酵母菌发酵对常用饲料原料营养指标和抗营养因子含量的影响
2019-01-03谢鹏袁园葛莹钟佳祎蔡羽娟
谢鹏 袁园 葛莹 钟佳祎 蔡羽娟


摘要:生物发酵饲料是目前动物营养研究的热点领域,具有广阔的应用前景。比较研究了5种常用饲料原料(小麦、玉米、豌豆、大豆和豆粕)经酿酒酵母、热带假丝酵母和马克斯克鲁维酵母发酵后营养物质和抗营养因子含量的变化规律。结果表明,小麦和玉米经3种酵母发酵后pH值显著降低。豌豆、豆粕和大豆在发酵后期pH值均升高。除豆粕经热带假丝酵母发酵后干物质回收率上升外,其他原料经酵母菌发酵后干物质量均降低。玉米原料分别经酿酒酵母和马克斯克鲁维酵母发酵后还原糖含量显著下降,而大豆、豌豆和豆粕原料经2种酵母发酵后还原糖含量均表现出先升高后降低的规律。热带假丝酵母发酵小麦、玉米、大豆和豆粕4种原料粗蛋白含量均显著升高。3种酵母菌能够有效降低饲料原料中的植酸含量,但仅对小麦和玉米中的抗性淀粉有一定的降解效果。综上所述,通过控制酵母菌发酵时间,能够有效提高饲料原料的营养价值并降低其中的抗营养因子含量。利用酵母菌对饲料原料进行预发酵处理成为提高饲料中营养物质消化吸收利用率的有效途径。
关键词:酿酒酵母;热带假丝酵母;马克斯克鲁维酵母;发酵;饲料;营养指标;抗营养因子
中图分类号: S816.6文献标志码: A
文章编号:1002-1302(2019)21-0241-06
收稿日期:2018-08-02
基金项目:江苏省淮安市科技计划项目——重点研发计划(现代农业)(编号:HAN201605)。
作者简介:谢鹏(1986—),男,江苏淮安人,博士,副教授,主要从事动物营养与生理学研究。E-mail:pengxiejqs@126.com。
随着全球经济的蓬勃发展和人们生活水平的不断提高,人类对于畜产品的需求量逐年增大,从而促使畜牧业在规模与质量上不断发展。在全球人口日益增多的背景下,如何利用生物技术方法开发新型饲料资源并提高动物对原有饲料原料利用率成为解决不断增长的畜产品需求量和日益紧缩的耕地面积之间矛盾的有效途径之一。
研究表明,饲料经益生菌发酵后,菌种产生的多种代谢产物不仅能够提高饲料原料中蛋白质、脂肪和糖类等成分的营养转化率,同时能够降解常用饲料原料中的抗营养因子等有害成分,扩大饲料原料的使用范围,提高原有饲料的营养价值[1-3]。另外,发酵饲料能够改善饲料适口性,调节动物肠道的微生态平衡,提高了畜禽产品的食品安全性[4-5]。目前,应用于微生物发酵饲料的菌种类型主要有酵母菌、乳酸菌、芽孢杆菌、曲霉等。其中,酵母菌是单细胞真核微生物的一种,能够发酵糖类物质,其本身含有丰富的氨基酸、维生素、酶类等物质,是直接食用最多的一种微生物,在畜牧业生产中已有广泛的应用。本研究分别采用酿酒酵母、热带假丝酵母和马克思克鲁维酵母对5种常用饲料原料小麦、玉米、豌豆、大豆和豆粕进行发酵处理,對发酵前后的营养指标和抗营养因子含量进行比较测定,为发酵原料在饲料工业中的应用提供理论基础和技术支撑。
