;②T1:Hoagland 营养液+15%PEG+喷施蒸馏水;③T2:Hoagland 营养液+15%PEG+喷施0.2 mmol·L-1Spd;④T3:Hoagland营养液+15%PEG+喷施0.4 mmol·L-1Spd;⑤T4:Hoagland 营养液+15%PEG+喷 施0.6 mmol·L-1Spd。每个处理20 株烟苗,重复3 次;处理期间每2 d 更换1 次处理液,各处理每天喷施1 次蒸馏水或不同浓度的外源Spd,均匀喷施在叶片正反两面,至形成细雾状小液滴欲滴为止,连续喷施6 d,于处理6 d 后取烤烟幼苗自上而下第3 片叶进行相关指标的测定。
1.3 测定项目与方法
1.3.1 生物量与根系形态的测定
选取各处理烟苗5 株,用吸水纸将叶片和根系水分吸干,从根茎结合处剪断,分别称量地上部和地下部的鲜质量;采用Epson 1680 型扫描仪(日本Seiko Epson 公司)及WinRHIZ0 2008 根系分析软件(加拿大Regent 公司)对根系总长度、根系表面积、根系平均直径和根尖数进行分析。
1.3.2 活性氧代谢及抗氧化酶的测定
采用氮蓝四唑光化还原法测定超氧化物歧化酶(SOD)活性[22];采用愈创木酚法测定过氧化物酶(POD)活性[22];采用紫外吸收法测定过氧化氢酶(CAT)活性[22];采用硫代巴比妥酸比色法测定丙二醛(MDA)含量(质量分数)[22];参照王爱国等[23]的方法测定超氧阴离子(O2.-)产生速率。
1.3.3 叶绿素相对含量的测定
采用SPAD-502PLUS 叶绿素仪(日本Konica Minolta 公司),选取每片叶的叶尖、叶中和叶基3 个部位进行测量,取平均值,以SPAD(Soil and plant analyzer development)值表征叶绿素相对含量。
1.3.4 光合参数的测定
采用Li-6400 型光合作用仪(美国Li-COR 公司)于上午09:00—11:00 进行测量。测定时选择Li-6400-02B 红蓝光源,使用开放式气路,温度为25 ℃,空气相对湿度为50%~70%,设定有效光合辐射(PAR)为800 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1。测定烤烟幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci),并计算水分利用效率(Water use efficiency,WUE)即Pn/Tr,气孔限制值(Ls)即1-Ci/Co(Co为设定的CO2浓度)。
