小麦转录因子TaWRKY26的克隆及其在抗、感白粉病和近等基因系中的差异表达
2019-05-31孙依帆牛欢冯静云黄建国张露露张超群范宝莉刘晓颖王振英
孙依帆 牛欢 冯静云 黄建国 张露露 张超群 范宝莉 刘晓颖 王振英


摘 要:WRKY转录因子家族在植物抗逆反应中发挥着重要的作用。本研究以抗白粉病小麦Brock为材料,从中克隆到一个WRKY基因,该基因全长1 485 bp,预测ORF长1 392 bp,编码463个氨基酸,分子量为49.8 kDa。经NCBI/Blastx比对发现,该基因与粗山羊草中WRKY26基因(序列号XP_020151406)所编码的转录因子同源性高达98%,故将其命名为TaWRKY26,并将扩增得到的序列全长上传至GenBank进行登记,序列号为MK358190。利用实时荧光定量PCR技术,对白粉菌胁迫后TaWRKY26在不同抗性小麦品种(抗病小麦Brock,感病小麦京411,抗病近等基因系Brock/京4117)中不同时间(0,2,4,8,12,24,48,72,120,168 h)的表达量进行分析,结果发现,在受到白粉菌侵染后,3个品种小麦的TaWRKY26表达量除12和72 hpi外,各时间段均表现为抗病小麦Brock>近等基因系Brock/京4117>感病小麦京411,抗白粉病小麦近等基因系Brock/京4117(抗病)中TaWRKY26表达趋势更偏向抗病亲本Brock。综合推测TaWRKY26在小麦对白粉菌侵染的应答反应过程中发挥了积极作用。
关键词:TaWRKY26基因;小麦;白粉病;实时荧光定量PCR技术
中图分类号: S512.1 文献标识码: A DOI 编码:10.3969/j.issn.1006-6500.2019.04.002
Cloning of Wheat TaWRKY26 Gene and Its Function Analysis in Powdery Mildew Resistance
SUN Yifan, NIU Huan, FENG Jingyun, HUANG Jianguo, ZHANG Lulu, ZHANG Chaoqun, FAN Baoli, LIU Xiaoying, WANG Zhenying
(College of Life Science, Tianjin Normal University, Tianjin 300387, China)
Abstract: The WRKY family plays an important role in the stress tolerance mechanism of plants. In this experiment, a gene of WRKY was cloned from the powdery mildew disease resistant wheat Brock. The gene contained a 1 392 bp ORF, and 463 amino acids with a predicted molecular weight of 49.8 kDa. The NCBI/Blastx alignment showed that this gene had 98% homology to the transcription factor encoded by the WRKY26 gene(SEQ ID: XP_020151406) in Aegilops tauschii, so we named it TaWRKY26. The full length of the amplified sequence was uploaded to GenBank for registration, and the accession number was MK358190. The expression patterns of TaWRKY26 in different resistant wheat[disease resistant wheat Brock, susceptible wheat variety Jing411, and Near-isogenic Lines Brock/Jing 4117(NILs)] after infected by powdery mildew were studied through real-time fluorescent quantitative PCR, the expression quantity at different time (0, 2, 4, 8, 12, 24, 48, 72, 120, 168 h) was analyzed. After infected by powdery mildew, the expression quantity of TaWRKY26 at 0, 2, 4, 8, 24, 48, 120, 168 h were all shown as follows: Brock>NILs >Jing411,and the expressive trend in NILs was more closer to disease resistant wheat Brock, indicating that TaWRKY26 participated in the interaction of wheat and powdery mildew.