1材料与方法
1.1试验材料
1.1.1菌种
热带假丝酵母(Candida tropicalis)和酿酒酵母(Saccharomyces cerevisiae)均购自中国工业微生物菌种保藏管理中心。马克思克鲁维酵母(Kluyveromyces marxianus)由淮阴师范学院生命科学学院张瞳老师馈赠。
1.1.2原料及处理
发酵原料为市售小麦、玉米、豌豆、大豆和豆粕。将原料除杂后进行粉碎,过40目筛,作为酵母菌发酵基础料。
1.1.3培养基配制
1号试剂:酵母膏10 g,蛋白胨20 g,琼脂粉20 g(液体培养基不加),蒸馏水900 mL;2号试剂:葡萄糖20 g,蒸馏水100 mL。将1号试剂瓶和2号试剂瓶分别配制灭菌,待温度冷却至60~70 ℃后混合均匀。
1.1.4试验地点及时间
试验地点为淮阴师范学院生命科学学院,试验时间为2018年3—5月。
1.2试验方法
1.2.1菌的活化及扩大培养
将酵母菌冻干粉瓶打碎后,用液体培养基溶解,吸取100 μL接种至固体培养基表面,均匀涂布,30 ℃培养24~48 h后,选择菌落生长良好的培养皿,相同条件下进行划线培养,挑取单菌落,装入液体培养基中震荡培养,菌悬液混合均匀后按照1 ∶50接种量扩大培养,测定菌液D值,制备接种菌液。
1.2.2发酵
每种原料称取200 g,110 ℃加热7 min,各种酵母的接种量均为1.0×107 cfu/g,原料与水分比例为1 ∶1。混合均匀后,装入发酵袋中排空空气静置发酵,发酵温度为30 ℃,发酵时间为24、48、72、96 h。每隔12 h检查1次将袋中气体排空,发酵完毕后样品置于-20 ℃冰箱里待测。
1.3指标测定
1.3.1pH值的测定
取每株菌发酵的各时间段的每种发酵原料样品5 g于小烧杯中,加入45 mL蒸馏水,每隔5 min搅拌1次,共搅拌3次,静置10 min后,pH计测量。
1.3.2干物质回收率的测定
干物质测定方法参照GB/T 8303—2013《茶磨碎试样的制备及其干物质含量测定》,计算公式:干物质回收率=(发酵原料干物质质量/发酵原料质量)/(原料干物质质量/原料质量)×100%。
1.3.3还原糖含量的测定
参照王俊刚等的3,5-二硝基水杨酸法测定[6]。准确称取2.0 g样品于100 mL三角瓶中,加入约50 mL蒸馏水,于50 ℃恒温水浴中保温20 min,期间不时搅拌使还原糖浸出。静止三角瓶使样品沉淀,吸取上清液8 mL于离心管中,3 000 r/min离心10 min,将上清液转移至新离心管中,此即为还原糖测定样品。还原糖含量以mg/g计算。
1.3.4粗蛋白含量的测定
