橡胶树野生种质资源抗寒性评价及其与生长的相关性分析
2019-04-16李小琴张凤良毛常丽
李小琴 张凤良 杨 湉 赵 祺 毛常丽 吴 裕
( 云南省热带作物科学研究所,云南 西双版纳 666100)
巴西橡胶树(Hevea brasiliensis)原产南美亚马逊河流域的热带雨林中,是典型的热带乔木树种。中国植胶区地处热带北缘,属于非传统植胶区,低温是限制该区域橡胶产业发展的主要因素。云南植胶区由于其纬度更偏北,海拔更高,冬季热量不足,在植胶历史上,已经出现过7次比较重的寒害,其中1970—1976年,就出现了3次,且有2次灾害最重[1-4]。橡胶树的生长(生产)周期长达30 a以上,研究表明,云南植胶区每间隔10~15 a会出现一次寒害,寒害的出现呈现周期性。为此,筛选培育抗寒高产的橡胶树品种成为了云南天然橡胶产业发展的重要任务。
低温能够引起橡胶树细胞结构、原生质胶体特性、水分状况、细胞渗透压、光合作用、呼吸作用、物质代谢和保护酶系统等发生一系列改变,最终影响植株的生长发育及胶乳合成[5]。目前,橡胶树抗寒种质筛选的方法主要有抗寒前哨梯度苗木系比鉴定法、自然低温鉴定法及人工模拟低温鉴定法等,鉴定指标包括形态、生理生化及分子细胞等[6-13]。橡胶树抗寒性鉴定多集中在对品种及优良无性系之间的比较,根据以往的研究可知,生长势较好、速生的品种往往抗寒性都不弱,如云研77-2、IAN873、热垦525、热垦628、云研75-1、RRIM524等[14-18],而就橡胶树生长与抗寒性之间关系的研究较少。本研究选择生长量有所差异的12份种质资源进行人工模拟低温寒害处理,测定其生理生化指标,利用隶属函数对抗寒性进行综合评价,结合生长量进行相关性分析,以期为橡胶树大规模种质资源的抗寒性鉴定提供一个新的思路,也为橡胶树抗寒育种选择优良材料。
1 材料与方法
1.1 试验方法
选取保存于云南省热带作物科学研究所“农业部景洪橡胶树种质资源圃”内的部分橡胶树野生种质资源,2016年采集芽条,以GT1开放授粉实生苗为砧木繁殖成无性系,芽接成活后,于2017年3月锯杆,按株行距1 m×1 m育苗保留,苗圃地按照常规育苗管理,苗木生长良好。根据苗期生长量,选择其中12份种质资源进行抗寒生理测定,编号为1~12号。
2018年1月15日,每个无性系随机选取4株长势均匀的植株,剪下从上而下第2蓬叶的枝条,随即放入冷藏取样箱内,带回实验室。依次用自来水和蒸馏水冲洗干净,吸水纸吸干水分,再将每根枝条剪成均匀的3段,同一无性系随机分为4组,每组3段,用塑封袋包好,并做好标记。用可调温冰柜设置4个温度梯度:4(对照)、0、-2 ℃和-4 ℃,将枝条分别放于设置好温度的冰柜进行低温处理,持续12 h后,取出放至室温再进行样品制备。
