镉胁迫对洋常春藤生长及生理特性的影响
2019-04-16程守梦李琬婷程小毛黄晓霞
程守梦 李琬婷 程小毛 黄晓霞
( 西南林业大学园林与园艺科学学院,国家林业局西南风景园林工程研究中心,云南 昆明,650224)
随着我国工业化进程的加快,大量矿物金属被采伐利用,导致进入土壤中大量重金属并未得到妥善治理,造成了严重的土壤重金属污染[1],土壤重金属污染毒性大、难察觉、难降解[2]。据统计,我国耕地重金属污染超标率约占全国耕地的30%以上,其中以镉污染超标率最高,镉污染农田面积达1.3万hm2,约占总污染农田的7.0%[3-4],涉及我国大部分地区,主要分布在我国东南、西南地区[5]。镉污染会影响土壤的理化性质;阻碍植物根系对养分和水分的吸收[6],损害植物细胞渗透和生理代谢能力,抑制生长,降低产量,甚至引起植物死亡[7]。近几年来我国类似“镉米”的粮食安全问题频发[8],因此,进一步对植物修复技术研究,对于重金属污染土壤的治理及农业生态环境安全具有重要意义[9]。
洋常春藤(Hedera helix),五加科(Araliaceae)常春藤属(Hedera),多年生常绿藤本,耐荫、抗旱[10-11],易繁殖且生长迅速,适应性强,在园林中多用作地被及坡面墙体绿化等。目前关于洋常春藤研究主要有对其栽培品种药用质量评价[12]、皂苷类物质的药理应用[13]、干旱研究[11,14]及扦插栽培[15-16]等方面,而关于其对土壤重金属污染的抗性研究较少。本研究以洋常春藤为供试材料,通过人工控制试验,研究不同镉浓度对洋常春藤的生长及部分生理特性的变化。通过研究不同浓度镉胁迫下洋常春藤的生长生理指标的变化,探讨洋常春藤的耐镉性,以期为植物修复重金属污染土壤技术奠定一定的理论基础,并为镉污染地区绿化植物的科学选择提供参考依据。
1 材料与方法
1.1 试验材料
于2016年4月初选取当年生洋春藤半木质化枝条扦插于营养袋中,生根后于6月中旬移栽至不含蔗糖和琼脂的MS培养基的组培瓶中,进行水培培养,pH值为5.5~6.5,同时用黑色的聚乙烯膜包裹组培瓶外部,防止光对根系的影响,培养液隔天更换1次。
1.2 试验设计