依据GB/T 6432—1994《饲料中粗蛋白测定方法》凯式定氮法进行测定。称取0.3 g左右测定干物质时烘干至恒重的样品,加入催化剂和硫酸,进行消化,直至溶液透明呈蓝绿色,待其冷却,应用自动凯氏定氮仪测定其含氮量。
1.3.5植酸含量测定方法
采用分光光度法[7]测定植酸含量。步骤大致如下:(1)称取0.5 g样品,加入6 mL1.8 mol/L HCl,35 ℃水浴2 h,期间不断振荡,水浴后静置过夜;(2)样品4 000 r/min离心15 min,取上清液,加入0.4 mL铁试剂,沸水浴45 min后再次离心,弃上清液;(3)用3 mL1.5 mol/L NaOH对沉淀进行溶解,再次离心,弃上清液;(4)加入1 mL 3.2 mol/L硝酸溶解沉淀后,依次加入0.2 mL 9%的柠檬酸三钠、0.2 mL 1%对苯二酚、0.4 mL 0.4%邻菲罗啉,纯水定容至50 mL,pH值调节至3~4;(5)35 ℃水浴2 h后自然冷却,于510 nm波长处测定吸光度。
计算公式:植酸含量=D510 nm×8.42÷样品质量(mg)×125×100%。
1.3.6抗性淀粉含量的测定
参照高鑫(2012)的方法[8]。步骤大致如下:(1)准确称取0.2 g样品于带塞玻璃试管中,加入蒸馏水0.4 mL,沸水浴20 min后冷却至室温;(2)加入HCl-KCl缓冲溶液(pH值为2.0)2.5 mL和10%胃蛋白酶105 μL,37 ℃恒温振荡1 h;(3)加入4 mL的醋酸钠缓冲液(pH值=6.9)和淀粉酶-糖化酶混合溶液4 mL,37 ℃振荡16 h;(4)4 000 r/min离心15 min,去掉上清液,50%乙醇溶液洗涤沉淀,重复2次;(5)依次加入2 mL 2 mol/L KOH溶液,8 mL 醋酸钠缓冲液(pH值为3.8),100 μL糖化酶溶液,60 ℃ 水浴1 h;(6)冷却至室温,4 000 r/min离心15 min,保留上清液;(7)根据“1.3.3”节还原糖测定方法测定上清液中的还原糖含量。
计算公式:抗性淀粉含量=还原糖含量×0.9。
1.4数据处理
统计学分析采用SPSS 17.0(SPSS Inc.,Chicago,IL)软件的One-Way ANOVA进行统计分析,计算结果以“平均值±标准误”表示。P<0.05表示差异显著。
2结果与分析
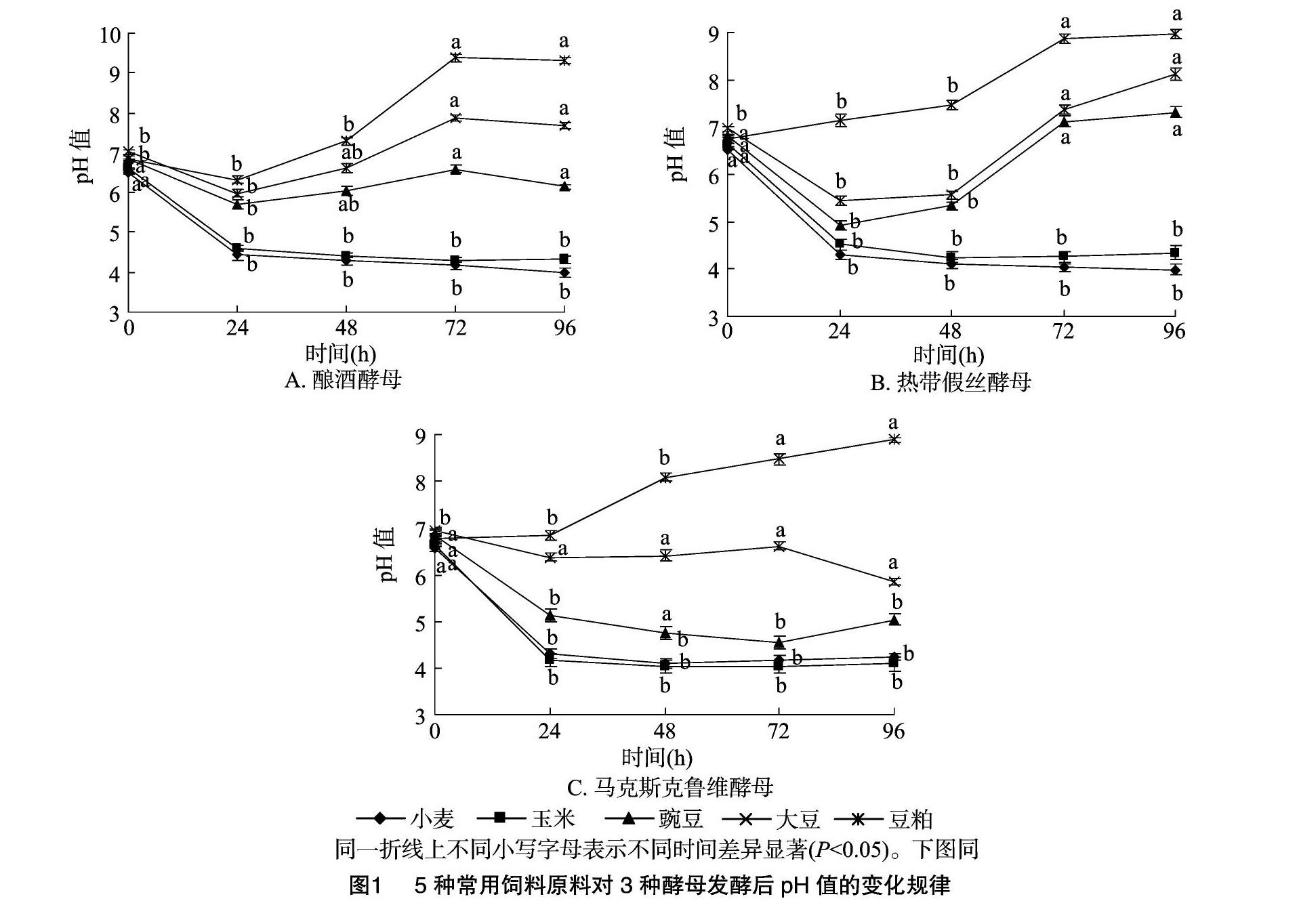
2.1不同饲料原料经3种酵母菌发酵后pH值的变化规律
由图1可知,小麦和玉米分别经3种酵母发酵24 h后pH值均显著降低(P<0.05),且于48~96 h内趋于稳定水平,呈明顯的酸性。豌豆和豆粕分别经酿酒酵母和热带假丝酵母发酵24 h后pH值均显著降低(P<0.05),但随后于72 h后逐渐恢复至初始pH值。大豆原料经3种酵母发酵72 h后pH值均显著升高(P<0.05),呈明显碱性。
2.2不同饲料原料经3种酵母菌发酵后干物质回收率的变化规律