1.2 测定方法
相对电导率的测定:将低温处理后的枝条用滤纸吸干水分,避开芽眼,准确裁取1 cm×1 cm的小方块枝条韧皮部,同一低温处理同一无性系对3段枝条混合取样共裁取6片,每2片放入一个试管中,加入双重蒸馏水20 mL,混合均匀,设3次重复,室温放置24 h,期间摇匀数次。用SG3-ELK型便携式电导率仪先测出初始电导值(C1),再盖上试管塞,沸水浴30 min,取出冷却至室温后,再测出终电导值(C2)。以相对电导率表示细胞膜透性大小,相对电导率按以下公式进行计算:
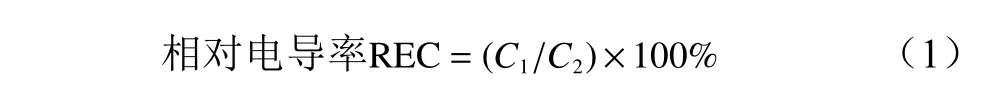
生理生化指标测定方法[19]:丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法;游离脯氨酸(Pro)含量采用酸性茚三酮染色法;可溶性蛋白含量采用考马斯亮蓝法;可溶性糖含量采用蒽酮比色法。各指标均采用枝条韧皮部(鲜质量)进行测定,使用苏州科铭生物技术有限公司生产的试剂盒进行测定及相关含量计算。
径围测量方法:用软尺于采样前对其1年生苗木1.0 m高处的粗度进行测量,每个橡胶树无性系随机测量50株,求其平均值。
1.3 分析方法
采用Excel 2003对数据进行整理作图,用SPSS23.0对数据进行统计分析。抗寒性的综合评价采用模糊数学中的隶属函数方法进行计算,隶属函数值的计算方法如下[20]:
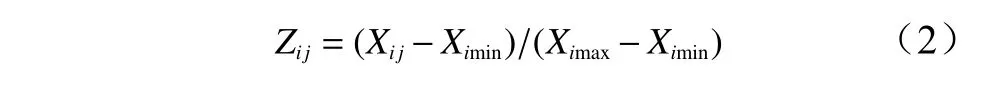
如果某一指标与抗寒性为负相关,用反隶属函数计算器抗寒隶属函数值,计算方法为:
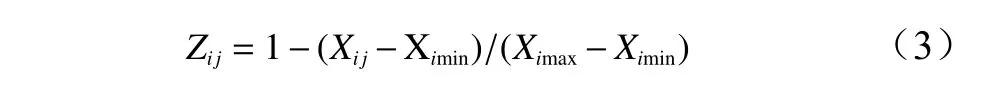
其中:Zij为第i个无性系第j个测定指标的抗寒隶属函数值;Xij为第i个无性系第j个指标的测定值;Ximax和Ximin分别为全部无性系第j项指标的最大值和最小值。
2 结果与分析
2.1 不同低温胁迫对相对电导率的影响
相对电导率是反映植物组织受冻后细胞原生质膜透性大小的重要指标,以相对电导率表示植物在低温伤害下细胞质膜透性变化是植物抗寒性鉴定常用的方法[21-22]。
