哺乳动物卵泡发育调控分子机制研究进展
2018-12-11曹俊国陈敏李文李丹丽常彤魏海军许保增
曹俊国,陈敏,李文,李丹丽,常彤,魏海军,许保增
(中国农业科学院特产研究所特种经济动物分子生物学重点实验室,长春 130112)
哺乳动物的繁殖需要雄性和雌性共同参与完成,进而将二者基因中优秀的部分传递给下一代,完成物种的繁衍。在这一过程中,哺乳动物还承担着生育后代的功能,在生殖过程中扮演着重要的角色。哺乳动物的生殖进程主要包括卵泡的生长发育、卵母细胞的成熟及排出,即雌性配子的形成、精卵融合、胚胎的形成和着床以及妊娠的维持等[1]。卵泡的生长发育对于繁殖力的高低具有重要意义,因为它可以影响排卵数及后续相关的进程。因此,深入了解哺乳动物卵泡发育的分子机制和相关信号通路的调控作用对于解决哺乳动物生殖不孕以及找出最佳的治疗方法具有重要的意义。
1 哺乳动物卵泡的发育
哺乳动物的卵泡发育过程主要包含原始卵泡、初级卵泡、次级卵泡、三级卵泡和成熟卵泡几个时期。原始卵泡是由一层扁平单层状的颗粒细胞和其所包围的未成熟的卵母细胞组成。随着原始卵泡生长启动后,卵母细胞周围的颗粒细胞由扁平单层状变成柱状,卵母细胞基因组被激活并进行转录,形成初级卵泡。当卵泡继续发育进入到次级卵泡阶段,卵泡中的颗粒细胞由单层变成多层,次级卵泡进一步发育变成三级卵泡,此时期主要变化是卵泡腔形成且卵泡腔内有卵泡液。最后卵泡发育到成熟期卵泡,该时期分为选择卵泡时期和成熟卵泡时期两个阶段。卵泡从生长到成熟期主要经历了募集、选择、优势化和闭锁几个重要环节。进入募集阶段的卵泡中会有一个卵泡定向发育成优势卵泡,同时还拥有排卵能力,这一过程被称为优势卵泡的选择。优势卵泡继续生长发育,其余卵泡则自行退化,发生闭锁。
在哺乳动物卵泡发育的各个时期,不仅在形状大小和功能上发生改变,而且也会受到一些与之相关的分子及信号通路的调控,包括雌激素(E)、卵泡刺激素(FSH)及黄体生成素(LH)等激素以及表皮生长因子(EGF)、神经生长因子(NGF)等相关细胞因子。研究发现,小鼠卵泡内颗粒细胞发生过早黄素化与卵泡内的LH水平较高相关,结果导致卵母细胞减数分裂提早发生,进而使信号转导发生变化,最终引发卵泡闭锁[2]。在大鼠的研究中发现,EGF可以促进颗粒细胞的增殖[3]。同时,激活素/抑制素作用通路、BMP/Smad信号通路、NPPC/NPR2信号通路、Nodal信号通路及FGF信号通路等也发挥着积极的作用。目前,人们对于相关的研究尚有欠缺,特别是信号通路中受体和细胞内信号分子等关键因子在哺乳动物卵泡发育不同时期颗粒细胞中的定量表达情况尚未明确,有待更进一步地深入研究。本文主要从激活素/抑制素作用通路、BMP/Smad信号通路以及NPPC/NPR2信号通路3种信号通路进行相关阐述。
2 卵泡发育调控的分子机制
2.1 激活素/抑制素信号通路
激活素(Activin)作为转化生长因子(Transforming growth factor s,TGF-s)超家族的成员,是一种主要由颗粒细胞分泌的细胞间信号分子,同时也是垂体分泌FSH的激动剂。激活素由编码的亚单位基因翻译而来[4],在结构上,激活素是同源异构二聚体的半胱氨酸结合蛋白,共享1个 亚单位。激活素类型主要包括激活素A、激活素B和激活素AB 3种。从结构上讲,激活素A和B是由二硫键相连的二聚体,由2个 亚单位构成,分别为 A和 B;而激活素 AB是 A-B异质二聚体。激活素蛋白作为一种生长因子,参与哺乳动物卵巢内多种生物学功能的改变,包括生殖细胞的生存和原始卵泡的募集、促进颗粒细胞增殖及卵泡刺激素受体(FSHR)的表达、延迟卵泡黄体化和闭锁及参与黄体溶解等[5~7]。