1.3.5 叶绿素荧光参数的测定
采用PAM-2100 便携式调制荧光仪(德国Walz公司)在光照条件下测定光下最大荧光(Fm')、稳态荧光(Fs)和光下最小荧光(Fo');然后将同部位烤烟幼苗叶片暗适应20 min,在黑暗条件下测定初始荧光(Fo)和最大荧光(Fm)。通过以上叶绿素荧光参数计算出光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ),测定时间同光合参数。
计算公式:
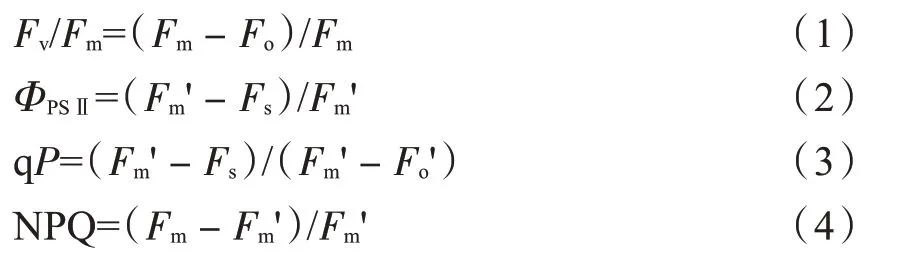
1.4 数据处理
采用Microsoft Excel 2016 和IBM SPSS 22.0 软件进行数据处理和分析,运用Duncan's 新复极差法进行处理间差异的显著性检验,显著水平设定为P<0.05,数据以平均值±标准误(Means±SE)表示,用Origin 9.0 软件进行绘图。
2 结果与分析
2.1 外源Spd 对烤烟幼苗鲜质量及根系形态的影响
由表1 可知,与对照相比,单纯渗透胁迫处理的烤烟幼苗的地上部鲜质量和地下部鲜质量降低30.67%和28.40%,总根长、总根表面积、根平均直径和根尖数分别降低33.98%、39.89%、33.33%和40.49%,说明渗透胁迫显著抑制了烤烟幼苗的生长发育。与单纯渗透胁迫相比,渗透胁迫条件下施加0.20、0.40 和0.60 mmol·L-1Spd 使烤烟幼苗的地上部鲜质量分别增加13.33%、27.12%和18.03%,地下部鲜质量分别增加13.79%、27.59%和15.52%,总根长分别增加20.88%、34.59%和28.26%,总根表面积分别增加15.27%、29.08%和21.90%,根平均直径分别增加18.75%、3.75%和10.94%,根尖数分别增加26.68%、86.52%和48.27%,差异均达到显著水平。说明外源Spd 处理能够缓解渗透胁迫对烤烟幼苗生长的抑制,增加生物量积累,促进根系生长发育,且以Spd 浓度为0.40 mmol·L-1时效果最为显著。
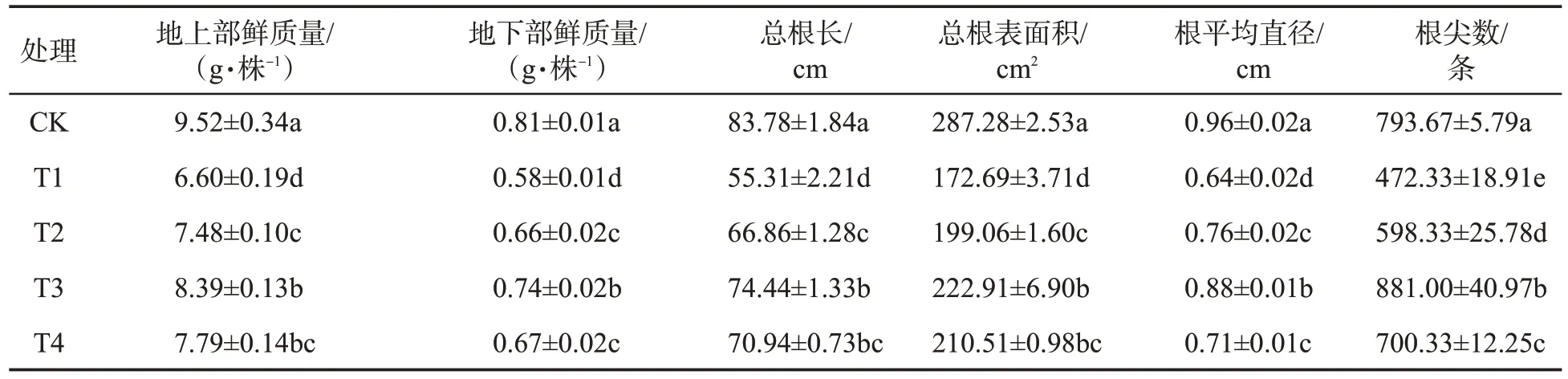
