氮、磷、硫对铬超富集植物李氏禾吸收CrⅥ的影响
2019-09-23伍婵翠刘杰梁英
伍婵翠 刘杰 梁英


摘要:通过水培试验,以用25% Hoaglands营养液培养的植株作为对照,研究无机营养元素氮、磷、硫對铬超富集植物李氏禾吸收六价铬[Cr(Ⅵ)]的影响。结果表明,李氏禾根系对Cr(Ⅵ)的吸收符合系Michaelis-Menten方程,其米氏常数(Km)为91.84 μmol/L。与全素培养条件相比,在缺氮、缺硫条件下,李氏禾的根系吸收位点与Cr(Ⅵ)的亲和力增加,根系分泌物能增加土壤中难溶性铬的活性,从而有利于对Cr(Ⅵ)的吸收;而在缺磷培养条件下,根系分泌物不利于土壤中难溶性铬的溶出,缺磷条件对李氏禾吸收Cr(Ⅵ)存在非竞争性抑制作用。由研究结果可知,适当施用磷肥有利于提高李氏禾对Cr(Ⅵ)污染土壤的植物修复效率。
关键词:超富集植物;李氏禾;氮;磷;硫;植物修复效率
中图分类号: X173 文献标志码: A 文章编号:1002-1302(2019)14-0316-05
受人类活动的影响,土壤中的六价铬[Cr(Ⅵ)]污染已经成为亟待解决的环境问题之一。李氏禾(Leersia hexandra Swartz)是我国境内发现并报道的首例铬超富集植物[1],因其强大的吸收和富集Cr(Ⅵ)的能力,在Cr(Ⅵ)污染土壤的修复方面具有良好的应用前景。然而,重金属污染土壤的植物修复效率受到很多外界因素的影响,土壤中的非金属营养元素正是影响因素之一。此外,氮(N)、磷(P)、硫(S)对土壤重金属形态转化及植物吸收累积的影响也成为环境领域的研究热点之一。
已有一些研究者分析了不同营养元素对植物吸收重金属的影响。楼玉兰的研究表明,硝态氮的施用会促进OH-或HCO3-的释放,使根际pH值上升,从而降低土壤中重金属的活性,减少了植物对重金属的吸收[2]。郭光光等的研究表明,施用磷肥提高了蓖麻地下部对铜(Cu)的吸收和积累[3]。López-Bucio等研究发现,磷抑制了拟南芥对铬的吸收[4]。Shi等研究发现,在铜污染土壤中施加硫肥有利于水稻对铜的吸收累积[5]。Zhong等研究发现,在砷(As)污染土壤中施用硫肥可减少油菜籽粒中的砷含量[6]。Marchiol等利用高粱(Sorghum bicolor)、向日葵(Helianthus annus)在各种金属污染土壤中进行植物提取试验,结果表明,施肥的植物叶片中富集的As、Cu和锌(Zn)要高于不施肥的植物[7]。由此可见,不同的营养元素对植物吸收土壤中重金属的影响存在一定差异,会因营养元素、植物和重金属种类的不同而有不同的影响。目前,关于氮、磷、硫影响铬超富集植物吸收Cr(Ⅵ)的研究未见报道。因此,研究氮、磷、硫对铬超富集植物李氏禾吸收 Cr(Ⅵ) 的影响,对于提高Cr(Ⅵ)污染土壤的植物修复效率、进一步完善植物修复技术都有重要意义。
1 材料与方法
1.1 供试材料
试验于2016年3月至2017年1月在桂林电子科技大学花江校区生命与环境科学学院环境工程实验室中进行。