经不同低温胁迫后,12个橡胶树无性系枝条的相对电导率变化情况见图1,0 ℃处理与对照4 ℃相比,变化最小,增幅最小的为1号和10号,增幅仅1%;增幅最大的为4号,增幅为39%。相对电导率0 ℃均值(39.27%)与4 ℃(34.02%)相比总体增幅为15%,说明0 ℃低温对多数橡胶树枝条组织伤害较小,细胞膜透性变化小。在-2 ℃时,部分无性系相对电导率上升明显,超过50%有3个号,包括4号(61.38%)、12号(57.52%)和10号(53.14%);而小于40%的有1号、3号和6号,表现出较强的耐低温能力。到-4 ℃低温时,各无性系与对照(4 ℃)相比相对电导率出现最高增幅且均已超过50%,总体均值已达68.25%,说明-4 ℃低温对橡胶树枝条组织已经造成严重伤害。从整个变化趋势可以看出橡胶树枝条对-2~-4 ℃范围的低温极度敏感,多数都表现出对此低温的不耐受性。
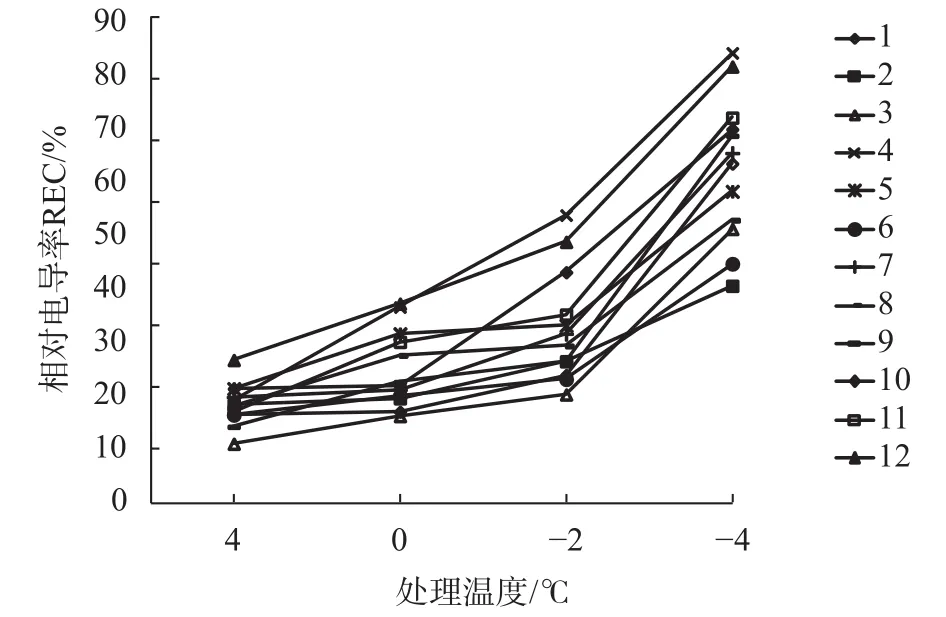
图1 不同低温胁迫下相对电导率的变化趋势Fig. 1 The changing trend of relative electric conductivity under different low temperature stress
2.2 不同低温胁迫对丙二醛含量的影响
橡胶树各无性系枝条经不同低温胁迫后丙二醛含量的变化如图2,各无性系丙二醛含量均表现为“先升高后降低”的变化趋势。0 ℃时,与4 ℃对照相比总体增长较慢,但各无性系间差异较大,增幅在5%~37%之间,MDA含量变幅为9.03~10.58 nmol/g。所有无性系MDA含量的最大值都出现在-2 ℃,其中6号MDA含量最高,为12.69 nmol/g,且增幅也最大,比对照增长64%,而2号值最低,为9.56 nmol/g。-4 ℃时,组织受寒害严重,细胞膜脂过氧化程度下降,MDA含量出现不同程度的下降,基本达到同一水平。在整个变化过程中,2号MDA含量最低,总体均值仅9.11 nmol/g,增幅较小,表明其膜的过氧化程度低,抗寒能力强;而6号和12号含量高,均值分别为10.41 nmol/g和10.31 nmol/g,其抗寒性较差。
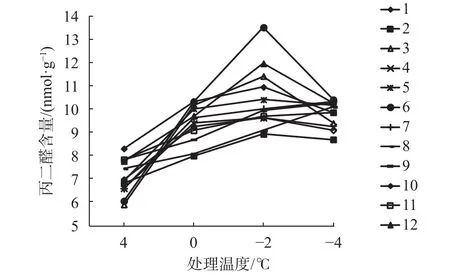
图2 不同低温胁迫对丙二醛含量的影响Fig. 2 The influence of malondialdehyde content under different low temperature stress
2.3 不同低温胁迫对脯氨酸含量的影响