表1 外源Spd 对渗透胁迫下烤烟幼苗生长的影响①Tab.1 Effects of exogenous Spd on growth of flue-cured tobacco seedlings under osmotic stress
2.2 外源Spd 对烤烟幼苗超氧阴离子产生速率的影响
由图1 可知,与对照相比,单纯渗透胁迫处理的烤烟幼苗O2.-产生速率提高207.13%。随着外源Spd 浓度的增加,烤烟幼苗的O2.-产生速率呈先降低后升高的趋势。与单纯渗透胁迫相比,渗透胁迫条件下施加0.20、0.40 和0.60 mmol·L-1Spd 的O2.-产生速率分别降低17.02%、34.95%和22.51%,差异均达到显著水平。说明外源Spd 能显著降低烤烟幼苗O2.-产生速率,有效减少活性氧的产生,且以0.40 mmol·L-1的Spd 处理效果最佳。
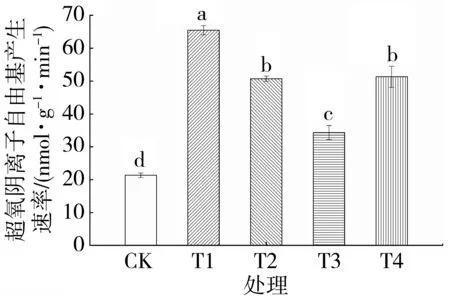
图1 外源Spd 对渗透胁迫下烤烟幼苗O2.-产生速率的影响Fig.1 Effects of exogenous Spd on O2.-generation rate of flue-cured tobacco seedlings under osmotic stress
2.3 外源Spd 对烤烟幼苗MDA 含量的影响
由图2 可知,与对照相比,单纯渗透胁迫处理的MDA 含量提高72.42%,说明渗透胁迫显著提高了烤烟幼苗的膜脂过氧化水平。随着外源Spd 浓度的增加,烤烟幼苗的MDA 含量呈先降低后升高的趋势。与单纯渗透胁迫相比,渗透胁迫条件下施加0.20、0.40 和0.60 mmol·L-1Spd 处理的MDA含量分别降低16.16%、24.19%和16.01%,差异均达到显著水平。说明外源Spd 能显著降低烤烟MDA含量,减轻膜脂过氧化对细胞的损伤,且以Spd 浓度为0.40 mmol·L-1时效果最为明显。

图2 外源Spd 对渗透胁迫下烤烟幼苗MDA 含量的影响Fig.2 Effects of exogenous Spd on MDA content of flue-cured tobacco seedlings under osmotic stress
2.4 外源Spd 对烤烟幼苗抗氧化酶活性的影响