本试验所用李氏禾幼苗采自桂林市灵川县灵田乡花江边的无污染农田。选取生长一致的李氏禾幼苗,用去离子水轻柔洗净后,放置于人工气候箱(LRH-550-GSI,珠江牌)中,用25% Hoaglands(霍格兰氏)营养液进行水培培养,每3 d更换1次营养液。培养条件如下:光—暗周期为12 h—12 h,白天温度控制在25 ℃,夜晚温度设置为20 ℃,相对湿度控制为75%,光照度为7 000 lx。每天需要加入超纯水以补充因水分蒸发而减少的培养液体积,每3 d更换1次营养液,预培养 15 d。
供试土壤取自用含铬废水处理的模拟人工湿地试验系统。待土壤自然风干、用玛瑙研钵研磨并过100目尼龙筛后备用。参照HJ 491—2009《土壤 总铬的测定 火焰原子吸收分光光度法》[8],采用微波消解仪(ETHOS A,Milestone,意大利)对土样进行消解,用原子吸收分光光度计(AA-6300,日本岛津)采用火焰法测定消解液中的铬浓度(mg/L)。供试土壤的总铬含量为1.80 mg/g。
1.2 试验方法
1.2.1 N、P、S与Cr(Ⅵ)的竞争吸收试验 试验前,将植物分成7组:(1)对照(用25% Hoaglands溶液培养);(2)用NO3-完全缺失的25% Hoaglands溶液培养;(3)用含有 5 mmol/L NO3-的25% Hoaglands溶液培养;(4)用H2PO4-完全缺失的25% Hoaglands溶液培养;(5)用含有5 mmol/L H2PO4-的25% Hoaglands溶液培养;(6)用SO42-完全缺失的25% Hoaglands溶液培养;(7)用含有5 mmol/L SO42-的25% Hoaglands溶液培养。将培养介质的pH值均调至5.5。48 h后,将K2Cr2O7溶液加入各组植物中,使其Cr(Ⅵ)浓度分别为20、40、60、80、100 μmol/L。每个处理设3个重复,于处理后4 h收获植物。
将收获后的李氏禾根系先用去离子水冲洗3次,再置于超声波清洗仪中,用10 mmol/L乙二胺四乙酸(EDTA)溶液清洗10 min,以排除植物根系表面吸附的Cr6+离子对试验结果的影响[9],最后用超纯水清洗3次。轻柔地吸干李氏禾根部表面的水分,并用陶瓷刀将根部切下,先于105 ℃杀青 30 min,再于80 ℃烘至恒质量。用陶瓷剪将李氏禾根系样品剪至长度为1~2 mm,得到植物干样。
使用微波消解仪(ETHOS A,Milestone,意大利),采用美国环保局(United States Environmental Protection Agency,简称USEPA)的方法3050B[10]对处理好的植物干样进行微波消解。在每个批次的试验中,菠菜标准物质(GBW10015)也用同样的方法进行消解,作为质控,仅当检测浓度在标定值的95%~105%范围内时,分析结果方可接受。
1.2.2 根系分泌物对难溶性铬的活化试验 试验前,将植物分成4组:(1)对照(用25% Hoaglands溶液培养);(2)用NO3-完全缺失的25% Hoaglands溶液培养;(3)用H2PO4-完全缺失的25% Hoaglands溶液培养;(4)用SO42-完全缺失的25% Hoaglands溶液培养。在试验第10天,光照2 h后将李氏禾取出,先用蒸馏水冲洗根部3遍,在新制备的超纯水中加入几滴0.05%百里酚溶液[11],浸泡李氏禾根部5 min,再用超纯水清洗根部3次。将李氏禾放入300 mL 0.5 mmol/L CaCl2溶液中[12],每杯加入3滴0.05%百里酚溶液,放回培养箱中光照收集4 h。4 h后,将收集液用0.45 μm滤膜抽滤后于40 ℃、90 r/min旋转蒸发浓缩,在比色管中定容至25 mL。