在逆境条件下,植物体内游离脯氨酸含量显著增加,脯氨酸增加量在一定程度上反映了其抗逆性,抗寒性强往往积累较多的脯氨酸。各无性系脯氨酸含量变化情况见图3,0 ℃与对照4 ℃脯氨酸含量差异极小,均值分别为46.65 μg/g和47.66 μg/g,表明0 ℃是组织所能承受的低温,并未能促使其产生脯氨酸含量增加的生理反应。在-2 ℃多数无性系出现大幅度上升,总体均值为68.02 μg/g,增幅达43%,除2、5、8及10号外,其余的均达到最大值,脯氨酸含量最大的为3号(118.02 μg/g),增幅为109%。-4 ℃ 时,2、5、8 及10号出现极大值,其余均有不同程度的下降,差异较大,脯氨酸含量为40.22~101.01 μg/g。整个低温处理中,3号脯氨酸含量最高,均值82.49 μg/g,增幅较大,其抗寒性强。

图3 不同低温胁迫对脯氨酸含量的影响Fig. 3 The influence of proline content under different low temperature stress
2.4 不同低温胁迫对可溶性蛋白含量的影响
多数植物在低温锻炼期间,细胞内可溶性蛋白含量与抗寒性之间表现出明显的正相关[23-25]。由图4可知,橡胶树枝条组织可溶性蛋白含量变化趋势存在较大差异,主要分为2类,一类是“降—升—降”,有2、5、10、6及12号等5个无性系;另一类是“升—升—降”,有7个无性系。0 ℃时,多数无性系可溶性蛋白含量变化较小,总体均值为 100.08 μg/mL,与对照 94.06 μg/mL 相比,增幅为13%,其中增幅最大的1号,为65%,可溶性蛋白含量158.85 μg/mL,表明其在受低温胁迫时,组织通过快速升高可溶性蛋白含量来增加抗寒性。到-2 ℃时,各无性系均达到最大峰值且差异极显著,可溶性蛋白含量在84.00~181.95 μg/mL之间,总体均值为128.23 μg/mL,增幅达41%,说明此低温已对组织造成了较大伤害。-4 ℃时,可溶性蛋白含量一致下降,下降幅度除1号较小外,其余无性系均较大,此时组织受极严重伤害,失去了利用可溶性蛋白含量抵御低温的生理调节能力。1号在整个低温胁迫中可溶性蛋白含量最高,均值为143.69 μg/mL,增幅最大75%,表现出对低温的极强耐受力,抗寒性最强;同时,4、7和12号,含量都低于60.00 μg/mL,耐低温能力最弱。
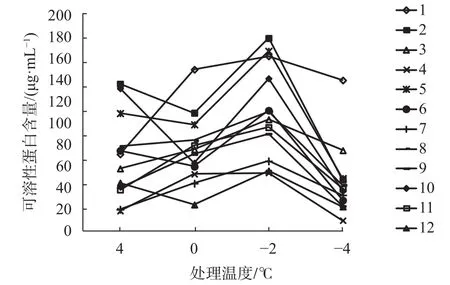
图4 不同低温胁迫对可溶性蛋白含量的影响Fig. 4 The influence of soluble protein content under different low temperature stress