激活素在发挥作用时必须要有其靶细胞表面的特异性受体(Act R)的参与,与其形成三元受体复合物。激活素的受体类型主要有I型和II型丝氨酸/苏氨酸激酶2种。当信号开始转导时,激活素与II型受体结合,开始磷酸化进程,之后激活I型受体,下游的信号分子(R-Smads)磷酸化,受体Smad会与磷酸化的R-Smads结合发生位置转移,进入到细胞核内与特异性受体结合,发挥调控作用(图1)。
干细胞因子(Stem cell factor,SCF,也称为Kit配体,KL)由卵巢颗粒细胞分泌,并通过卵母细胞表面受体c-Kit与卵母细胞发挥作用[8]。SCF及c-Kit作为颗粒细胞和卵母细胞发生作用的纽带有重要的作用[9]。激活素可以通过SCF及c-Kit的纽带作用,间接促进卵母细胞发育。研究证明,大鼠卵巢内的激活素 A通过 smad2/ERK5/nur77信号通路促进颗粒细胞的增殖并提高其活性,使SCF分泌增多,然后与c-Kit发生特异性的结合,结合后的SCF/c-Kit在大鼠卵巢内的表达水平提高,从而促进卵母细胞的发育[10]。
同样作为TGF-超家族成员之一的抑制素(Inhibin)是性腺分泌的一类大分子糖蛋白质激素,其结构与激活素相似,只不过它是一种异二聚体结构,没有共享1个 亚单位,而是分别亚基和 亚基2种亚单位通过二硫键相连。抑制素的生理作用主要是与激活素一起调控垂体FSH的合成和分泌。许多研究表明,抑制素通过内分泌作用调节雌激素的分泌,进而调控垂体FSH的分泌[11,12]。该二硫键发挥着非常重要的作用,单独存在亚单位和 亚单位是没有生物活性的[13]。 亚单位有两种类型,即 A和 B,它们结构上类似,但性质上有些差异,所以,抑制素有抑制素A(基和B亚基组成)两种存在形式[14]。对于抑制素A,在卵基和 A亚基组成)和抑制素B(泡发育的不同阶段均发挥作用,但在优势卵泡发育阶段和黄体形成阶段表达较高。在优势卵泡发育阶段,抑制素A的浓度增加,加大了优势卵泡对FSH的敏感性,防止优势卵泡发生闭锁;在黄体形成阶段,抑制素A的主要作用是促进卵泡黄体化,抑制素B主要在中、小窦卵泡中表达,并在早期卵泡发育阶段发挥重要作用。抑制素B可以强化FSH抑制非优势卵泡进入排卵前期,有利于优势卵泡筛选[15],此外,低浓度的抑制素B与生殖机能老化也有关,如卵泡发育异常及卵泡囊肿等。研究发现,多囊卵巢综合症患者卵泡液中抑制素B水平显著下降[16]。因此,抑制素的作用对于调节颗粒细胞的生长、发育以及调控卵母细胞的成熟方式等,最终调节卵泡的发育发挥着重要的作用[17]。
抑制素它在阻碍激活素的作用通路方面效果显著,抑制素通过 亚基竞争性与激活素II型受体结合,导致激活素I型受体磷酸化受阻,进而R-Smads激活也被阻断,起到一种拮抗的效果。Tgfbr3是TGF-超家族的受体,也被称作是Betaglycan,现在被认为是抑制素的共受体[18],它与抑制素结合能增强抑制素与激活素II型受体结合,从而增强抑制素阻断激活素作用通路[19]。