称取0.200 g三氧化二铬[分析纯(AR)]、0.200 g铬酸铅(AR)、2.000 g土壤样品,分别倒入装有根系分泌物的各个比色管中。在室温下连续振荡(振速为150 r/min)3 h后取上清液。用0.45 μm针式过滤器过滤后待测。每组设3个平行样。
1.3 样品的检测与统计分析
用原子吸收分光光度计(AA-6300,日本岛津公司)采用火焰法对样品中的铬浓度进行测定。标准系列使用液采用铬标准溶液[GBW(E)080257]配制而成。试验数据以3个平行样品测定值的算术平均值±标准偏差表示,数据用单因素方差分析法(ANOVA)进行统计分析,用最小显著差数法(LSD)评价数据间的差异水平(P<0.05表示差异显著)。
2 结果与分析
2.1 NO3-与Cr(Ⅵ)的竞争吸收
在5 mmol/L NO3-和NO3-缺失的条件下,考察氮对李氏禾吸收Cr(Ⅵ)的影响。由图1可见,李氏禾对Cr(Ⅵ)的吸收速率随着Cr(Ⅵ)浓度的增加而提高。在本试验的Cr(Ⅵ)浓度范围内,缺氮培养条件下李氏禾的铬吸收速率均高于对照;在5 mmol/L NO3-的培养条件下,李氏禾的铬吸收速率均低于对照。当营养液中的Cr(Ⅵ)浓度为20 μmol/L时,缺氮培养的李氏禾对铬的吸收速率比对照增加了18.0%;而在 5 mmol/L NO3-培養条件下, 李氏禾对铬的吸收速率比对照减少了25.4%。说明NO3-抑制了李氏禾根系对Cr(Ⅵ)的吸收,而氮元素的缺失则明显促进了李氏禾根系对铬的吸收。
为了进一步研究NO3-与李氏禾根系吸收Cr(Ⅵ)之间的关系,将缺氮、对照和5 mmol/L NO3-培养条件下李氏禾根系对Cr(Ⅵ)的吸收速率用Michaelis-Menten方程进行拟合(图2),根据双倒数法求得Michaelis-Menten方程的各参数,列于表1。由图2、表1可以看出,在5 mmol/L NO3-的培养条件下,李氏禾根系对Cr(Ⅵ)的最大吸收速率(vmax)比对照降低了16.2%,但米氏常数(Km)较对照明显提高,说明在此条件下,NO3-与Cr(Ⅵ)竞争李氏禾根部相同的吸收位点。而在缺氮条件下,李氏禾根系对Cr(Ⅵ)吸收的米氏常数较对照略小,表明在此条件下,李氏禾根系吸收位点与Cr(Ⅵ)的亲和力略微提高。
2.2 H2PO4-与Cr(Ⅵ)的竞争吸收
为了研究磷与李氏禾吸收Cr(Ⅵ)之间的关系,本试验在5 mmol/L H2PO4-和H2PO4-缺失的条件下考察李氏禾根系中的铬含量。由图3可见,铬吸收率随Cr(Ⅵ)浓度增加而增加;在本试验范围内的任何Cr(Ⅵ)浓度处理中,5 mmol/L H2PO4-培养条件下的铬吸收速率均高于对照,且在缺磷培养条件下的铬吸收速率均低于对照。当营养液中Cr(Ⅵ)浓度为100 μmol/L时,5 mmol/L H2PO4-培养条件下的铬吸收速率比对照提高了23.9%,而缺磷培养条件下的铬吸收速率比对照减少了14.4%。由上述分析得出,H2PO4-有利于李氏禾对Cr(Ⅵ)的吸收。