2.5 不同低温胁迫对可溶性糖含量的影响
可溶性糖是一种重要的渗透调节物质,可溶性糖的积累对植物受冷冻害有保护作用,其含量与抗寒性呈正相关。各无性系可溶性糖含量变化情况见图5。
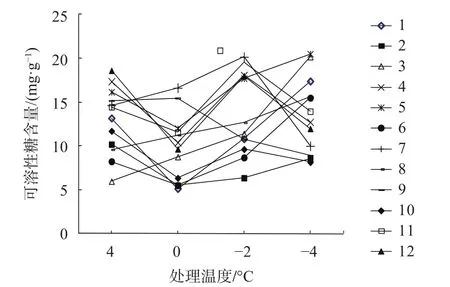
图5 不同低温胁迫对可溶性糖含量的影响Fig. 5 The influence of soluble sugar content under different low temperature stress
由图5可看出,不同无性系经低温处理可溶性糖含量变化趋势差异较大,并未呈现一致规律,所有无性系在整个处理期间都在5.00~20.50 mg/g范围内。0 ℃时,除3、7、8和9号外,其余均有不同程度的下降;到-2 ℃时,除9号外,均表现为上升趋势,整体均值最大,为13.65 mg/g,与4 ℃对照均值12.94 mg/g相比,仅增长了5.5%,进一步说明可溶性糖含量在整个低温胁迫过程中含量变化不大;-4 ℃时,有50%呈下降趋势,而1、2、3、5、6和8号表现为上升,说明这6个无性系组织内通过增加可溶性糖含量来调节抗寒能力的生理响应较晚。整个低温处理过程中,可溶性糖含量最高的是5号和7号,均值为16.59 mg/g和15.35 mg/g,最低的为2号和10号,均值为7.66 mg/g和8.96 mg/g。
2.6 利用隶属函数对抗寒性进行综合评价
以相对电导率、丙二醛含量、脯氨酸、可溶性蛋白及可溶性糖含量等5个指标的均值为依据,计算各指标的隶属函数值并进行综合评价结果见表1,测定的12份橡胶树野生种质资源抗寒性由强到弱排序为:3>5>2>8>1>11>7>9>6>4>10>12。
2.7 橡胶树无性系生长与抗寒指标相关性分析
按径围多重比较结果(表1)可将12个无性系分成3个等级:生长量最大的有2、3、4和5号;生长量中等的有1、6和8号;生长量一般的有7、9、10、11和12号。同一等级内径围差异不显著,而等级间差异均达极显著水平(P<0.01)。由抗寒性排序可知,4号和6号生长量较大,但抗寒性排序相对靠后,按保留和去除4号和6号2种方法,分别对无性系各指标隶属函数值、综合平均值及径围等7个指标进行相关性分析。
由表2可知,保留4号和6号时径围仅与隶属函数综合值呈显著正相关;相对电导率与可溶性蛋白、可溶性糖显著相关,多数指标间无显著相关性。而当去除4号和6号时,径围与隶属函数综合值呈极显著正相关,此时相关系数(0.824)明显要比前者(0.643)高很多;且分别与相对电导率、丙二醛和可溶性蛋白等指标隶属函数值呈显著或极显著相关,说明仅少部分(4号和6号)生长较好的种质资源与抗寒性之间的相关性不显著。