Key words: TaWRKY26 gene; wheat; powdery mildew; RT-qPCR
小麥作为当今世界重要粮食作物之一,保证其产量对社会经济发展有着重要的意义。小麦产量容易受到各种病原体侵害的影响,其中,白粉病是危害最大的病害之一,白粉病流行会造成小麦减产30%[1]。
植物在长期进化过程中,形成了各种抵抗自然环境不利因素的抗性机制,主要分为分子模式触发的免疫反应(Pattern-triggered immunity,PTI)和效应子触发的免疫反应(effector-triggered immunity,ETI)[2-3]。植物受到病原菌侵染时会诱发PTI反应,使植物发生一系列的变化,如Ca2+外流、激活下游信号通路和丝裂原活化蛋白激酶(Mitogen-activated proteinkinase,MAPK)级联反应等[4-5]。
在植物MAPK级联反应中有一类植物所特有的转录因子——WRKY家族[6-7]。WRKY家族已在很多的植物、农作物中被分离鉴定出来,对其功能进行研究后发现WRKY基因在植物的生长发育和抗逆境胁迫过程中均发挥着重要的作用。樟树中发现的WRKY基因参与单萜类化合物芳樟醇的合成[8];在栗和狗尾巴草中发现WRKY基因对非生物胁迫和激素信号有较强的响应[9];香蕉中MaWRKY26基因参与香蕉对冷胁迫的响应[10];棉花WRKY 基因对盐、甘露醇和ABA胁迫均有响应[11]。WRKY家族不仅在抗非生物胁迫中起到重要作用,在植物抗病原菌的过程中,也起到了至关重要的作用。拟南芥中的AtWRKY33通过直接抑制ABA的表达水平来提高对灰霉病菌的抵抗作用[12];葡萄中的VvWRKY1过表达能够显著地提高对霜霉病的抗性[13];煙草中的WRKY基因沉默后植株对灰霉病的易感性增加[14]。
王振英教授实验室对白粉菌胁迫栽培小麦Brock的早期应答基因转录组进行了分析,发现多个抗病通路的基因出现表达差异,其中包括MAPK级联反应中的基因片段F5;通过Blastx比对发现F5与粗山羊草中WRKY26基因(序列号XP_020151406)所编码的转录因子的同源性高达98%,与普通小麦中的TaWRKY2同源性也高达95%(序列号ACD80357)。目前,对植物WRKY26和TaWRKY2基因的研究多集中在植物抗逆和生长发育方面,尚未见到关于小麦WRKY26基因在抗白粉病方面的研究报道。
本课题组为了分析F5基因与抗白粉病的相关性,根据相关序列设计引物对其全长进行了克隆,并通过实时荧光定量PCR技术对不同抗性小麦在受白粉菌胁迫的条件下该基因的表达模式进行分析,希望能够阐述WRKY26基因的功能,为其在小麦分子育种中的应用提供理论依据。
1 材料和方法
1.1 试验材料
白粉病抗性品种小麦Brock,由中国农业大学杨作民教授惠赠。
白粉病感性品种小麦京411,由王振英教授实验室保存。
白粉病抗性杂交品种近等位基因系Brock×京4117(Near-Isogenic Lines,NILs),是王振英教授实验室早期以抗白粉病栽培小麦Brock为抗病基因供体,以感病品种京411为轮回亲本,通过杂交获得F1,然后再以京411进行连续回交,每一代都在白粉菌选择压力下,选取抗病性状明显的单株,持续回交、选择6代后,再自交一代获得的抗病近等基因系。Brock×京4117在农艺性状上偏轮回亲本京411,抗白粉病性状偏供体亲本Brock。
小麦白粉菌15号生理小种,由中国农业科学院植物保护研究所提供,用于对小麦材料进行胁迫。
1.2 TaWRKY26基因克隆