待植株生长稳定后,将生长势一致的幼苗放于人工气候箱中培养,进行Cd2+胁迫,设定5个Cd2+胁迫浓度,分别为 CK(0 μmol/L)、Cd1(20 μmol/L)、Cd2(80 μmol/L)、Cd3(200 μmol/L)、Cd4(400 μmol/L),温度25/20 ℃(昼/夜),相对湿度80%,日照时数为 12 h/天(光强≥56.6 μmol/(m2·s))。每个浓度10株,每株1瓶,重复3次。处理60 d后测定各项指标。
1.3 测定方法
1.3.1 生长指标测定
胁迫结束后,选取长势一致的幼苗,用游标卡尺(精确到0.02 mm)和卷尺(精确到1 mm)测定株高、节间距、节间数、叶片数;洗净根系,量取根长,后将根、茎、叶分别放入80 ℃的烘箱烘至质量恒定,用分析天平(精确到0.0001 g)称量根、茎、叶及总生物量,每个处理重复测定5次。
1.3.2 生理指标测定
丙二醛(MDA)含量测定参照Hodges等[17-18]的方法,脯氨酸(Pro)含量测定参照Bates等[19]的方法,可溶性蛋白含量测定参照Bradford[20]的方法,超氧阴离子产生速率测定参考孙胜等[21]的方法。
取0.2 g叶片于5 mL提取缓冲溶液(50 mmol Na2HPO4-NaH2PO4缓冲液,pH为7.0,含1 mmol EDTA,体积分数为0.05%的Trion X-1,质量体积分数为2%的不溶性聚乙烯比咯烷酮)中研磨成匀浆,经过1000 r/min离心20 min,取上清液测定抗氧化酶活性[22]。过氧化物酶(POD)活性参照Lin等[23]的方法测定(酶活单位μmol/(min·g));过氧化氢酶(CAT)活性采用紫外分光光度法[24]测定(酶活单位U/g);抗坏血酸过氧化物酶(APX)的活性参照Nakano等[25]的方法测定(酶活单位 μmol/(min·g));超氧化物歧化酶(SOD)活性参照 Giannopolitis等[18,26]的方法测定(酶活单位U/g),每个指标重复测定3次。
1.4 数据处理
用SPSS 19对数据进行一元方差分析,Ducan’s检验法对平均数进行多重比较,EXCEL绘制图表。
2 结果与分析
2.1 不同镉浓度对洋常春藤生长指标的影响