由图3 可知,与对照相比,单纯渗透胁迫处理的POD、CAT 和SOD 活性增加41.63%、15.45%和12.96%。随着外源Spd 浓度的增加,烤烟幼苗的抗氧化酶活性呈先升高后降低的趋势。与单纯渗透胁迫相比,渗透胁迫条件下施加0.20、0.40 和0.60 mmol·L-1Spd 的POD 活性分别增加47.58%、83.94%和51.21%,CAT 活性分别增加28.51%、54.27%和23.33%,SOD 活性分别增加19.08%、35.08%和20.55%,差异均达到显著水平。说明外源Spd 能显著提高烤烟幼苗抗氧化酶活性,有助于维持活性氧代谢平衡,且以Spd 浓度为0.40 mmol·L-1时效果最佳。
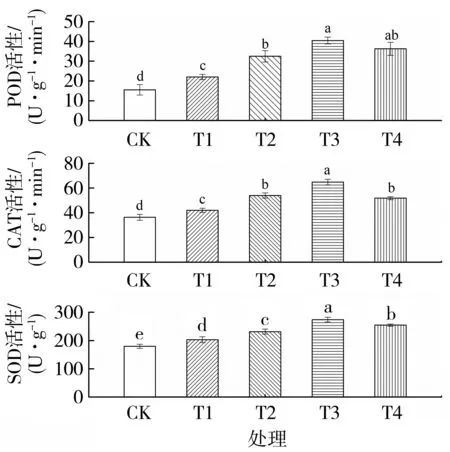
图3 外源Spd 对渗透胁迫下烤烟幼苗POD,CAT 和SOD活性的影响Fig.3 Effects of exogenous Spd on POD,CAT and SOD activities of flue-cured tobacco seedlings under osmotic stress
2.5 外源Spd 对烤烟幼苗叶绿素相对含量的影响
由图4 可知,与对照相比,单纯渗透胁迫处理的叶绿素相对含量降低44.81%,说明渗透胁迫显著抑制了烤烟幼苗的叶绿素合成。随着外源Spd浓度的增加,烤烟幼苗的叶绿素相对含量呈先升高后降低的趋势。与单纯渗透胁迫相比,渗透胁迫条件下施加0.20、0.40 和0.60 mmol·L-1Spd 处理的叶绿素相对含量分别增加33.79%、53.85%和32.42%,差异均达到显著水平。说明外源Spd 能显著增加烤烟叶绿素相对含量,促进光合色素的形成,且最适宜的Spd 浓度为0.40 mmol·L-1。
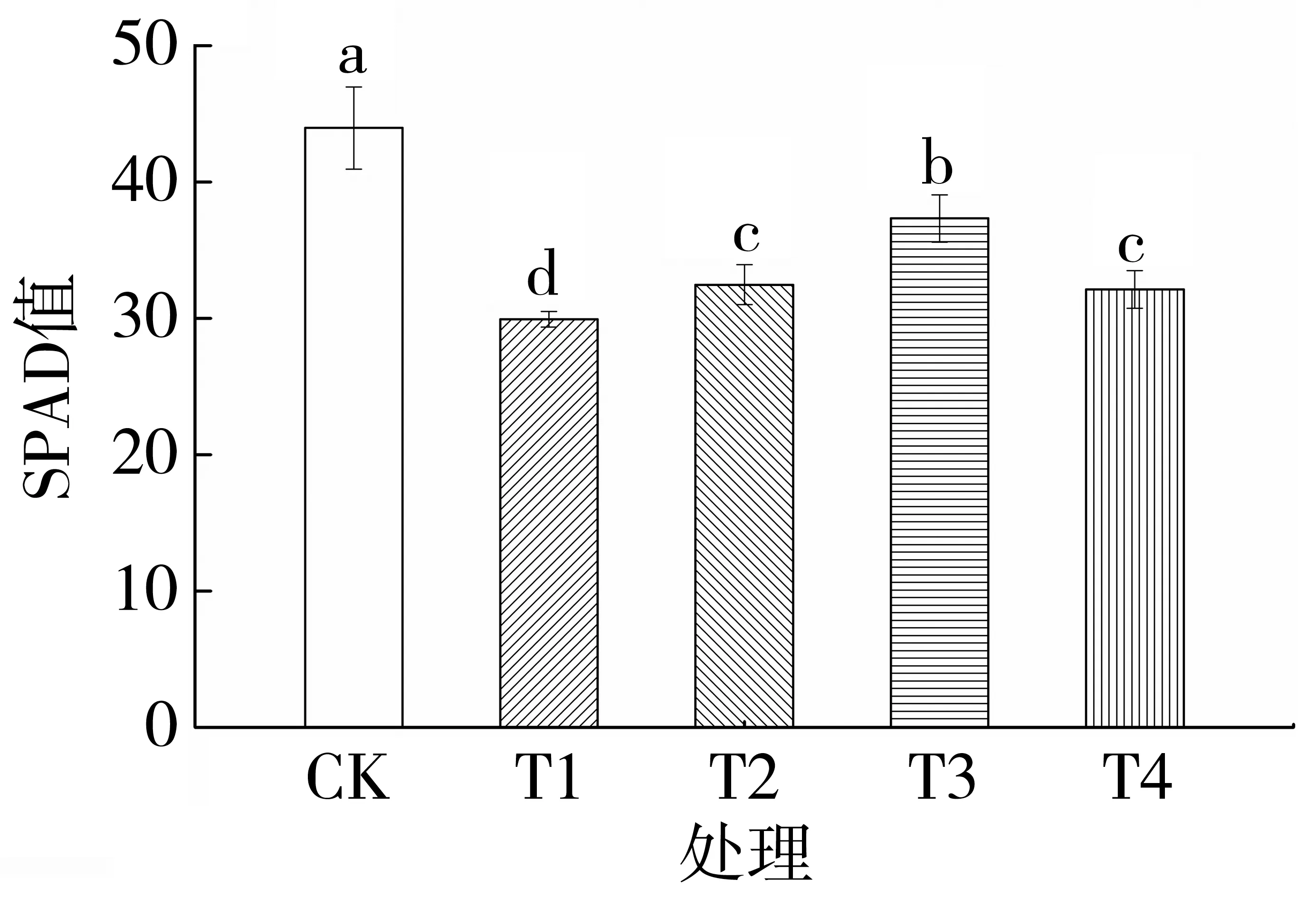
图4 外源Spd 对渗透胁迫下烤烟幼苗叶绿素相对含量的影响Fig.4 Effects of exogenous Spd on relative chlorophyll content in flue-cured tobacco seedlings under osmotic stress
2.6 外源Spd 对烤烟幼苗光合作用的影响