综上可知,径围生长量可用作橡胶树种质资源抗寒性选择的一个间接指标,但需要指出的是用径围生长量正向选择时存在部分误选,而进行淘汰选择时效果较好。如本研究以径围生长量第1等级为选择对象,则出现4号一个误选;如以第3等级为淘汰对象,则排名最前的一个也仅排到第6名,可以都做淘汰处理。
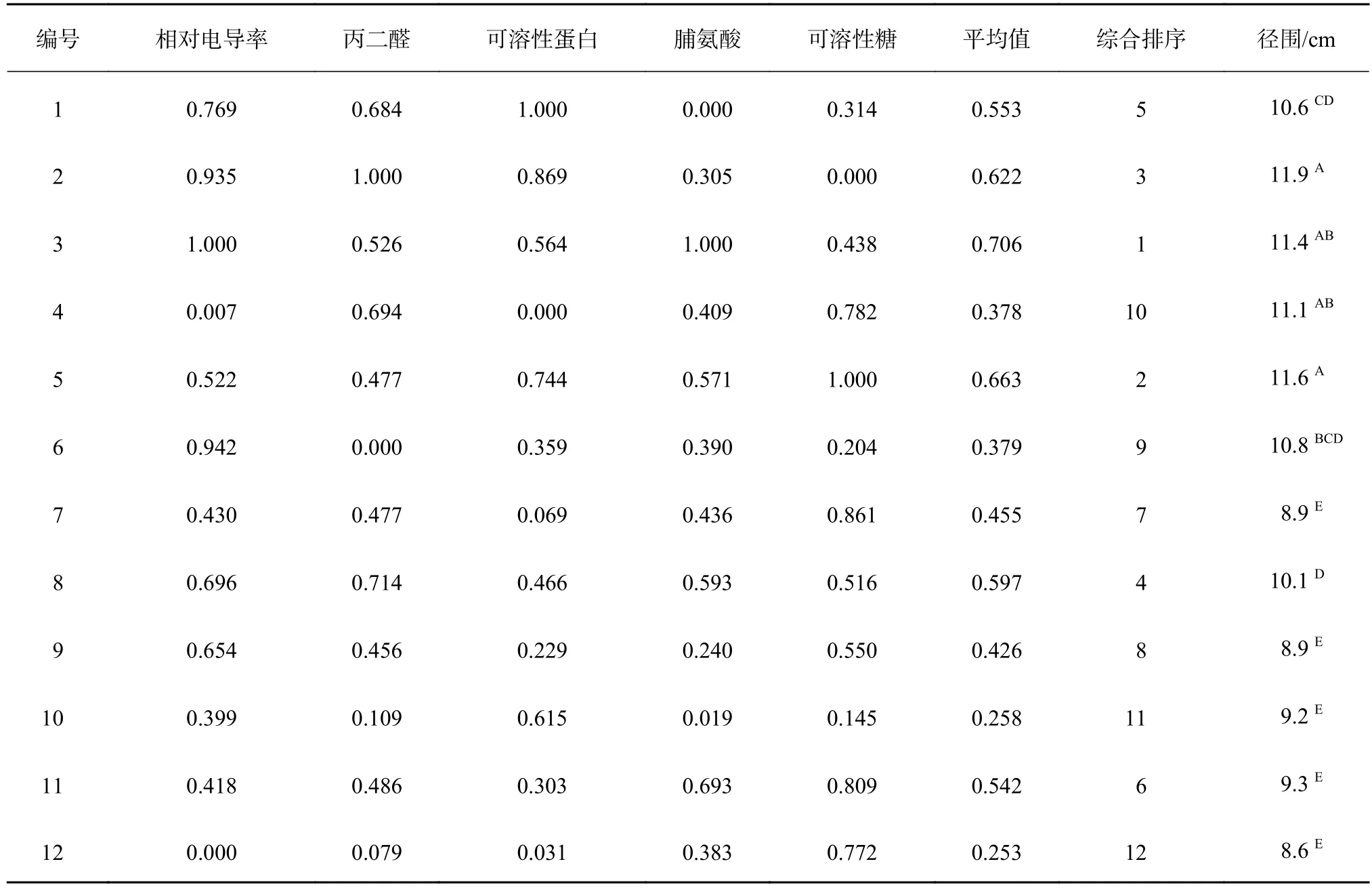
表1 利用隶属函数对不同橡胶树无性系抗寒性进行综合评价结果Table 1 The comprehensive evaluation result of cold resistance of different H. brasiliensis clones using membership function
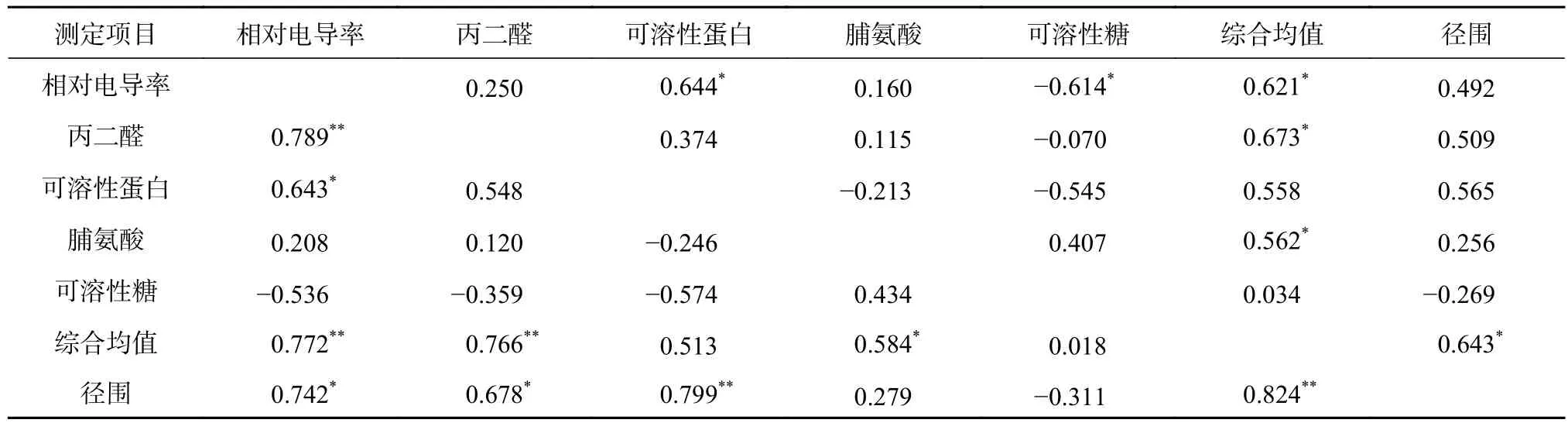
表2 橡胶树无性系生长与抗寒指标相关性统计Table 2 The statistic of correlation between growth and cold resistance indexes of H. brasiliensis clones
3 结论与讨论
橡胶树的生长发育经常会受到寒冷胁迫的影响,抗寒性比较强的品种或无性系在长期应对低温等逆境胁迫的过程中形成了功能独特的物质条件[26]。本研究表明,低温胁迫可诱导橡胶树启动抗寒响应机制,在生理上产生了一系列的变化。低温胁迫使橡胶树幼苗枝条内积累了更多的游离脯氨酸、可溶性蛋白、可溶性糖等物质,而这些渗透调节物质的积累可减轻渗透胁迫、防止膜脂和蛋白质的过氧化作用,但同时丙二醛也同步增加,不同无性系间各物质的含量变化差异较大,但整体看,这几类物质随不同低温胁迫后均呈现出了“降—升—降”的变化趋势,这与多个树种上的研究结果相一致[27-28]。
植物抗寒性是一个复杂的数量性状,在进行抗寒性鉴定时,生理指标的选择尤为重要,张卫华等[29]研究指出相对电导率、水分状况、游离脯氨酸和可溶性糖含量等指标对相思的抗旱性评价影响力会占到88%以上,尤其相对电导率成为评价其抗旱性的主导因子。本研究以相对电导率来进行选择时,抗寒性强弱综合排名靠前的相对电导率相对较小,排名靠后的较大,值得指出的是5号,综合排名为2,但相对电导率处于中等位置,6号综合排名靠后,但相对电导率较小,有待多次验证。