利用Promega公司的EastepR Super Total RNA Extraction Kit提取Brock小麦的总RNA,利用琼脂糖凝胶电泳和Nanodrop1000检测RNA的质量以及浓度。然后以Oligo (d T)18为引物,利用Promega公司的M-MLV Reverse Transcriptase反转录成cDNA。根据Blastx比对结果设计WRKY基因引物(表1),进行PCR扩增。扩增体系为:5×PSGXL Buffer 5 μL,dNTP Mix 2 μL,WRKY-F 0.5 μL,WRKY-R 0.5 μL,cDNA1 μL,PrimeSTARR Max DNA Polymerase 0.5 μL,灭菌水15.5 μL。PCR反应程序为:94 ℃预变性5 min;98 ℃变性10 s,58 ℃退火15 s,68 ℃延伸2 min,反应进行35个循环;68 ℃终延伸7 min。将目标条带进行切胶回收,加A尾,连接到pGEM-T easy Vector上。将重组载体转化到大肠杆菌,进行蓝白斑筛选,选取阳性克隆送公司测序。
1.3 序列分析
将测序结果提交 NCBI GenBank进行同源性分析。用APE软件进行基因序列分析和拼接,同DNAMAN进行蛋白质序列比对分析并构建进化树。
1.4 白粉菌胁迫下TaWRKY26表达模式分析
为了探究目标基因与小麦抗白粉病特性之间的关系,采用实时荧光定量PCR技术对白粉菌胁迫下小麦中的目标基因进行表达模式分析。在相同且适宜的条件下培养Brock、京411和NILs小麦,待材料第一片叶完全打开时,以抖拂法将白粉菌15号生理小种的分生孢子均匀的接种在小麦叶片上,接种0,2,4,8,12,24,48,72,120,168 h后,取相同部位的叶片,用1.2中所述方法提取叶片的总RNA并进行反转录,将所得cDNA用于表达模式分析。
根据所扩增TaWRKY26片段设计特异的实时荧光定量PCR引物qWRKY(表1),所扩增片段长127 bp,用Promega公司的EastepRqPCR Master Mix在Applied Biosystems 7500 Real-Time PCR Systems上进行基因扩增、荧光信号检测和溶解曲线分析(此步骤为仪器自动进行)。小麦Actin基因作为内参基因,引物序列详见表1。所用程序为:50 ℃预变性2 min;95 ℃变性10 min;95 ℃变性15 s,58 ℃延伸30 s,反应进行40个循环。将所得数据用2-△△Ct法进行数据分析[15]。选用京411小麦0 h叶片中TaWRKY26表达量作为对照。
2 结果与分析
2.1 TaWRKY26基因的克隆
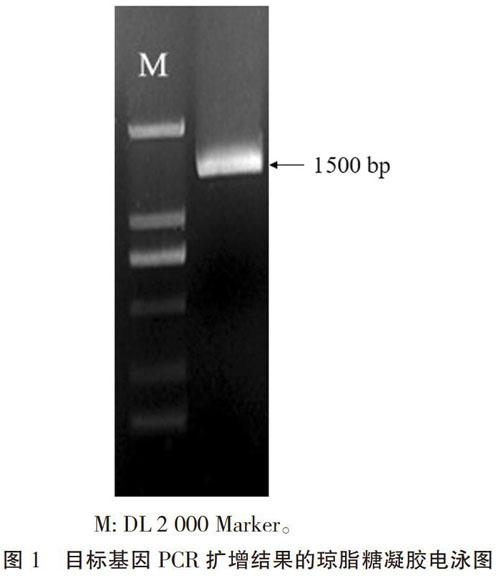
提取白粉菌侵染24 h的Brock叶片总RNA,反转录合成cDNA后用于基因扩增,扩增结果如图1所示,在1 500 bp位置有一明显条带,将该条带克隆、测序。测序结果(图2)显示,该片段长1 485 bp,与片段F5进行比对分析,发现包含F5片段;预测开放阅读框(Open Reading Frame,ORF)長1 392 bp,编码463个氨基酸,分子量为49.8 kDa;通过氨基酸序列分析发现,该序列具有两个WRKY基因的典型结构域:WRKYGQK结构。
2.2 多序列比对分析和进化树构建
利用DNAMAN软件对TaWRKY26与针茅(Stipa purpurea,Sp)、黍(Panicum hallii,Ph)、粗山羊草(Aegilops tauschii,At)和小麦(Triticum aestivum,Ta)中的WRKY转录因子家族其他基因进行多序列比对分析(图3)发现,TaWRKY26与这些基因均具有较高同源性。进一步的系统进化树分析(图4)发现,TaWRKY26与粗山羊草中WRKY26基因亲缘关系最近,与小麦中的TaWRKY2次之,这一比对结果与本实验室前期工作中的Blastx比对结果一致,故将其命名为TaWRKY26,并将扩增得到的序列全长上传至GenBank进行登记,序列号为MK358190。
