细鳞鲑对水溶性氨基酸的嗅觉行为反应
2018-12-27蔡红英陈春山张旭光郭弘艺郭明磊
魏 凯,蔡红英,陈春山,张旭光,郭弘艺,李 超,马 龙,郭明磊
(1. 北京市水生野生动植物救护中心,北京 102100; 2. 上海海洋大学水产与生命学院,上海 201306;3. 北京市鲟鱼鲑鳟鱼创新团队,北京 102100; 4. 中国水产科学研究院东海水产研究所,上海 200090)
细鳞鲑(Brachymystaxlenok),隶属鲑形目(Salmoniformes),鲑科(Salmonidae),细鳞鲑属,多分布于西伯利亚、朝鲜半岛和中国的黑龙江、图们江、鸭绿江等水域,在秦岭和北京地区部分水系有零星分布,属冷水性珍贵鱼类。春季雨水引起河水上涨时,细鳞鲑由江河进入具有清水的支流中,一般生活在支流的山涧深潭中,秋季支流的水变浅时,由支流进入江河中越冬。随着不同年份洪水的情况,细鳞鲑向主河道洄游的时间略有不同。由于环境恶化和栖息地环境变化等原因,细鳞鲑的分布区域逐渐缩小,野生资源量也日益减少,对细鳞鲑野生种群的保护已迫在眉睫[1]。目前,国内对细鳞鲑繁殖生物学[1]、栖息地的地理环境因子[2]等展开了研究,但对其洄游行为的研究相对较少。鲑类洄游机制具有嗅觉印迹和性外激素诱导两种假设[3-5]。最近,有研究提出大马哈鱼(Oncorhynchusketa)可利用其出生地的单体氨基酸进行洄游的假设[6]。支流水体中化学成份的研究表明,其常见的水溶性单体氨基酸有谷氨酸(Glu)、天冬氨酸(Asp)、甘氨酸(Gly)、丙氨酸(Ala)、丝氨酸(Ser)等[7-10]。细鳞鲑由主河道进入支流的习性,尚不清楚是否为支流水体中主要单体氨基酸对其嗅觉行为的影响。本实验采用Y型迷宫,研究了4种主要水溶性氨基酸对细鳞鲑嗅觉行为的影响,以期一定程度阐明细鳞鲑的洄游行为机理。
1 材料与方法
1.1 实验材料
实验使用的细鳞鲑幼鱼来自吉林省延边细鳞鲑良种场,平均体质量(6.0±2.4)g,平均体长(10.0±1.8)cm。试验鱼暂养于冷水机循环系统的圆形帆布水槽(1 m×1.6 m)中,平均水温(13.0±1.5)℃,正常自然光照。暂养期间每日投喂一次人工颗粒饵料。实验前从暂养水槽随机取20尾健康的鱼,分别放入位于水槽内的隔离网笼进行分离暂养48 h,以适应封闭环境,减少实验应激反应。
1.2 实验方法