从表1可知,随着Cd2+浓度的增大,株高逐渐降低。CK的株高最高,最矮为Cd4,与CK相比显著降低28.68%(P<0.05)。Cd1节间距,比CK增加了10.9%,差异不显著,而Cd4的节间距最短,比CK显著减少了22.5%(P<0.05)。Cd2洋常春藤节间数最少,比CK下降了37.5%(P<0.05)。Cd1、Cd2和Cd3的叶片数量分别比CK显著降低了24%、20.6%和29%。与其他指标不同,Cd2+胁迫会促进洋常春藤根系增长,其中Cd2的根系最长,比CK长1.5倍。
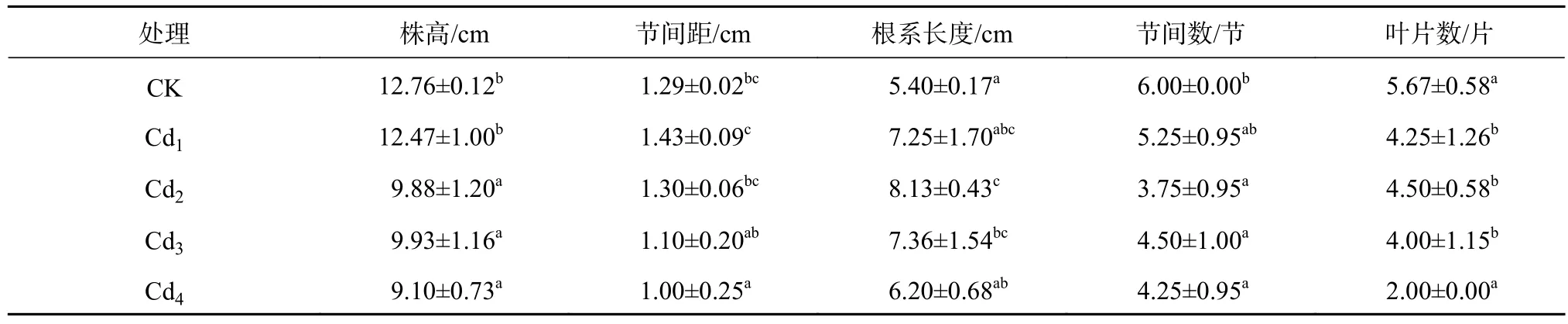
表1 不同镉浓度对洋常春藤生长指标的影响Table 1 Effects of different Cd2+ concentrations on the index of growth of H.helix
2.2 不同镉浓度对洋常春藤生物量的影响
由表2可知,随着Cd2+胁迫浓度的升高,洋常春藤根系的生物量呈先降后升再降的趋势。Cd3根系生物量最高,比CK显著高了约86%(P<0.05)。茎、叶、总生物量随着胁迫浓度增加而下降。Cd4茎、叶、总生物量均达到最低,茎生物量比CK降低20%,差异不显著,而叶生物量比CK降低了83.0%,总生物量与CK比总生物量显著降低36.13%(P<0.05)。
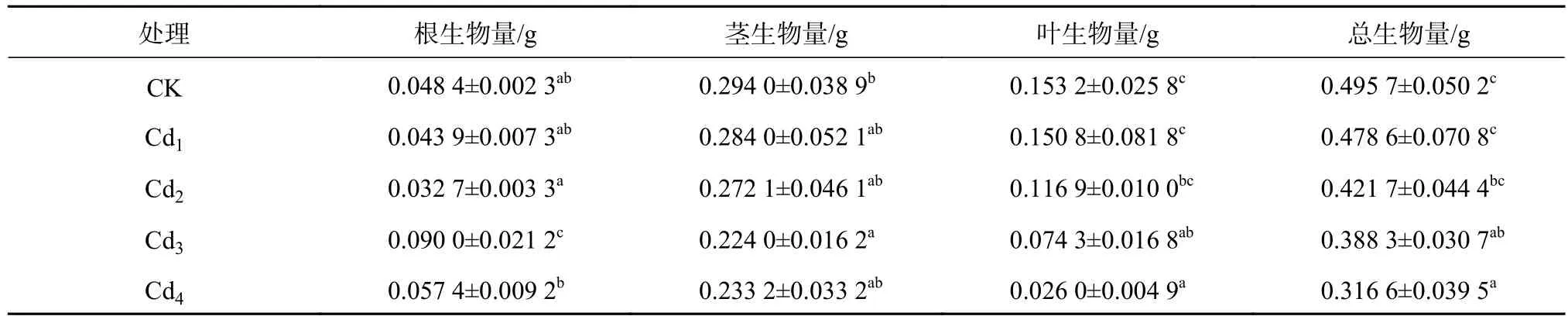
表2 不同镉浓度对洋常春藤生物量的影响Table 2 Effects of different Cd2+concentrations on biomass of H.helix
2.3 不同镉浓度对洋常春藤叶片生理指标的影响
从图1可知,Cd2+处理下,洋常春藤的含量均高于CK,呈增加的趋势,其在中、低Cd2+胁迫下变化不大,在高Cd2+胁迫下变化显著(P<0.05),Cd4的值最大,比 CK高 72.3%;MDA含量比CK增加56.2%;而Cd3的MDA含量最少,差异不显著。随着Cd2+浓度的增大,可溶性蛋白含量也增加,Cd3和Cd4的可溶性蛋白含量分别比CK增加了约158.1%和202.1%;脯氨酸含量也呈上升趋势,Cd1的脯氨酸含量增加不明显,Cd3、Cd4的脯氨酸含量分别比CK增加了约4倍和6倍。
2.4 不同镉浓度对洋常春藤叶片抗氧化酶活性的影响