2.3 白粉菌胁迫下TaWRKY26表达模式分析
为了验证TaWRKY26是否参与小麦抗白粉病过程,利用实时荧光定量PCR技术,分别分析白粉菌侵染不同时间段TaWRKY26基因在不同抗性小麦中表达量趋势变化。由图5可知,在感病小麦京411中,白粉菌侵染后,TaWRKY26基因表达量呈升高-降低-升高-降低-升高的波动变化趋势,至12 hpi 时达到高峰,为对照的4.2倍,至168 hpi与对照组(京411 0 hpi)接近;在抗病小麦Brock中,TaWRKY26基因表达量亦呈升高-降低-升高-降低-升高-降低-升高的波动变化趋势,除12 hpi外,在其他各时间点均高于同时期感病小麦京411,通过多重比较发现,在0,24,72,120 hpi时表达量差异显著(P<0.05),2,168 hpi时表达量差异极显著(P<0.01),且在24 hpi表达量最高,为Brock叶片0 hpi时的2.9倍,对照组(京411 0 hpi)的13.2倍;在抗病小麦NILs中,受白粉菌侵染后,TaWRKY26表达量呈降低-升高-降低-升高-降低-升高-降低的波动变化趋势,在24 hpi时表达量达到最高,为NILs叶片0 hpi时的2.0倍,对照组(京411 0 hpi)的5.6倍,NILs中TaWRKY26的表达变化介于感病小麦京411和抗病小麦Brock之间,但更趋向于抗病亲本Brock。
3 结论与讨论
植物在受到逆境胁迫时会发生一系列的抗逆反应,这些抗逆反应受多种转录因子的调控,同时一种转录因子能响应多种抗逆反应,即一因多效。WRKY转录因子最早是从甘薯中克隆出来的[16]。后来在多种植物中克隆到了WRKY转录因子家族并对其功能进行了研究,发现WRKY家族多参与植物抗逆、生物胁迫和衰老等过程。在小麦中,多个WRKY转录因子已被证实参与了小麦抗逆反应:吴华玲等[17]发现TaWRKY10、TaWRKY19-a、TaWRKY71和TaWRKY72-b可能是小麦抗逆调控网络中重要的交流因子;还有研究发现,TaWRKY46能够提高小麦植株的抗氧化能力[18];TaWRKY35过表达的小麦植株耐盐性会显著提高[19]。
在植物对白粉菌的响应中,WRKY基因也发挥着重要的作用,在大麦中沉默HvWRKY10、HvWRKY19和HvWRKY28后其对白粉菌的易感性显著上升,而将这3个基因瞬时过表达后能够正向调控大麦对白粉菌的响应[20];在拟南芥中发现的WRKY18和WRKY40会抑制植株对白粉病的响应[21];小麦中TaWRKY34在抗白粉病近等基因系中受白粉病菌诱导后表现出了上调趋势,证明TaWRKY34可能参与了对白粉菌的防卫应答反应[22];MdWRKY40b长时间高量表达可能是导致富士苹果易感白粉病的主要原因之一[23]。上述试验都表明WRKY基因参与植物对白粉菌胁迫的响应。
目前,有关WRKY转录因子的研究均显示WRKY26基因可能与植物抗逆密切相关。拟南芥中WRKY26基因能够提高植株的耐热性[24];葡萄中WRKY26基因可能应答多种植物逆境胁迫相关信号[25]等。本试验克隆到的TaWRKY26具有两个典型的WRKYGQK结构,经对比发现与粗山羊草中WRKY26基因同源性最高;从系统进化树可以看出小麦中与TaWRKY26同源性最高的基因为TaWRKY2。TaWRKY2在植物中多参与抗逆应答反应,如将TaWRKY2过表达载体转化拟南芥,转基因植株的耐盐性和耐旱性均显著提高[26];将TaWRKY2转入短柄草中,在室温条件下能够提高短柄草对赤霉病的抵抗性[27];在小麦中过表达TaWRKY2,小麦的耐旱性显著提高[28]。说明小麦中的TaWRKY2基因参与植物耐盐、耐干旱及对赤霉病的应答,TaWRKY26与粗山羊草中WRKY26基因和小麦TaWRKY2基因均有较高同源性,可推断其可能在小麦-白粉菌互作过程中发挥重要功能。