由图2可知,小麦原料经3种酵母发酵72 h后干物质回收率显著降低(P<0.05),玉米原料经酿酒酵母发酵后干物质回收率呈现先降低后升高的趋势,而经热带假丝酵母和马克斯克鲁维酵母发酵72 h和96 h后干物质回收率则显著降低(P<0.05)。豌豆经酿酒酵母和热带假丝酵母发酵96 h后干物质回收率显著降低,但不受马克斯克鲁维酵母发酵影响(P>0.05)。大豆分别经热带假丝酵母和马克斯克鲁维酵母发酵72 h和48 h后干物质回收率显著降低(P<0.05),豆粕原料分别经酿酒酵母和马克斯克鲁维酵母发酵72 h后干物质回收率显著降低(P<0.05),而经热带假丝酵母发酵72 h后干物质回收率却显著上升(P<0.05)。
2.3不同饲料原料经3种酵母菌发酵后还原糖含量的变化规律
由图3可知,玉米原料分别经酿酒酵母和马克斯克鲁维酵母发酵24 h后还原糖含量显著下降(P<0.05),而大豆、豌豆和豆粕原料经2种酵母发酵后还原糖含量均表现出先升高后降低的规律。豌豆原料经热带假丝酵母发酵24 h后还原糖含量显著降低(P<0.05),而其他原料均表现出先升高后降低的规律。
2.4不同饲料原料经3种酵母菌发酵后粗蛋白含量的变化规律
由图4可知,酿酒酵母发酵豆粕原料96 h时粗蛋白含量显著降低(P<0.05),而发酵其他4种饲料原料对蛋白含量无显著影响(P>0.05)。热带假丝酵母发酵小麦、玉米、大豆和豆粕4种原料72~96 h内粗蛋白含量均出现显著升高现象(P<0.05)。马克斯克鲁维酵母发酵玉米24 h时,粗蛋白含量显著升高(P<0.05),但随后降低至初始水平,而发酵豆粕原料72 h后粗蛋白含量显著升高(P<0.05)。
2.5不同饲料原料经3种酵母菌发酵后植酸含量的变化规律
由图5可知,酿酒酵母发酵24 h后,大豆、豆粕、小麦植酸含量显著降低(P<0.05);豌豆发酵72 h后,植酸含量的去除率达到49.3%(P<0.05);玉米的植酸含量在发酵48 h内无显著变化(P>0.05)。热带假丝酵母发酵24 h时,大豆、豌豆、小麦植酸含量显著降低(P<0.05);发酵48 h之后豆粕植酸含量的去除率达到55.3%(P<0.05),玉米的植酸含量无显著变化(P>0.05)。马克思克鲁维酵母发酵24 h后,大豆、豆粕、小麦植酸含量显著降低(P<0.05),玉米与豌豆发酵48 h 后植酸含量显著降低(P<0.05)。马克思克鲁维酵母对大豆植酸含量降低效果最为显著,对其植酸去除率达到68.1%。
2.6不同饲料原料经3种酵母菌发酵后抗性淀粉含量的变化规律
由图6可知,酿酒酵母发酵48 h后,小麦原料的抗性淀粉含量显著降低(P<0.05),而豌豆和玉米则在发酵72 h抗性淀粉含量显著降低(P<0.05)。豌豆和小麦经热带假丝酵母发酵24 h后抗性淀粉含量显著降低(P<0.05)。马克思克鲁维酵母发酵48 h后豌豆抗性淀粉含量降低44%(P<005);发酵72 h后,小麦与玉米抗性淀粉含量分别降低了43%和52%(P<0.05);大豆和豆粕则在发酵过程中抗性淀粉含量无显著变化(P>0.05)。