由图2可见,在Cd2+胁迫下,洋常春藤叶片中的CAT表现为先增后降的趋势,但均高于CK,在Cd2下CAT活性达到最高,比CK增加4倍,而在Cd4胁迫下CAT活性出现一定程度的下降,但仍比CK的CAT活性增加了约2倍。POD活性随着Cd2+浓度的增加而增加,呈不断上升的线性关系,Cd4的POD活性最高,比CK显著高14.7倍(P<0.05)。APX除在Cd2下活性出现下降,低于CK外,大体呈上升趋势;Cd2的APX活性最低,但与CK比较,差异不显著;Cd4的APX活性最高,比CK高约2倍。Cd4的SOD活性与CK比较无显著差异,而在低、中Cd2+胁迫下SOD活性均显著低于CK(P<0.05)。
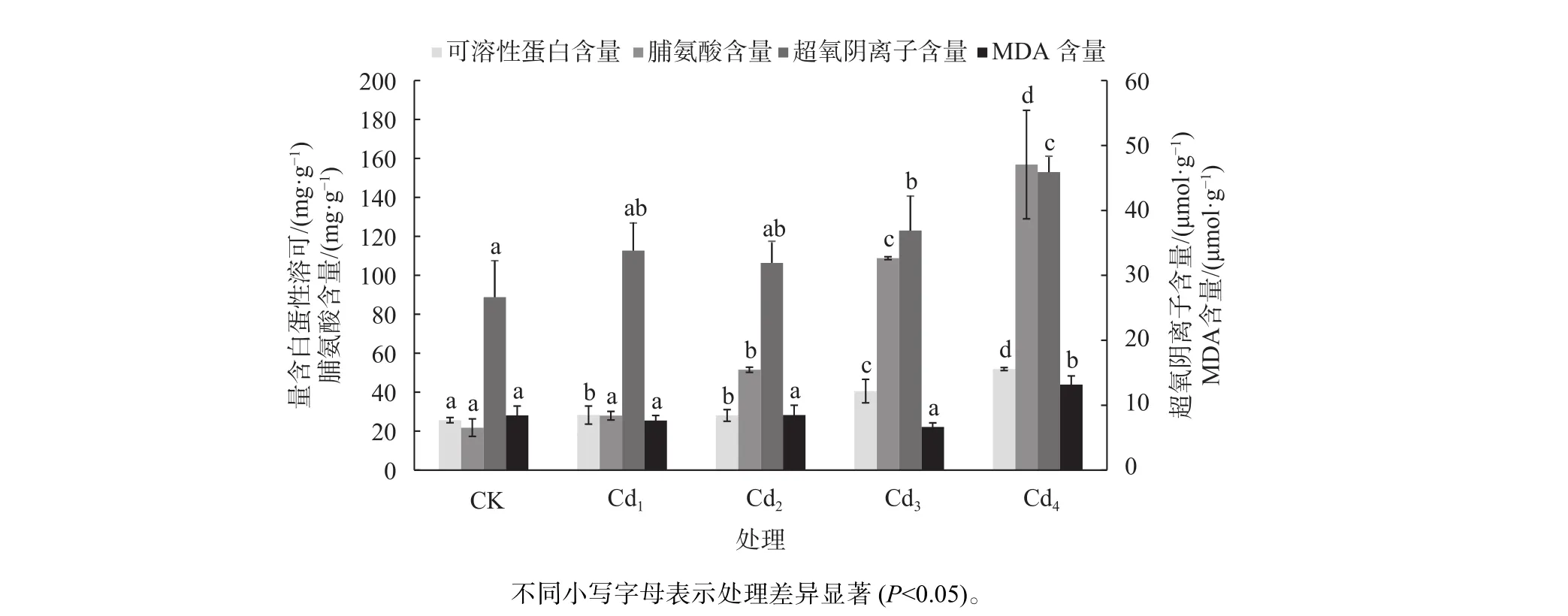
图1 不同镉浓度对洋常春藤 、MDA可溶性蛋白、脯氨酸含量的影响Fig. 1 Effects of different Cd2+ concentrations on superoxide anion, MDA soluble protein and proline content of H.helix
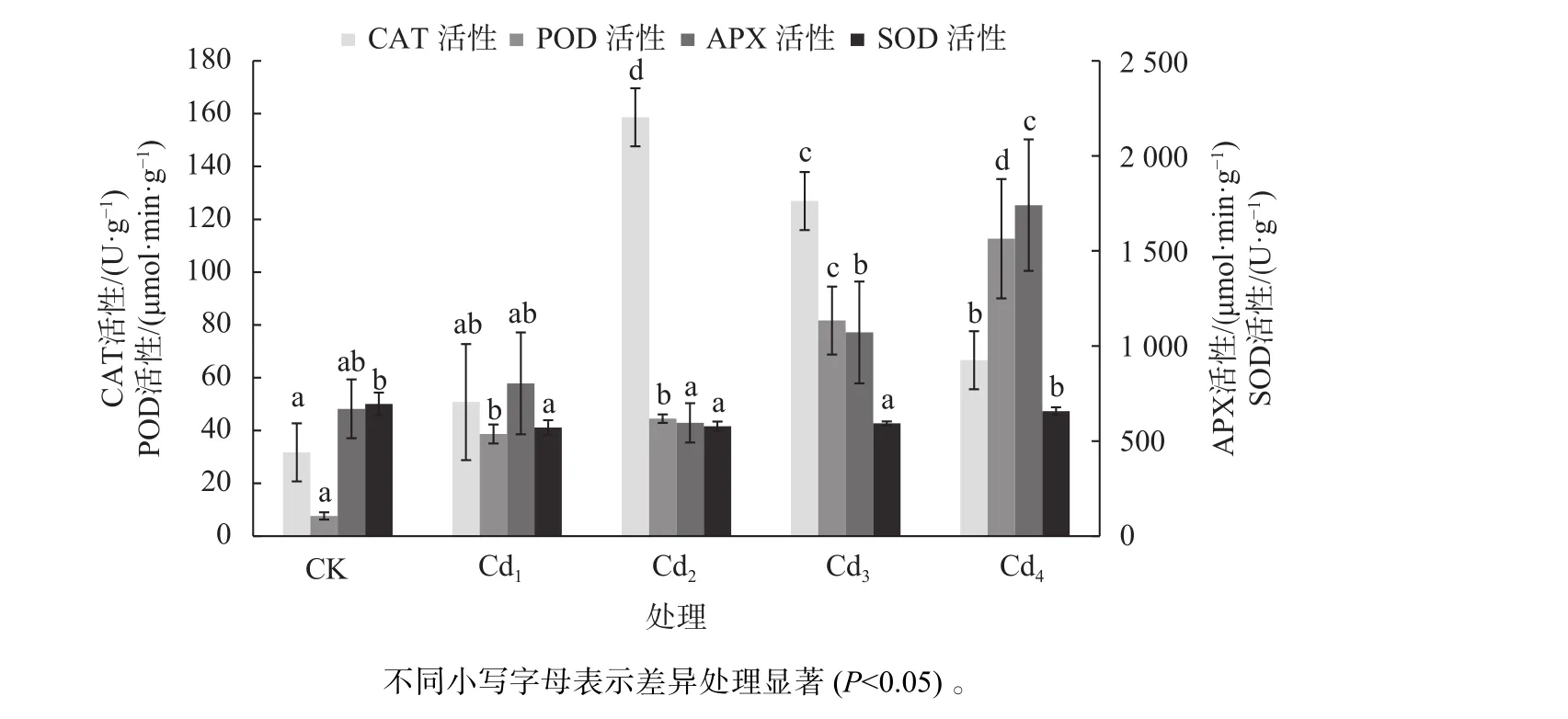
图2 不同镉浓度对洋常春藤抗氧化酶活性的影响Fig. 2 Effects of different Cd2+ concentrations on antioxidant enzymes activities of H.helix
3 结论与讨论
镉胁迫下的植物体通常具有植株矮小、生物量减少、渗透调节失衡和保护酶活性下降等特点[27-28]。植株生长和生物量的变化是镉胁迫下的最易观察的表观性状。本试验中,在20 μmol/L和80 μmol/L Cd2+胁迫下,Cd2+对洋常春藤茎、叶、总生物量影响微弱,并略微促进根系生物量的累积和根系增长;在 200 μmol/L 和 400 μmol/L Cd2+胁迫下,洋常春藤各生物量都减少,尤其是叶片数明显减少,叶生物量显著下降;说明高Cd2+会阻碍其正常的生长。