将5 mmol/L H2PO4-、对照和缺磷培养条件下李氏禾根系对Cr(Ⅵ)的吸收用Michaelis-Menten方程进行拟合(图4)。由表2可以看出,用5 mmol/L H2PO4-培养的李氏禾根系对Cr(Ⅵ)的最大吸收速率(vmax)较对照提高了17.5%,但米氏常数(Km)较对照明显降低了17.0%,表明在此条件下,李氏禾根系吸收位点与Cr(Ⅵ)的亲和力增加。缺磷条件培养的李氏禾根系对Cr(Ⅵ)的最大吸收速率(vmax)较对照有所降低,其米氏常数(Km)与对照接近,表明缺磷对李氏禾吸收Cr(Ⅵ)存在非竞争性抑制作用。
2.3 SO42-与Cr(Ⅵ)的竞争吸收
在5 mmol/L SO42-和SO42-缺失的条件下,考察硫对李氏禾吸收Cr(Ⅵ)的影响。由图5可见,铬吸收素速率随着Cr(Ⅵ)浓度增加而提高。在本试验的Cr(Ⅵ)浓度范围内,缺硫条件培养的李氏禾对铬的吸收速率均高于对照,5 mmol/L SO42-条件下培养的李氏禾对铬的吸收速率均低于对照。当营养液中Cr(Ⅵ)浓度为20 μmol/L时,缺硫条件培养的李氏禾对Cr(Ⅵ)的吸收速率较对照提高了29.8%,而5 mmol/L SO42-条件下培养的李氏禾对铬的吸收速率比对照减少了18.8%。
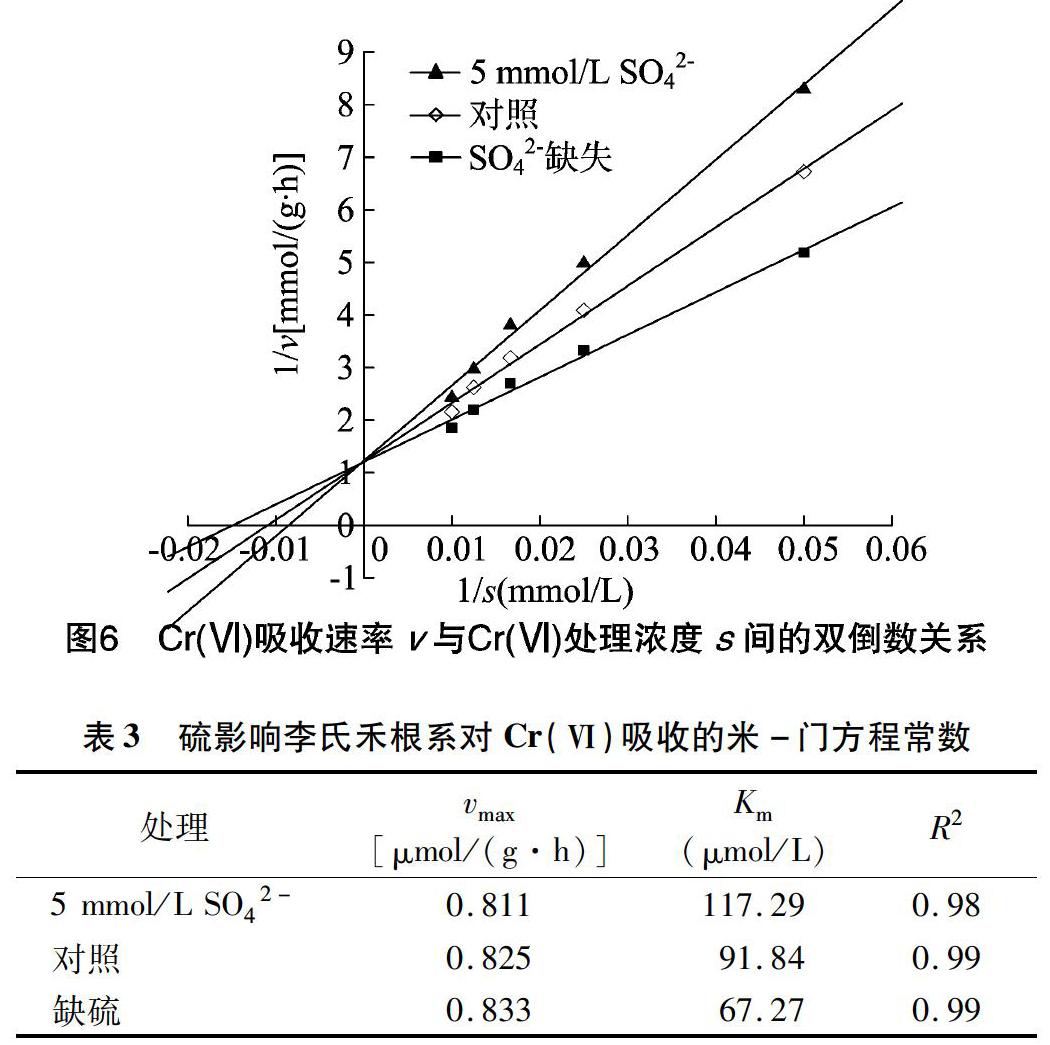
将缺硫、5 mmol/L SO42-和对照培养条件下李氏禾根系对Cr(Ⅵ)的吸收用Michaelis-Menten方程进行拟合(图6)。由表3可以看出,5 mmol/L SO42-条件培养的李氏禾对 Cr(Ⅵ) 的最大吸收速率(vmax)与对照相近,但米氏常数(Km)较对照提高了27.7%。这表明添加SO42-对李氏禾吸收 Cr(Ⅵ) 存在竞争性抑制。在缺硫培养条件下,李氏禾根系对Cr(Ⅵ)吸收的米氏常数较对照减少了26.8%,表明在此条件下, 李氏禾的根系吸收位点与Cr(Ⅵ)的亲和力增加, 即缺硫促进了李氏禾根细胞对Cr(Ⅵ)的吸收。
2.4 缺N/S/P培养对根系分泌物活化难溶性铬的影响
为了进一步验证李氏禾吸收Cr(Ⅵ)与非金属营养元素氮、硫和磷之间的竞争关系,取缺氮、缺硫和缺磷培养条件下的李氏禾根系分泌物,对难溶性铬进行活化试验,并与全素培养条件下的李氏禾根系分泌物对难溶性铬的活化效果进行对比。由图7可以看出,与全素培养的李氏禾根系分泌物相比,缺氮、缺硫条件培养的根系分泌物对Cr2O3的活化量虽略微增加,但并无显著差异(P<0.05),而缺磷条件培养的根系分泌物对Cr2O3的活化量显著降低,较全素条件培养的根系分泌物活化量减少了29.8%。缺氮和缺硫条件培养的根系分泌物对PbCrO4的活化量与全素条件培养的根系分泌物相比的显增加,而缺磷条件培养的根系分泌物对PbCrO4的活化量仅为全素条件培养的根系分泌物活化量的43.4%。在不同培养条件下,根系分泌物对于铬污染土壤中铬的活化量也存在显著差异。缺氮和缺硫条件培养的根系分泌物比全素条件培养的根系分泌物活化效果明显增加,而缺磷条件培养的根系分泌物校全素条件培养的根系分泌物的活化效果显著降低。