3讨论
pH值是生物發酵工艺中的一个非常重要的参数,饲料原料发酵过程中pH值降低主要归结于发酵菌种增殖过程中产生并积累的有机酸[9],微生物发酵饲料中酸含量的增加反过来有利于饲料的长期保存[10]。本试验中,小麦和玉米原料经酿酒酵母、热带假丝酵母和马克斯克鲁维酵母发酵后pH值均呈现显著下降趋势。研究发现,酵母菌本身产酸能力并不强,在发酵过程中可将葡萄糖代谢产物——丙酮酸转化为乳酸、乙酸或乙醇[11],这可能是诱发原料pH值在发酵初期降低的因素之一。随着发酵后期乙醇的积累,酵母细胞的生长受到抑制,丙酮酸代谢产酸途径被切断[12],因此pH值无法持续降低。也有研究人员认为,原料本身未被灭菌的乳酸菌在发酵过程中得到一定程度的增殖,且酵母菌增殖过程中产生的丙酮酸、琥珀酸等代谢产物能够刺激乳酸菌的活动[13],为其提供营养物质,因而诱导原料pH值的迅速降低[14]。本试验还发现大豆在酵母菌发酵后期pH值逐渐升高,说明蛋白类原料和淀粉类原料被酵母菌发酵的效果存在较大差异。作者推测这种差异可能归结于酵母菌在不同发酵底物下积累的代谢产物不同所导致,在本试验发酵过程中发现,大豆发酵后期出现腥味,可能是挥发性氨类和硫化物。刘倚帆等报道,酵母菌和米曲霉协同发酵整粒菜籽后期pH值也出现一定程度的升高[15]。因此,全脂大豆或菜籽中较高含量的蛋白质以及脂肪可能是酵母菌发酵后期pH升高的主要因素。本研究中豆粕和豌豆在3种不同酵母菌发酵下pH值呈现不同的变化规律也说明不同酵母菌在单一发酵底物下的代谢方式和途径存在一定差异。
Terebiznik等认为益生菌固态发酵过程中干物质损失量和生物量呈正相关[16]。这一点在马克斯克鲁维酵母发酵中尤为明显,酵母菌分解利用发酵底物中的物质合成自身成分,并释放出代谢产物,在这一过程中,干物质量在发酵后期呈现明显的下降趋势。但本试验也发现,热带假丝酵母发酵豆粕后期干物质回收率出现升高趋势 说明发酵物料中的生物量增长速度超过了发酵底料的消耗速度,笔者推测这种现象可能只是短暂出现,随着发酵的继续进行,发酵底物的消耗速度势必会超过生物量的增长。
酵母菌最重要的营养物质和能量来源大部分来自于单糖,而发酵纤维素等多糖的能力有限,这一点与枯草芽孢杆菌不同。大量研究报道,在果酒、泡菜等酿造发酵过程中,酵母菌能够迅速降解底物中已有的还原糖成分,将其转化为酸类、酯类或醇类物质[17-19]。本研究结果发现,玉米分别经酿酒酵母和马克斯克鲁维酵母发酵后还原糖含量持续下降,而大豆、豌豆和豆粕原料经2种酵母发酵后还原糖含量均表现出先升高后降低的规律。豌豆经热带假丝酵母发酵后还原糖含量持续降低,而其他原料均表现出先升高后降低的规律。由此说明,不同酵母菌作用于不同饲料原料底物,产生能够分解多糖的酶的性质不同,后期当淀粉等多糖经酵母菌酶解成为还原糖后反过来又促进了酵母菌的增殖。