实验在自制的Y型迷宫水槽中(长80 cm×宽20 cm×高10 cm)进行,水槽的一端设有长30 cm的挡板,将水槽一端分成左右两个同样的选择区A、B(图1)。两种相同浓度的氨基酸溶液通过调节阀以相同的速度(2 mL·s-1)流入选择区。水槽另一端设有出水口,并通过水流阀控制氨基酸的扩散速度,调节氨基酸溶液扩散至选择臂末端的时间为10~12 min。实验时,试验鱼由活动网隔离在选择区前方(图1),适应15 min后移去活动网,同时打开氨基酸刺激液的调节阀,并同步视频记录试验鱼对不同选择区的行为偏好反应。为了避免两种氨基酸在挡板末端混和,实验记录时间设置为10 min,并离线分析试验鱼的行为反应。
分别测试了20尾细鳞幼鱼对甘氨酸(Gly)、谷氨酸(Glu)、丙氨酸(Ala)、天冬氨酸(Asp)的选择偏好性。前期预实验表明,当氨基酸母溶液配制浓度为10-3摩尔每升(M)时,实验水槽内的氨基酸刺激液可对细鳞幼鱼有较好的吸引效应。经计算,氨基酸母溶液扩散至挡板末端时实验水槽内实际浓度约为2.5×10-6M。为了比较试验鱼对上述氨基酸的偏好性,随机选择以甘氨酸为参照进行两两比较分析,分别进行了Gly vs Glu、Gly vs Ala、Gly vs Asp 3组实验。实验用水与暂养水同,以减少试验鱼的应激反应。
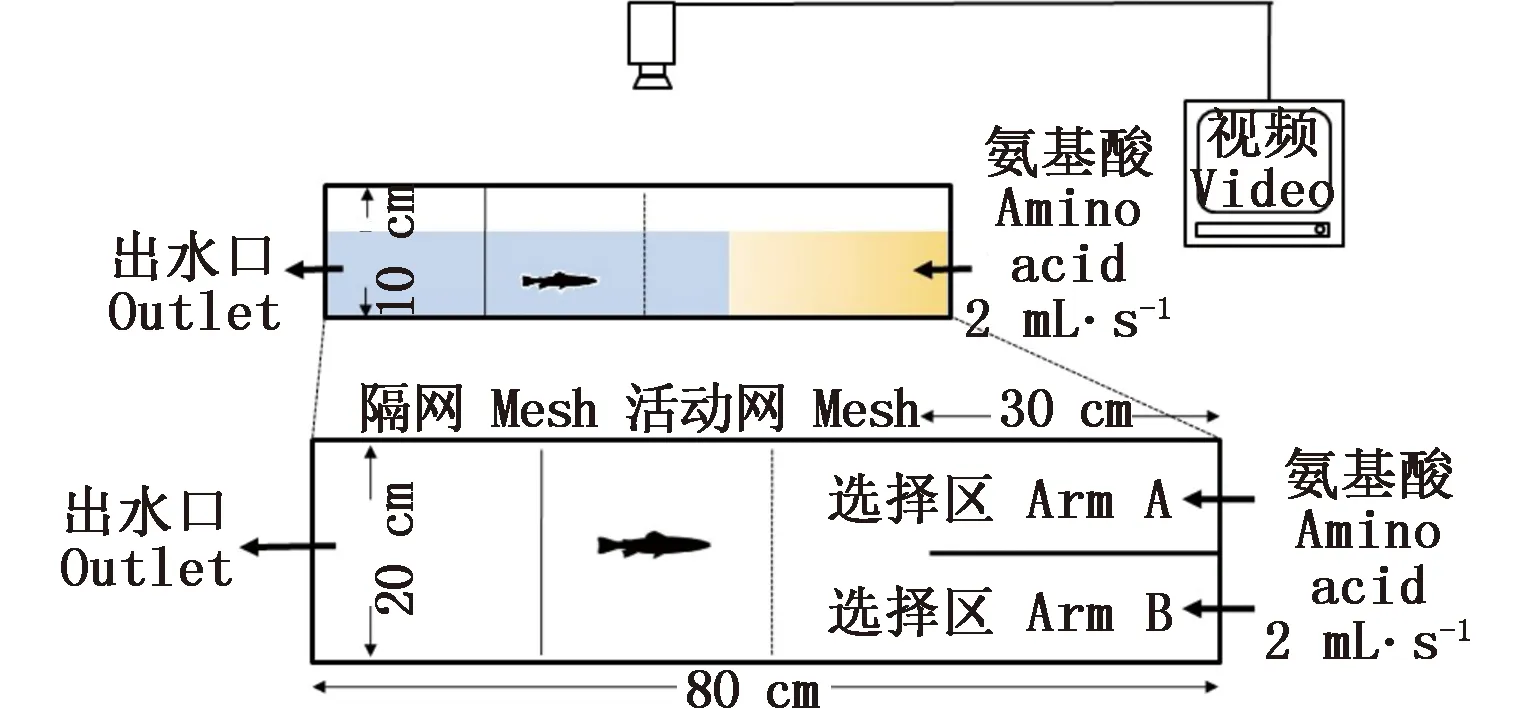
图1 实验装置示意图Fig.1 Schematic diagram of the Y-maze flow-through system
1.3 数据分析
实验中,细鳞鲑幼鱼对某一种氨基酸的行为选择偏好性,主要因素为氨基酸种类。试验鱼选择偏好性主要由试验鱼是否进入选择区,以及在选择区的停留时间决定。依据ZHANG等[11]、张旭光[ 12]研究,试验鱼对某一氨基酸的绝对偏好度(preference degree)定义为P=(N×Ta)/Tl,式中:P为绝对偏好度;N为进入频次;Ta为进入选择区的平均停留时间(activity time);Tl为第一次进入选择区的反应时间或延迟时间(latency time)。该公式表明,绝对偏好度与试验鱼在选择区的停留时间成正比,停留时间越长,选择偏好性越大;与试验鱼第一次趋向选择某种氨基酸的延迟时间成反比,延迟时间越长表明偏好性越小。