3 讨论与结论
土壤中存在的各种螯合剂及活性功能团可与重金属竞争吸附位点,从而影响重金属的生物有效性[13]。本试验结果表明,NO3-与Cr(Ⅵ)竞争李氏禾根部相同的吸收位点,缺氮有利于李氏禾对Cr(Ⅵ)的吸收。据报道,森林生态系统中的植物对Pb的吸收会因为NO3-的存在而受到明显抑制,从而减少Pb对植物的毒害作用[14]。还有研究表明,氮可以影响玉米根系对铬的吸收,且两者呈现负相关,即当氮含量增加时,玉米根系中的铬含量不断下降[15],这说明氮的存在可能会抑制玉米根系对铬的吸收。以上结论都与本试验的结果一致。当植物吸收NO3-时,会促使根系分泌OH-或HCO3-,使根际环境pH值升高,造成根际土壤碱化,从而降低土壤中的重金属溶解度,减少根系对重金属的吸收[2]。这有可能是NO3-抑制李氏禾吸收Cr(Ⅵ)的原因。
由于磷自身形态的复杂性,使得关于磷影响植物对土壤中重金属吸收的研究存在争议。关于磷影响植物对重金属吸收的报道也有不同结论。本试验结果表明,H2PO4-有利于李氏禾对Cr(Ⅵ)的吸收。這与一些学者的研究结论一致。如Singh的试验结果表明,在含镉土壤中加入过量的磷,可以促进油菜和燕麦对镉的吸收[16]。黄化刚等通过盆栽试验,发现高浓度的磷有助于东南景天对镉、锌的积累[17]。陈同斌等的研究发现,添加大量磷(含量为400 mg/kg以上)会使超富集植物蜈蚣草地上部、地下部的含砷量明显升高,磷过量对蜈蚣草富集砷具有明显的促进作用[18]。其原因有可能是磷的加入降低了土壤pH值,从而提高了土壤重金属的有效性[19]。此外,也有研究认为,磷对植物吸收重金属存在抑制作用。刘世亮等研究发现,磷抑制了油麦菜对镉的吸收[20],这与本研究结论相反。由此可见,添加磷对植物吸收重金属的影响与植物品种、重金属种类以及磷化合物组成的不同有密切关系。
硫不仅是植物必需的营养元素,也对植物重金属吸收调控具有重要影响。本试验结果表明,缺硫促进了李氏禾根细胞对Cr(Ⅵ)的吸收,添加SO42-对李氏禾吸收Cr(Ⅵ)存在竞争性抑制。这与前人的研究结论一致。如Hu等研究了SO42-对水稻吸收砷的影响,发现随着硫施加量的增加,水稻地上部分的砷累积量减少[21]。Fan等研究发现,施加过量硫能显著降低水稻糙米中镉的累积量[22]。Shewry等认为,硫酸根离子对大麦幼苗吸收Cr(Ⅵ)起到抑制作用[23]。其原因有可能是因为SO42-常被还原成S2-,并与土壤中其他金属离子如Hg2+、Cu2+、Cd2+等形成金属硫化物沉淀,从而降低金属的移动性[24]。
根分泌现象是植物根系为适应环境变化而作出的反应[25],植物的根分泌活动会受到一些因素(如本身基因型、生长环境等)的影响,其活动过程十分复杂。上述试验结果充分证明,李氏禾在不同营养条件(缺氮、缺硫或缺磷)下,其根系分泌物具有一定的特异性,其对特定物质的活化能力也有所不同。McGrath等通过根袋试验证明,超富集植物天蓝遏蓝菜(Thlaspi caerulescens)能够将土壤中难溶态的锌转化为可溶态锌,从而有利于植物对锌的吸收[26]。本试验表明,在缺氮条件下,李氏禾根系分泌物能有效活化土壤中的铬,使其活性提高,这与楼玉兰等认为的施用硝态氮会促使根系分泌OH-或HCO3-,从降低土壤重金属活性的研究结论[2]相符。