本研究结果表明,热带假丝酵母发酵小麦、玉米、大豆和豆粕4种原料可提高原料的粗蛋白水平,这一点优于酿酒酵母和马克斯克鲁维酵母。研究报道,生产上利用淀粉、葡萄糖、玉米深加工副产物等作为热带假丝酵母、酿酒酵母大量培养的底物,酵母本身蛋白质含量为45%~60%,其大量增殖的同时提高了发酵底物的粗蛋白含量[20-22],从而使底物的可饲性大大提高,可最大限度上替代鱼粉等其他动物源蛋白质饲料。本研究中单一饲料原料的成分及含量均有所不同,可能是造成不同酵母增殖效果差异的重要因素。
植酸在植物种子的胚芽和谷物的麸皮中含量较高,尤其是在豆科植物的种子中含量最高。植酸可与常见的矿物元素产生不溶性化合物,大大降低其有效性,因而成为重要的抗营养因子之一。本试验测定结果显示,大豆和豌豆中植酸含量较高,但豆粕中植酸含量甚至低于玉米和小麦。这可能与大豆在压榨获得油脂过程中经历高温高压等物理因素有关。研究报道,酵母菌能够有效降解植物籽实中的植酸[23],因而有利于单胃动物对饲料中营养物质的吸收和利用,这可能与酵母菌能够产植酸酶有直接关系[24-25]。本试验中马克思克鲁维酵母在发酵48 h后各原料的植酸含量下降均较为明显,显示出较好的去除效果,因而在实际生产上具有一定的借鉴意义。
抗性淀粉是一种细胞壁成分,部分与水相溶形成粘性物质,阻碍了动物对营养物质的吸收。本试验中酵母菌对玉米、小麦和豌豆等抗性淀粉含量较高的原料降解效果较好,而对大豆和豆粕的抗性淀粉降解效果较差。目前利用微生物发酵法降解植物籽实中的抗性淀粉尚未见报道。笔者认为不同原料中抗性淀粉的类型存在差异,酵母菌在不同底物上的发酵效果也有所不同,分解抗性淀粉可能主要靠分泌的纤维素酶或淀粉酶发挥作用,但由于酵母菌对复杂多糖的利用效果有限,因此仅依靠酵母菌发酵难以达到有效降解饲料中抗性淀粉的目的。
综上所述,不同种类酵母菌对能量型和蛋白型饲料原料的发酵效果存在差异。总体上,经过3种酵母菌发酵后的能量型饲料(小麦和玉米)酸度显著增加,而蛋白型饲料(豌豆、大豆和豆粕)经酿酒酵母和热带假丝酵母发酵后期pH值升高。通过控制菌种和时间可在发酵早期增加原料中还原糖和粗蛋白的含量,但随着发酵的进行,还原糖逐渐被消耗。此外,3种酵母菌能够有效降解饲料原料中的植酸,但仅对能量型饲料的抗性淀粉有一定的降解效果。
参考文献:
[1]张宇红. 不同复合菌比例发酵饲料对饲料营养价值及生长猪生产性能的影响[D]. 雅安:四川农业大学,2004.
[2]李龙. 复合益生菌发酵饲料工艺参数优化及品质评定[D]. 上海:上海交通大学,2010.
[3]刘艳新,刘占英,倪慧娟,等. 微生物发酵饲料的研究进展与前景展望[J]. 饲料博览,2017(2):15-22.