由图5 可知,渗透胁迫显著降低了烤烟幼苗光合作用。与对照相比,单纯渗透胁迫处理的Pn、Tr、Gs、WUE 和Ls均显著降低,Ci显著提高。随着外源Spd 浓度的增加,烤烟幼苗的Pn、Tr、Gs、WUE 和Ls表现出先升高后降低的趋势,Ci先降低后升高,差异均达到显著水平;其中施加0.40 mmol·L-1Spd处理的Pn、Tr、Gs、WUE 和Ls较单纯渗透胁迫处理分别提高178.95%、55.49%、30.01%、41.22%和34.64%,Ci降低了23.26%。说明外源Spd 能减缓渗透胁迫对烤烟幼苗的光合抑制作用,显著提高烤烟幼苗叶片的光合性能,从而增强烤烟幼苗的抗旱性。
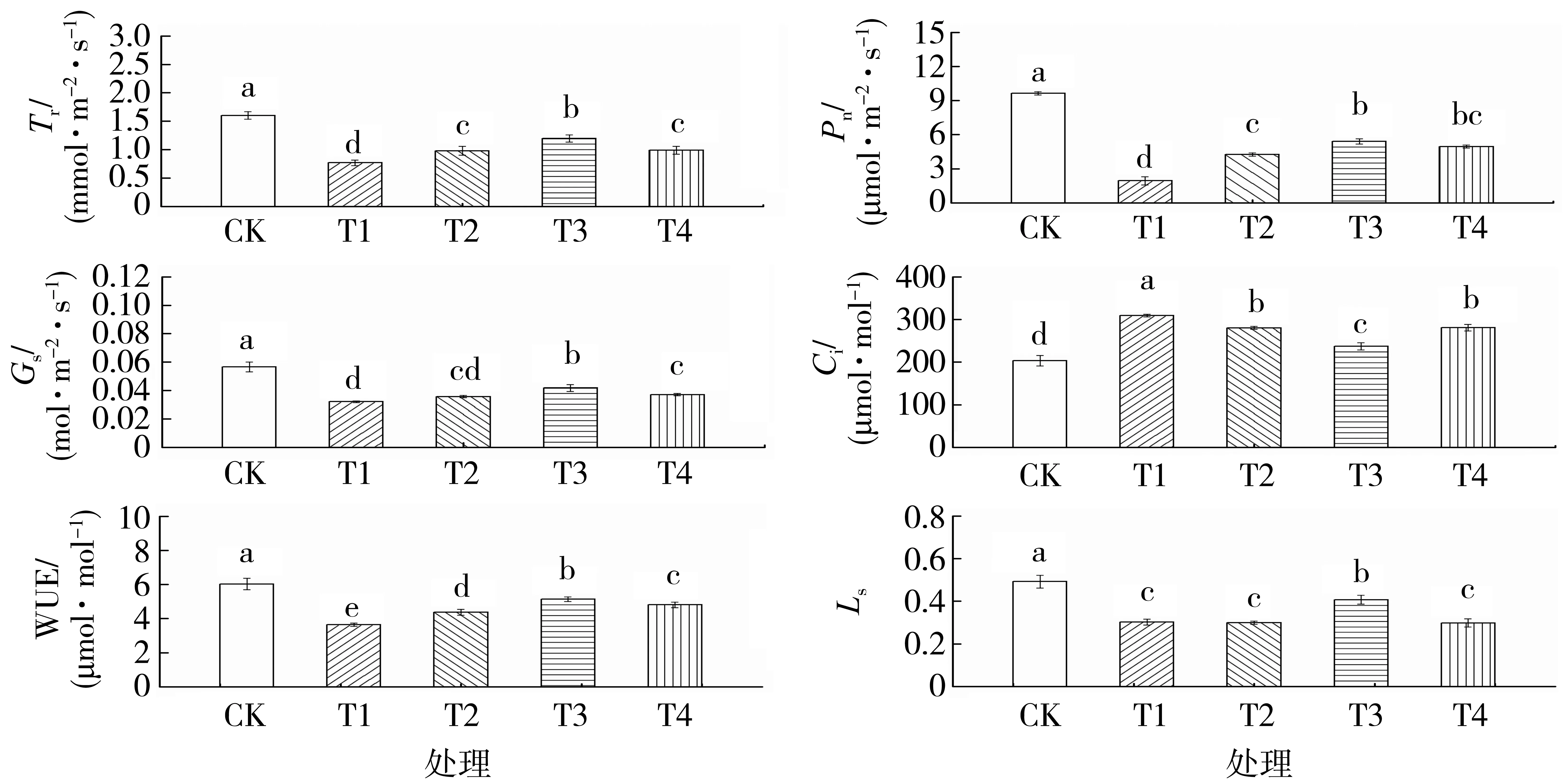
图5 外源Spd 对渗透胁迫下烤烟幼苗光合性能的影响Fig.5 Effects of exogenous Spd on photosynthetic characteristics of flue-cured tobacco seedling under osmotic stress
2.7 外源Spd 对烤烟幼苗叶绿素荧光参数的影响
由图6 可知,与对照相比,渗透胁迫下烤烟幼苗的PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)和光化学猝灭系数(qP)分别降低13.81%、29.44%和17.71%,非光化学猝灭系数NPQ提高75.24%。随着外源Spd 浓度的增加,烤烟幼苗的Fv/Fm、ΦPSⅡ、qP 表现为先升高后降低的趋势,NPQ 先降低后升高,差异均达到显著水平;其中施加0.40 mmol·L-1Spd 处理的Fv/Fm、ΦPSⅡ、qP 较单纯渗透胁迫处理分别提高11.27%、26.58%和16.15%,NPQ 显著降低30.93%。说明渗透胁迫使烤烟幼苗的PSⅡ潜在活性中心受损,而叶面喷施外源Spd能有效减缓渗透胁迫下烤烟幼苗的光抑制效应,提高光能的吸收和电子传递速率,且以0.40 mmol·L-1Spd 处理的效果最佳。
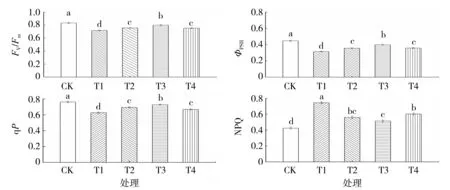
图6 外源Spd 对渗透胁迫下烤烟幼苗叶绿素荧光参数的影响Fig.6 Effects of exogenous Spd on chlorophyll fluorescence parameters of flue-cured tobacco seedling under osmotic stress