前期工作中发现栽培小麦Brock中含有一个抗白粉病新基因,且由显性单基因控制[29],但Brock与我国大多数栽培小麦花期不遇,故利用Brock进行小麦抗白粉病育种工作进展缓慢,在Brock中开展抗病基因的相关研究对生产上合理利用该品种资源有重要意义。小麦抗白粉病近等基因系是进行小麦抗白粉病遗传机制研究的宝贵材料,理论上,近等基因系与轮回亲本遗传背景相似,仅有一对抗病基因的差别,本研究通过对TaWRKY26表达模式进行分析后发现,其表达量在抗病小麦Brock中除12 hpi外均高于感病小麦京411,在抗白粉病小麦近等基因系Brock/京4117(抗病)中介于感病亲本京411和抗病亲本Brock之间,且更偏向抗病亲本Brock,说明TaWRKY26受Brock中抗白粉病基因的正调控作用,可能参与小麦的抗白粉病应答反应,在小麦对白粉菌侵染的应答通路中起到一定的作用。
參考文献:
[1]CONNER R L, KUZYK A D, SU H. Impact of powdery mildew on the yield of soft white spring wheat cultivars[J]. Canadian journal of plant science, 2003, 83(4):725-728.
[2]BIGEARD J, COLCOMBET J, HIRT H. Signaling mechanisms in pattern-triggered immunity(PTI)[J]. Mol plant, 2015, 8: 521-539.
[3]COUTO D, ZIPFEL C. Regulation of pattern recognition receptor signalling in plants[J]. Nat rev immun, 2016, 16: 537-552.
[4]严霞,牛晓磊,陶均.病原菌诱发的植物先天免疫研究进展[J].分子植物育种,2018,16(3):821-831.
[5]CHEVAL C, ALDON D, GALAUD J P, et al. Calcium/
calmodulin-mediated regulation of plant immunity[J].Biochim biophys acta, 1833(7): 1766-1771.
[6]Eulgem T, Rushton P J, Robatzek S, et al. The WRKY superfamily of plant transcription factors[J]. Trends in plant science, 2000, 5(5):199-206.
[7]余迪求,陈利钢,张利平.转录调控因子WRKY超级家族:起源、结构和功能[J].植物分类与资源学报,2006,1(1):73-81.
[8]陈彩慧,任艳芳,肖蓉,等.樟树WRKY转录因子的克隆与表达分析[J].分子植物育种,2018,16(15):4872-4879.
[9]MEHANATHAN M, BONTHALA V S, ROHIT K, et al. Global analysis of WRKY transcription factor superfamily in Setariai dentifies potential candidates involved in abiotic stress signaling[J]. Frontiers in plant science, 2015, 6:910.
[10]叶玉洁. MaWRKY26与MaVQ5蛋白互作调控MeJA诱导香蕉果实耐冷性的机制[D].广州:华南农业大学, 2016.