[4]胡新旭,周映华,刘惠知,等. 无抗发酵饲料对断奶仔猪生长性能、肠道菌群、血液生化指标和免疫性能的影响[J]. 動物营养学报,2013,25(12):2989-2997.
[5]王长彦. 微生物发酵饲料替代饲用抗生素技术在商品猪生产中的应用研究[D]. 杨凌:西北农林科技大学,2008.
[6]王俊刚,张树珍,杨本鹏,等. 3,5-二硝基水杨酸(DNS)法测定甘蔗茎节总糖和还原糖含量[J]. 甘蔗糖业,2008(5):45-49.
[7]李桂荣. 燕麦籽粒植酸含量影响因素及生理机制研究[D]. 北京:中国农业大学,2007.
[8]高鑫. 山药块茎抗性淀粉形成的影响因素及理化特性的研究[D]. 呼和浩特:内蒙古农业大学,2012.
[9]刘晓明. 乳酸菌、枯草芽孢杆菌、酵母菌发酵不同饲料原料参数的研究[D]. 泰安:山东农业大学,2015.
[10]Muck R E. Factors influencing silage quality and their implications for management[J]. Journal of Dairy Science,1988,71(11):2992-3002.
[11]Hartikainen K,Poutanen K,Katina K. Influence of bioprocessed wheat bran on the physical and chemical properties of dough and on wheat bread texture[J]. Cereal Chemistry,2014,91(2):115-123.
[12]张秋美,赵心清,姜如娇,等. 酿酒酵母乙醇耐性的分子机制及基因工程改造[J]. 生物工程学报,2009,25(4):481-487.
[13]Leroi F,Pidoux M. Detection of interactions between yeasts and lacticacid bacteria isolated from sugary kefir grains[J]. Journal of Applied Bacteriology,1993,74(1):48-53.
[14]崔晨晓,朱科学,郭晓娜,等. 酵母菌发酵对小麦麸皮成分的影响研究[J]. 中国粮油学报,2016,31(7):25-29.
[15]刘倚帆,王星凌,岳寿松,等. 复合微生物固态发酵整粒菜籽的研究[J]. 中国饲料,2017(1):36-40,44.
[16]Terebiznik M R,Pilosof,R A M. Biomass estimation in solid state fermentation by modeling dry matter weight loss[J]. Biotechnology Techniques,1999,13(3):215-219.
[17]王嘉怡,宋艳,吕晓华. 泡菜和酒渣中酵母菌的分离及其发酵作用初探[J]. 中国调味品,2017,42(7):25-28,32.
[18]朱娟娟,郑少阳,李炎杰,等. 不同酿酒酵母对脐橙果酒发酵特性的影响[J]. 南方农业学报,2017,48(5):870-875.
[19]姜越,潘婷,惠竹梅. 模拟葡萄汁中可同化氮和还原糖对酵母发酵特性的影响[J]. 食品科学,2018,39(2):131-137.
[20]尹玮,温海琴,汪群慧,等. 玉米秸秆乳酸发酵残渣制取饲料的研究[J]. 黑龙江大学自然科学学报,2008,25(4):480-484.
[21]张敏,侯先志,考桂兰,等. 酵母菌发酵生产饲料酵母条件的优化[J]. 饲料工业,2010,31(8):28-30.
[22]宾冬梅,黄河清涛,易诚,等. 基于饲料酵母二次发酵啤酒糟条件响应面优化研究[J]. 湖南生态科学学报,2017,4(4):14-19.
[23]黄耀武,周建平. 发酵法降解油菜籽废水中植酸的工艺研究[J]. 现代食品科技,2012,28(1):66-68,73.
[24]王晨祥,张苓花. 克鲁斯假丝酵母植酸酶的筛选及其发酵条件[J]. 大连轻工业学院学报,2002,21(3):184-188.
[25]李晓宇,柳志强,田富,等. 响应面法优化热带假丝酵母ypy06植酸酶发酵条件的研究[J]. 热带农业科学,2011,31(6):41-44.