为了确定试验鱼群体对4种氨基酸的偏好性差异,将甘氨酸(Gly)的绝对偏好度标准化为1,根据公式Rx=Px/PGly进行标准化转化计算对应的氨基酸的相对偏好有效值[13],其中Rx为氨基酸的相对偏好有效值,Px为氨基酸的绝对偏好度,PGly为甘氨酸的绝对偏好度。根据相对偏好有效值的大小来确定细鳞鲑幼鱼对不同氨基酸的行为选择偏好性大小顺序。所有数据均以平均值±标准误(Mean±SEM)来表示,由Sigmaplot(Version 12.5)进行绘制图表和显著性分析。
2 结果与分析
2.1 嗅觉行为反应
在实验记录时间内,大多数细鳞鲑幼鱼(n=56,93%)都表现出对单体氨基酸的正趋向反应,但也有个别幼鱼(n=4)在记录结束才表现出趋向反应。在经过一定反应延迟时间后能一次或数次进入前方的选择区,反应延迟时间为289~398 s,平均时间为(329±14)s,该时间可能与氨基酸的扩散速度以及试验鱼对氨基酸嗅觉敏感性有关。细鳞鲑幼鱼在选择区通常停留一段时间,为12~400 s,平均停留时间为(90±11)s。实验发现细鳞鲑幼鱼(71%)会同时表现出对两种氨基酸的偏好,它们往往选择进入一侧氨基酸区后也选择进入另一侧选择区,但实验中也发现有的试验鱼(29%)只选择在一侧选择区停留。
2.2 偏好度分析
在3组两两对比实验中,试验鱼对参照组甘氨酸(Gly)的平均偏好度分别为(0.50±0.16)、(0.42±0.11)和(0.58±0.12),3组间试验鱼甘氨酸(Gly)的偏好度无显著性差异(P>0.05)(图2)。在Gly vs Ala比较组中,试验鱼对Gly的平均选择偏好度(0.50±0.16)略小于Ala (0.60±0.13),但二者差异不显著(P>0.05);但在Gly vs Glu组,试验鱼对Glu的平均偏好度(0.82±0.19)大于Gly(0.42±0.11),且二者差异达显著性水平(P<0.05);在Gly vs Asp组,试验鱼对Asp的平均偏好度(0.70±0.15)略大于Gly(0.58±0.12),但二者未达显著性差异水平(P>0.05)。
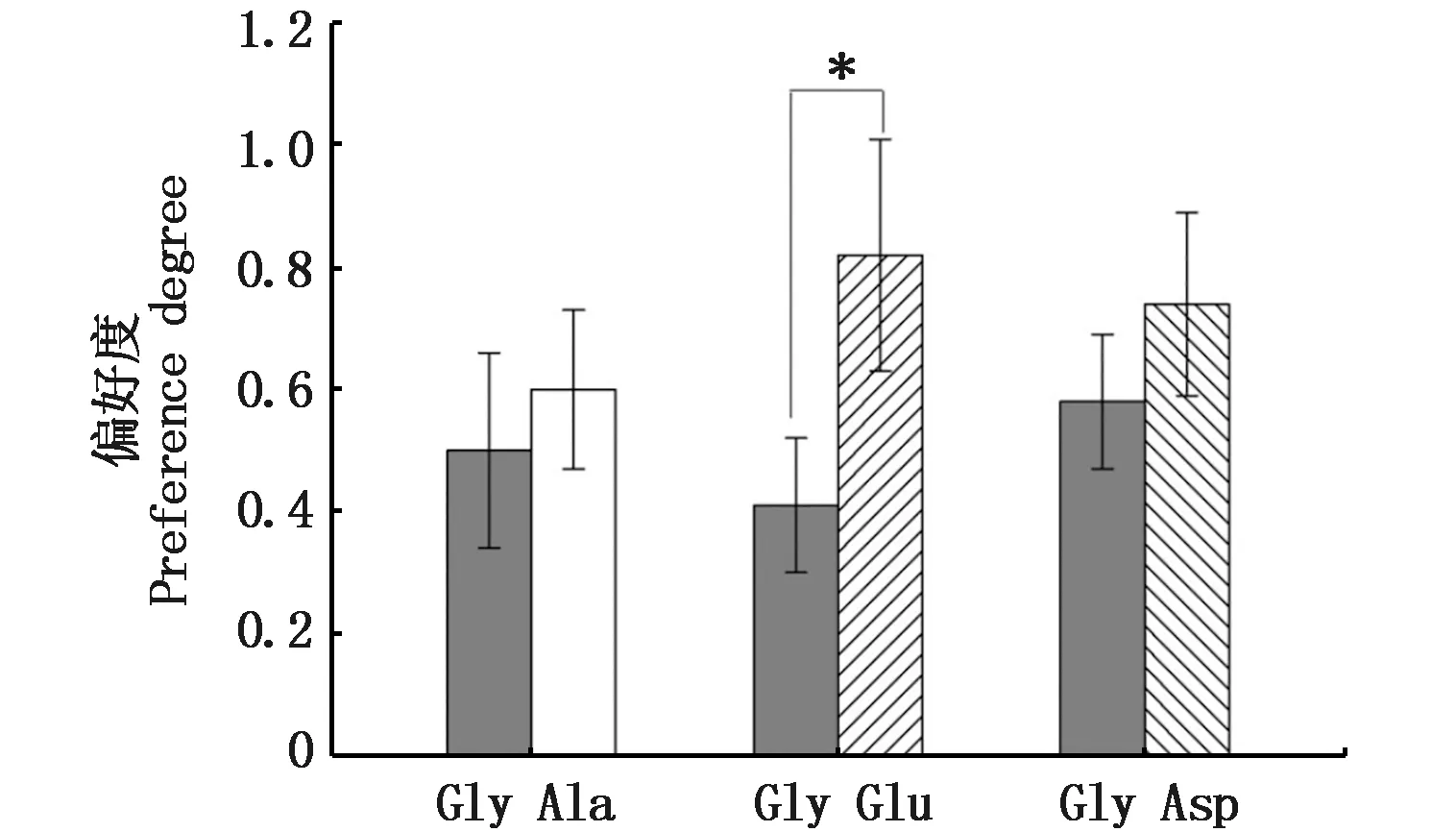
图2 细鳞鲑幼鱼对氨基酸的偏好度Fig.2 Preference degree to amino acids in Brachymystax lenok注:* 表示差异显著(P<0.05)Note: * means significant differences (P<0.05)