冯建灿等[30]指出植物组织内脯氨酸含量的高低可以作为衡量喜树抗寒性强弱的指标,本研究可知,脯氨酸含量较高的几个无性系(3、5、8、11号),抗寒性综合评价都不差,但也有部分无性系如1和2号,脯氨酸含量较低,但抗寒性综合评价也较强,根据李晓宇等[31]对杨树(Populusspp.)苗期抗寒性研究得出脯氨酸含量与抗寒性关系不显著,进一步说明最好不要单一地利用脯氨酸含量这一生理指标对植物抗寒性鉴定。丙二醛是膜脂过氧化指标,表示细胞膜过氧化程度和植物对逆境条件反应的强弱,在低温胁迫下,膜的过氧化程度越低,丙二醛含量越低,抗寒性越强[32]。综上,单一指标很难代表植物复杂的抗寒性,使用多指标联合分析,才能较好地反映植物的抗寒生理过程。本研究利用隶属函数法对12个无性系的相对电导率、丙二醛含量、脯氨酸、可溶性蛋白及可溶性糖含量等5个指标进行综合评价,其抗寒性由强到弱的排序依次为:3>5>2>8>1>11>7>9>6>4>10>12。从相关性分析结果来看,选择的抗寒指标间只有相对电导率与丙二醛和可溶性蛋白含量呈显著或极显著相关性,其他大多数指标两两相关性均不强,特别是脯氨酸与其他指标间的相关系数均较小,而综合隶属函数值与大多数指标间相关系数都在0.5以上(可溶性糖除外),且与多数指标达到显著或极显著水平,说明用隶属函数值综合评价橡胶树无性系的抗寒性比单一指标较科学。
前人对林木的生长与抗寒性之间的关系也做了一些研究,刘勇等[33]研究指出三倍体毛白杨(Populus tomentosa)速生前期合理施肥,能使苗高、地径生长量变大,高径比恰当,苗木生长好,苗木的抗寒性也得到提高;解荷锋等[34]对美洲黑杨(Populus nigra)无性系的生长与抗寒性研究表明生长和抗寒性与无性系的起源有密切关系,其生长量与抗寒性呈负相关;何旭东等[35]对细叶桉(Eucalyptus tereticornis)以及杨秀艳等[36]对杨树无性系的生长和抗寒性进行了联合选择,评选出了大量速生、抗寒的无性系,表明速生性与抗寒性可以同步进行选择。本研究重点分析了橡胶树生长与抗寒性的关系,结果表明,橡胶树无性系抗寒性与径围呈显著正相关,即大多数生长势好的无性系具有较强的抗寒性,少数几个无性系例外(如4号),而生长势较差的几个无性系其抗寒性均很差(如10号和12号)。本研究因从生理生化指标方面综合评价抗寒性,故选择的无性系数量有限,但结合前人在其他树种上的部分研究结果,可将橡胶树的生长量与抗寒性相结合进行种质资源批量鉴定,建议以木材利用为主且寒害较重的生态区域选育品种时,进行超速生资源优选的策略;在以胶乳产量为主要选育目标时,先进行生长量淘汰选择,再进一步做精准鉴定。