3 讨论
植物在干旱环境中会形成各种抗旱耐旱的形态结构,其中生物量的积累是植物对逆境响应的最直接体现,根系是植物对胁迫环境感应最早和最敏感的器官,植物的早期生长发育主要也是依赖于根系从周围环境中吸收养分和水分,以维持其正常的生命活动[24-25]。本研究结果表明,渗透胁迫下烤烟幼苗生物积累量及根系生长受到显著抑制,施加外源Spd 能显著缓解渗透胁迫对烤烟幼苗的伤害作用,提高其地上部与地下部生物量,促进总根长、总根表面积、根平均直径和根尖数的增加,且Spd 浓度以0.40 mmol·L-1效果最佳,说明在渗透胁迫下,适宜浓度的外源Spd 可有效增强植株根系对水分及营养物质的吸收、转运、积累和利用,促进烤烟幼苗的生长,以适应渗透胁迫,这与吴旭红等[26]在燕麦幼苗上的研究结果基本一致,但表现出的最佳Spd 浓度不同,可能是由于植物材料及胁迫伤害程度的差异所致,还需进一步的大田试验验证。
逆境胁迫下,植物细胞产生的自由基逐渐增加,其中以O2.-对植物造成的损伤最为严重。O2.-通过膜脂过氧化作用使细胞膜受到损害,导致生物体内活性氧大量积累[27],维系膜结构的功能键断裂,蛋白质变性失活[28-29]。本试验中,渗透胁迫下烤烟幼苗体内的O2.-产生速率和MDA 含量明显提高,打破了烤烟幼苗细胞内抗氧化代谢的平衡,产生了过多的活性氧,破坏了细胞膜结构;外施不同浓度Spd 均能显著降低O2.-产生速率和MDA 含量,这与李丽杰等[12]在玉米上的研究结果基本一致,说明外源Spd 具有清除活性氧的作用,这有助于降低干旱胁迫下植物幼苗细胞膜脂过氧化程度,原因可能在于Spd 是H+的载体,具有SOD 的部分功能,能通过歧化反应来直接或间接地清除植物组织中的活性氧自由基[30]。SOD、POD 和CAT是植物体内活性氧自由基清除系统的保护酶,共同作用能够调节细胞渗透势,缓解逆境胁迫对植物细胞造成的氧化损伤[31-32]。本研究结果表明,渗透胁迫下烤烟幼苗体内的SOD、POD 和CAT 活性均不同程度提高,表明渗透胁迫会激活植株体内抗氧化酶相关基因,使其过量表达,自我调节以适应渗透胁迫,施用不同浓度的外源Spd 均能显著提高烤烟幼苗体内的SOD、POD 和CAT 活性,且随外源Spd 浓度的提高呈先升高后降低的趋势,以Spd 浓度为0.40 mmol·L-1时效果最为显著,这与李州等[10]的研究结果一致,说明适宜浓度的外源Spd 能有效提高烤烟幼苗的抗氧化能力,这可能是由于多胺通过参与DNA 的复制、转录和翻译,促进功能蛋白的合成,控制生物体内自由基的产生,提高抗氧化酶相关基因的表达量,从而促进逆境胁迫下植物幼苗的生长[33-35]。