根据Rx=Px/PGly计算,细鳞鲑幼鱼对4种氨基酸的相对偏好度有效值分别为1(Gly)、1.18(Ala)、1.98(Glu)、1.27(Asp)。依据相对偏好度有效值大小,细鳞鲑对氨基酸的偏好性顺序为谷氨酸(Glu)>天冬氨酸(Asp)>丙氨酸(Ala)>甘氨酸(Gly)。
3 讨论
本实验通过行为学的方法研究了细鳞鲑幼鱼对谷氨酸(Glu)、甘氨酸(Gly)、天冬氨酸(Asp)、丙氨酸(Ala)4种单体氨基酸的偏好反应。试验鱼对氨基酸的平均反应延迟时间为300 s以上,表明试验鱼对氨基酸的反应存在刺激阈值浓度。假设氨基酸母溶液以匀速扩散,那么引起试验鱼进入选择区的实际氨基酸浓度约为1×10-6M,与引起鱼类的嗅觉行为反应的刺激阈值浓度相一致[14]。而高于阈值浓度的刺激则会引起鱼类的逃避或抑制反应[15]。这也可以解释,试验鱼在进入选择区后通常停留一段时间后离开的现象。
根据相对偏好度的有效值,细鳞鲑幼鱼嗅觉反应最敏感的为谷氨酸,其次为天冬氨酸、丙氨酸和甘氨酸。这可能与细鳞鲑嗅觉对氨基酸的生理敏感性有关。但对于上述4种氨基酸,虹鳟(Salmogairdneri)嗅觉电生理研究表明,其嗅觉上皮对丙氨酸(Ala)最敏感,其嗅电图反应的相对有效幅值较高,达108,而其它氨基酸的有效幅值较低(Gly:102; Glu:59.2; Asp:42)[16-17],虹鳟嗅觉上皮对氨基酸敏感性大小顺序为:丙氨酸(Ala)>甘氨酸(Gly)>谷氨酸(Glu)>天冬氨酸(Asp)。虽然现阶段没有基于氨基酸刺激的细鳞鲑嗅觉电生理研究资料,但基于已报道的虹鳟的嗅觉电生理研究资料和本实验数据,推测细鳞鲑对氨基酸的行为敏感性可能与其嗅觉上皮的氨基酸生理敏感性相匹配。这一假设还需要在后续的实验中开展细鳞鲑嗅电图反应的研究来验证。
嗅觉行为研究也证实了单体氨基酸可以有效地刺激鱼类的摄食行为[18]。例如,未开口的虹鳟和大西洋鲑幼鱼,经丙氨酸、甘氨酸、谷氨酸等单体氨基酸刺激,可诱发其摄食相关的行为反应,如咬合(snapping)、游泳加速(darting)等[19]。因此,鲑类对氨基酸的敏感性及其反应可能是一种内源性行为。单体氨基酸也可以诱发鲑类幼鱼形成嗅觉印迹[20]。嗅觉印迹是鱼类一种主要洄游机制,产生嗅觉印迹的因素还涉及各种性外激素[5, 21]或同类的气味[3, 6]。行为学和电生理的研究都显示单体氨基酸的嗅觉印迹是鲑类生殖洄游的重要机制[8, 20, 22-23]。