[11]ZHOU L, WANG N N, GONG S Y, et al. Overexpression of a cotton (Gossypium hirsutum) WRKY gene, GhWRKY34, in Arabidopsis enhances salt-tolerance of the transgenic plants[J]. Plant physiology & biochemistry, 2015, 96:311-320.
[12]LIU S A, KRACHER B, ZIEGLER J, et al. Negative regulation of ABA signaling by WRKY33 is critical for Arabidopsis immunity towards Botrytis cinerea 2100[J].Elife, 2015, 4:e07295.
[13]CHLO? M, C?LINE L, KAPPEL C, et al. Over-Expression of VvWRKY1 in grapevines induces expression of jasmonic acid pathway-related genes and confers higher tolerance to the downy mildew[J]. Plos one, 2013, 8 (1): e54185.
[14]HIROAKI A, NOBUAKI I, TAKAAKI N, et al. Nicotiana benthamiana MAPK-WRKY pathway confers resistance to a necrotrophic pathogen Botrytis cinerea[EB/OL].[2018-11-29].http://www.tandfonline.com/doi/full/10.1080/15592324.2016.
1183085.
[15]KENNETH J L, THOMAS D S. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method[J]. Methods, 2001, 25(4):402-408.
[16]ISHIGURO S, NAKAMURA K. Characterization of a c DNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5' upstream regions of genes coding for sporamin and beta-amylase from sweet potato[J].Molecular and general genetics, 1994, 244(6): 563-571.
[17]吴华玲,倪中福,姚颖垠,等.15个普通小麦WRKY基因的克隆与表达分析[J].自然科学进展,2008,18(4):378-388.
[18]卫琳.小麦盐、碱胁迫应答基因TaWRKY46和TaNRT1.2的功能研究[D].济南:山东大学,2018.
[19]刘自成,苗丽丽,王景一,等.普通小麦转录因子基因TaWRKY35的克隆及功能分析[J].中国农业科学, 2016, 49(12):2245-2254.
[20]WISE R P . HvWRKY10, HvWRKY19, and HvWRKY28 regulate Mla-triggered immunity and basal defense to barley powdery mildew[J]. Mol plant microbe interact, 2012, 25(11):1492-1505.
[21]PANDEY S P, Roccaro M, MORITZ S, et al. Transcriptional reprogramming regulated by WRKY18 and WRKY40 facilitates powdery mildew infection of Arabidopsis[J].The plant journal, 2010, 64(6):12.
[22]秦偉,赵光耀,曲志才,等.小麦白粉病菌诱导的TaWRKY34基因的鉴定与分析[J].作物学报,2010,36(2):249-255.
[23]罗昌国,袁启凤,裴晓红, 等. 富士苹果MdWRKY40b基因克隆及其对白粉病的抗性分析[J].西北植物学报, 2013, 33(12):2382-2387.
[24]LI S J, FU Q T, CHEN L G, et al. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermotolerance[J]. Planta, 2011, 233(6):1237-1252.
[25]于和平.葡萄WRKY26、WRKY27基因启动子克隆及功能分析[D].大连:辽宁师范大学, 2013.
[26]NIU C, WEI W, ZHOU Q, et al. Wheat WRKY genes TaWRKY2 and TaWRKY19 regulate abiotic stress tolerance in transgenic Arabidopsis plants[J]. Plant cell & environment, 2012, 35(6):1156-1170.
[27]郭秀秀,王亮,苏培森,等.利用短柄草进行TaWRKY2的抗赤霉病功能分析[J].山东农业大学学报:自然科学版,2017,48(4):570-575.
[28]GAO H, WANG Y, XU P, et al. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat[J]. Front plant sci,2018,9:997.
[29]WANG Z Y, ZHAO P, CHEN H, et al. Identification of RAPD markers and development of SCAR markers linked to a powdery mildew resistance gene, and their location on chromosome in wheat cultivar brock[J]. Plant production science, 2005, 8(5):578-585.