大量研究表明,干旱胁迫会导致植物叶片气孔关闭,叶绿素合成受阻,光能利用效率下降,叶片光合机构遭受破坏,光合作用减弱[36]。Spd 内部带电荷的氨基和亚氨基通过以非共价键形式与蛋白质、核酸及带电荷的磷脂等共同作用,维持叶片类囊体膜结构的稳定性,抑制叶绿素的降解,减轻光系统内部光化学反应中心受到的抑制[37]。在本试验中,渗透胁迫下烤烟幼苗的光合抑制,主要是由于叶绿素合成能力下降和非气孔限制因素造成的。喷施不同浓度外源Spd 均能显著提高烤烟幼苗的SPAD 值、Pn、Tr、Gs、WUE 和Ls,这与李丽杰等[12]的研究结果一致,其原因可能是外源Spd 通过影响叶绿素合成或降解相关酶的活性,保护光合电子传递系统,维持较高的碳同化能力,从而增强其光合作用能力,提高其抵抗渗透胁迫的能力,进而促进植株生长发育。
叶绿素荧光参数是研究植物光化学效率、光抑制和光破坏防御的有效工具,能准确地检测出完整植株在逆境条件下的光合作用真实行为,反映逆境胁迫下叶片对光能的吸收、传递、转换和耗散情况[38-39]。本研究结果表明,渗透胁迫下烤烟幼苗叶片的PSⅡ反应中心活性和开放程度明显降低,过剩光能大量积累,光合机构受到损伤,从而导致其光能利用能力降低。外源Spd 能缓解渗透胁迫对PSⅡ反应中心的损伤,降低热耗散程度,增加光合电子传递速率,使更多的光能用于光合作用,维持光合机构的稳定,提高光合效率,有效缓解渗透胁迫对烤烟幼苗生长的抑制作用,这与李泽等[40]在油桐幼苗上的研究结果一致。但有关外源Spd 在渗透胁迫下对作物生长及光合系统影响的分子机理,还有待进一步深入研究。
4 结论
在实验室人工模拟渗透胁迫条件下,水培试验结果表明:外源Spd 能有效缓解渗透胁迫对烤烟幼苗生长的抑制作用,以Spd 浓度0.40 mmol·L-1时效果最佳。外源Spd 可通过提高烤烟幼苗叶片抗氧化酶活性,降低O2.-产生速率和MDA 生成量,减轻膜脂过氧化损伤程度,提高叶绿素含量,增强光合作用能力,减轻光抑制程度,维持光合电子传递速率,促进植株根系的生长发育,从而提高烤烟幼苗的抗旱性。