Y型迷宫选择实验也表明,生殖期的马苏大马哈鱼(Oncorhynchusketa)对氨基酸种类和含量配比与其出生地相似的人工刺激液具有明显的选择偏好[22]。同时,该人工刺激液诱发了马苏大马哈鱼幅值较高的嗅电图电位,而对无机盐或胆酸配比相似的人工刺激液的反应则较小[8]。这些研究都表明鲑类对氨基酸的敏感性在其洄游行为中起着重要的线索作用[8, 22-23]。
在对马苏大马哈鱼的栖息地天盐川河(Teshio River)中水溶性氨基酸成分和含量进行的5年(2006-2010年)跟踪调查表明,在自然条件下,氨其酸的成分是基本稳定的,7种氨基酸丙氨酸、天冬氨酸、谷氨酸、甘氨酸、亮氨酸、丝氨酸和缬氨酸在5年内变化差异较小,其中丙氨酸和天冬氨酸含量最高[8]。同样,对细鳞鲑栖息河流的水溶性氨基酸调查也表明,其氨基酸总量为4.2~13.8 μM,其中单体氨基含量20%左右,约为0.84~2.76 μM[7]。本实验中,氨基酸的实际效应浓度为1.00~2.58 μM,与自然水域的氨基酸浓度相接近。细鳞鲑对谷氨酸保持的显著敏感性,一方面由其嗅觉上皮的生理敏感性决定,另一方面与自然栖息水域中谷氨酸最高含量的特点相匹配。但同时,细鳞鲑也保持了对其它几种主要氨基酸的行为偏好性。这表明,不同流域或其支流的主要氨基酸成分的差异可能决定不同种鲑类的氨基酸行为选择偏好。马苏大马哈鱼对氨基酸比例相似的人工刺激液表现出与栖息地水溶液相似的反应[8],也可以说明这一点。本实验中,只验证了试验鱼对单个氨基酸的行为反应,对两种或多种的混和氨基酸行为反应还待进一步研究。
细鳞鲑肌肉中的谷氨酸(1.75%干重)、天冬氨酸(1.59%)、甘氨酸(0.85%)和丙氨酸(0.75%),也是其所有氨基酸中含量较高的几种[24]。每年的4-6月,性成熟的细鳞鲑有从中游溯河至上游进行生殖洄游的习性,繁殖后有部分亲鱼死于栖息地[25],单体氨基酸从鱼体释放和溶解可能是部分水体中的主要氨基酸的来源。另一方面,上游新鲜程度较高的有机物排放也影响水体中的氨基酸成分和含量变化。保护细鳞鲑野生资源的最终目标是使其能够在自然条件下繁殖,恢复群体数量[25]。在细鳞鲑生殖期,保证其上游水体以及其溯河通道水域的氨基酸成分的稳定是提高细鳞鲑溯河生殖洄游效率的重要保证。如今,人类活动的各种化学污染可能会阻断细鳞鲑的洄游路径,从而影响其自然繁殖的成功率。因此,对细鳞鲑野生资源的保护还涉及水体中氨基酸的长期监控,以及时发现和控制排放源。