李冬琴等[29]的研究中,耐镉性弱的玉豆(Phaseolus vulgaris),其株高和生物量随着Cd2+浓度的增加而显著下降;施宠等[30]的研究中,低Cd2+能够促进野燕麦(Awena fatua)幼苗株高、叶长、根长、茎粗的增加以及生物量的积累,高Cd2+则表现出抑制作用,其适于修复Cd2+浓度小于8 mg/L的污染土壤;在刘柿良等[31]的研究中表明,白车轴草(Tritolium repens)对Cd2+污染具有较强耐性,较低Cd2+处理对白车轴草无明显影响,而较高Cd2+则显著抑制其生物量与养分积累量分配;这说明Cd2+胁迫对不同植物生长和生物累积量的影响不同,本试验中洋常春藤表现出一定的耐镉性,叶对的镉的敏感程度大于茎、根。
可溶性蛋白可降低细胞渗透势,能与游离的重金属离子结合形成金属结合蛋白,减轻重金属的毒害作用,且含量越高,其生理生化代谢活动就越旺盛[32-35]。脯氨酸具有强亲水性,可调节细胞渗透平衡,增强细胞结构的稳定性,阻止氧自由基的产生,对植物体中的酶具有保护作用[36-39]。在植物受到高Cd2+胁迫时会通过增加渗透调节物质的含量来提高细胞渗透势以减轻受损程度。在杨叶萍等[40]的研究中160 mg/L Cd2+处理苎麻幼苗49 d,可诱导苎麻(Boehmeria nivea)体内大量形成可溶性糖、可溶性蛋白、游离脯氨酸等,而陈晶等[41]的研究中发现,0.5 mg/L Cd2+处理会略降低玉米(Zea mays)幼苗叶片脯氨酸含量,有研究用25 μmol/L Cd2+处理2个箭筈豌豆(Vicia stativa)品种7 d,镉敏感箭筈豌豆品种ZM叶中脯氨酸的含量较CK显著上升;镉耐性品种L3叶中脯氨酸含量较CK增加不显著[42]。本试验中,低Cd2+胁迫下,可溶性蛋白、脯氨酸含量增加不明显,在400 μmol/L Cd2+处理下两者的含量均极显著增加,说明洋常春藤叶片对低Cd2+胁迫不敏感,在高Cd2+胁迫下,则引起叶片内的渗透调节物质积极响应。Cd2+胁迫程度越大,其叶片在一定程度下会通过增加大量的可溶性蛋白、脯氨酸来平衡细胞内的渗透势,缓减损伤,表现出一定的抗镉性。