在缺氮、缺硫培养条件下,李氏禾根系分泌物通过活化土壤中的难溶性铬,促进植物对铬的吸收,与本研究中缺氮、缺硫条件有利于李氏禾根系对Cr(Ⅵ)吸收的试验结果一致,与全素培养条件相比,缺磷培养条件下的根系分泌物不利于土壤中难溶性铬的溶出,因此不利于李氏禾根系对铬的吸收,与本研究中缺磷对李氏禾吸收Cr(Ⅵ)存在非竞争性抑制作用的结论一致,也与赵晶等认为的磷的加入降低了土壤pH值,提高了土壤重金属有效性的研究结论[19]相符。
综上所述,缺氮、缺硫有利于李氏禾对Cr(Ⅵ)的吸收,缺磷不利于李氏禾对Cr(Ⅵ)的吸收,适当添加H2PO4-有利于李氏禾对Cr(Ⅵ)的吸收。在后续工作中有必要进一步研究添加不同浓度的磷对李氏禾吸收Cr(Ⅵ)的影响,以确定H2PO4-的最佳施用量。试验结果对促进铬超富集植物李氏禾对污染土壤中Cr(Ⅵ)的吸收,从而进一步提高铬污染土壤的植物修复效率和完善植物修复技术有着重要的理论和现实意义。
参考文献:
[1]Zhang X H,Liu J,Huang H T,et al. Chromium accumulation by the hyperaccumulator plant Leersia hexandra Swartz[J]. Chemosphere,2007,67(6):1138-1143.
[2]楼玉兰,章永松,林咸永. 氮肥形态对污泥农用土壤中重金属活性及玉米对其吸收的影响[J]. 浙江大学学报(农业与生命科学版),2005,31(4):392-398.
[3]郭光光. 氮磷施用对蓖麻吸收,转运铜的影响及机制[D]. 武汉:华中农业大学,2017:52.
[4]López-Bucio J S,Dubrovsky J G,Raya-González J,et al. Arabidopsis thaliana mitogen-activated protein kinase 6 is involved in seed formation and modulation of primary and lateral root development[J]. Journal of Experimental Botany,2014,65(1):169-183.
[5]Shi J Y,Lin H R,Yuan X F,et al. Enhancement of copper availability and microbial community changes in rice rhizospheres affected by sulfur[J]. Molecules,2011,16(2):1409-1417.
[6]Zhong L,Hu C,Tan Q,et al. Effects of sulfur application on sulfur and arsenic absorption by rapeseed in arsenic-contaminated soil[J]. Plant Soil and Environment,2011,57(9):429-434.