植物在逆境下可通过调节抗氧化酶的活性来缓解或消除破坏膜系统结构的膜脂过氧化物。其中SOD、POD、CAT能够歧化成H2O2和O2,APX则将H2O2分解成H2O和O2;4种酶协调作用清除细胞内的活性氧自由基、过氧化物[43-44],防止细胞内过量的自由基对细胞质膜和细胞内生物大分子的破坏,避免细胞内大量积累的过氧化物损伤细胞[45]。本试验中,洋常春藤中各抗氧化酶的活性随着处理浓度的增大而出现不同的变化,但大体呈上升趋势,有研究发现,野燕麦(Awena fatua)具有一定的耐镉能力,随着Cd2+浓度的增加,其幼苗体内的POD、APX、CAT活性也相应增加了;SOD活性则表现出“低促高抑”的现象[30],何梨香等[46]研究报道,在低浓度下圆叶决明(Chamaecrista rotundifolia)幼苗叶片内SOD活性虽有所降低,但由于其POD和CAT被激活,其幼苗的生长发育未受显著影响。有报道抗镉性差的土沉香(Aquilaria sinensis)幼苗,随着Cd2+胁迫浓度的增加,其叶片中SOD和CAT活性均出现不同程度的下降[47],表明镉胁迫下植株各抗氧化酶的具体调节是复杂多变的,本试验中的洋常春藤能够通过有效协调抗氧化酶活性来减缓低Cd2+胁迫下氧自由基和过氧化产物对细胞的损伤。
本试验中,低Cd2+胁迫时,洋常春藤的生长发育未受显著影响,大多数生长指标与CK的差异不显著;虽然MDA含量升高,SOD酶活性值低,但由于可溶性蛋白和脯氨酸的含量升高且POD、CAT和APX酶被诱导激活,缓解了镉胁迫对其的毒害作用。高Cd2+胁迫时,特别是400 μmol/L Cd2+胁迫下,洋常春藤的叶、茎生物量受到显著影响,镉毒害作用加强,各渗透调节物质和抗氧化酶虽有不同程度的显著增加,但不足以消除胁迫损伤,导致其正常的生长发育受到阻碍,生物量累积减少。总之,低Cd2+胁迫会引起洋常春藤细胞发生氧化毒害作用,叶片中的和MDA没有显著积累,此时,渗透调节物质没有显著增加,主要通过刺激POD和CAT活性显著上调减缓氧化损伤。400 μmol/L Cd2+胁迫下,洋常春藤叶片氧化毒害及渗透受损明显,叶片中显著增加大量和MDA,POD、CAT和APX活性显著上调,且渗透调节物质脯氨酸和可溶性蛋白也显著增加减缓氧化损伤,但其叶片数及生物量急剧减少,未能很好的减缓镉毒害。综上所述,洋常春藤具有一定的耐镉能力,能够通过调节植物细胞内可溶性蛋白、脯氨酸含量以及CAT、POD和APX活性等来抵抗低Cd2+胁迫,可作为低镉污染土壤地区园林绿化植物及生态修复植物推广应用。