[7]Marchiol L,Fellet G,Perosa D,et al. Removal of trace metals by Sorghum bicolor and Helianthus annus in a site polluted by industrial wastes:a field experience[J]. Plant Physiology and Biochemistry,2007,45(5):379-387.
[8]中华人民共和国国家环境保护标准. 土壤 总铬的测定 火焰原子吸收分光光度法:HJ 491—2009[S]. 北京:中国环境科学出版社,2009.
[9]Leita L,Contin M,Maggioni A. Distribution of cadmium and induced Cd-bingding proteins in roots,stems and leaves of Phaseolus vulgaris[J]. Plant Science,1991,77(2):139-147.
[10]de Oliveira L M,Ma L Q,Santos J A G,et al. Effects of arsenate,chromate,and sulfate on arsenic and chromium uptake and translocation by arsenic hyperaccumulator Pteris vittata L.[J]. Environmental Pollution,2014,184:187-192.
[11]张玉凤,冯 固,李晓林. 丛枝菌根真菌对三叶草根系分泌的有机酸组分和含量的影响[J]. 生态学报,2003,23(1):30-37.
[12]Pinto A P,Simoes I,Mota A M. Cadmium impact on root exudates of sorghum and maize plants:a speciation study[J]. Journal of Plant Nutrition,2008,31(10):1746-1755.
[13]Karlsson T,Skyllberg U. Complexation of zinc in organic soils EXAFS evidence for sulfur associations[J]. Environmental Science & Technology,2007,41(1):119-124.
[14]Burzyński M. The uptake and accumulation of phosphorous and nitrates and the activity of nitrate reductase in cucumber seedlings treated with PbCl2 or CdCl2[J]. Acta Societatis Botanicorum Poloniae,1988,57(3):349-359.
[15]李國军. 大量营养元素对玉米苗期铬吸收及生理特性的影响研究[D]. 太原:山西大学,2010:15.
[16]Singh B R. Cadmium and fluoride uptake by oats and rape from phosphorus fertilizers in two different soils[J]. Norwegian J Agric Sci,1990,4:239-250.
[17]黄化刚,李廷强,朱治强,等. 可溶性磷肥对重金属复合污染土壤东南景天提取锌/镉及其养分积累的影响[J]. 植物营养与肥料学报,2012,18(2):382-389.
[18]陈同斌,范稚莲,雷 梅,等. 磷对超富集植物蜈蚣草吸收砷的影响及其科学意义[J]. 科学通报,2002,47(15):1156-1159.
[19]赵 晶,冯文强,秦鱼生,等. 不同氮磷钾肥对土壤pH和镉有效性的影响[J]. 土壤学报,2010,47(5):953-961.
[20]刘世亮,刘忠珍,介晓磊,等. 施磷肥对Cd污染土壤中油麦菜生长及吸收重金属的影响[J]. 河南农业大学学报,2005,39(1):30-34.
[21]Hu Z Y,Zhu Y G,Li M,et al. Sulfur (S)-induced enhancement of iron plaque formation in the rhizosphere reduces arsenic accumulation in rice (Oryza sativa L.) seedlings[J]. Environmental Pollution,2007,147(2):387-393.
[22]Fan J L,Hu Z Y,Ziadi N,et al. Excessive sulfur supply reduces cadmium accumulation in brown rice (Oryza sativa L.)[J]. Environmental Pollution,2010,158(2):409-415.
[23]Shewry P R,Peterson P J. The uptake and transport of chromium by barley seedlings (Hordeum vulgare L.)[J]. Journal of Experimental Botany,1974,25(87):785-797.
[24]孙丽娟,段德超,彭 程,等. 硫对土壤重金属形态转化及植物有效性的影响研究进展[J]. 应用生态学报,2014,25(7):2141-2148.
[25]涂書新,孙锦荷,郭智芬,等. 植物根系分泌物与根际营养关系评述[J]. 土壤与环境,2000,9(1):64-67.
[26]McGrath S P,Shen Z G,Zhao F J. Heavy metal uptake and chemical changes in the rhizosphere of Thlaspi caerulescens and Thlaspi ochraleucm grown in contaminated soils[J]. Plant and Soil,1997,188(1):153-159.