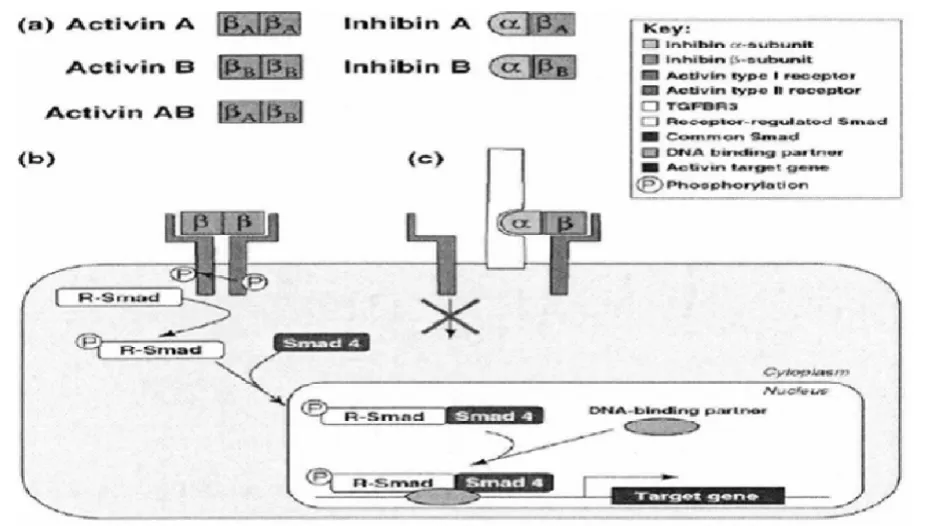
图1 激活素/抑制素作用信号通路[20]Figure1 Activins and inhibins and their mechanism of signaling pathway
2.2 BMP/Smad信号通路
骨形态发生蛋白(Bone morphogenetic protein,Bmps)是一种与骨及神经组织等形成有关的蛋白,它可以诱导未分化的间充质定向分化成骨细胞,骨细胞进一步分化成钙化骨组织[21]。Bmps是TGF-超家族中一个重要成员,对于哺乳动物骨骼形成及发育等有重要的作用。合成Bmps过程中,首先合成的是前体,前体蛋白主要包括N端引导序列(也称为信号肽)、C端成熟区以及中间区域(也称为前肽)3个区域。其中,C端成熟区参与多肽链内部与肽链间二硫键的形成,由其所含的保守半胱氨酸执行,并将2个多肽链合成未成熟的二聚体结构,然后在蛋白水解酶的作用下形成具有活性的成熟二聚体,再与对应的受体结合来发挥作用[22]。
哺乳动物的Bmps受体是一种膜蛋白受体,其分子结构主要有细胞外区、内区以及中间部分的跨膜区等几个部分。目前,我们所知道的主要有2种,分别为 I型和 II型受体。其中,I型受体主要包括 I A(BMPR-A 或ALK-3)和 I B(BMPR-IB或ALK-6)2个亚型;II型受体有5种,分别是BMPR-II、ActR-IIA、ActR-IIB、AMHR-II和 T13R-II[23]。Bmp信号通路就是通过配体与这2种受体结合形成异源复合物,进而发挥其生物学功能[24]。
Bmps家族的信号转导主要分为2种途径,分别为Smads依赖型通路和磷脂酰肌醇3激酶(Phosphoinositide protein kinase PI3K)等非Smads依赖型通路。对于Smads家族,最初是从果蝇和线虫的相关蛋白中提取得到的,Smads家族的每个成员在信号转导通路中行使的功能会有所不同,大概可分为3种类型:受体调节型 Smads,包括 Smad1、Smad2、Smad3、Smad5、Smad8和Smad9;共同介导型Smads,包括Smad4;抑制型Smads,包括Smad6、Smad7,且它们主要起拮抗因子的作用[25]。哺乳动物卵泡的生长发育调控机制主要受到上述Smads依赖性通路的调控,即Bmps/Smads信号通路。
Bmps家族成员及相关信号转导对于哺乳动物卵泡的生长、颗粒细胞生长分化以及卵母细胞成熟、排卵等有重要的调控作用。对于 Bmps/Smads信号通路,其信号转导模式为Bmps与细胞膜上的BMPR-II受体结合,进而使其发生磷酸化过程,磷酸化的BMPR-II受体与 BMP-I型受体结合形成复合物,BMP-I型受体被相应的蛋白激酶激活发生磷酸化过程,然后激活Smads信号分子,被激活的R-Smads(主要有 Smadl、Smad5、Smad8)结合共用型的 CoSmad4共同形成Smad蛋白复合物,进入到细胞核与特异的DNA序列结合,启动下游靶基因的启动子,使下游基因开始进行转录[26~28],下游信号分子 R-Smads在BMP/Smad信号通路中也占有重要地位。敲除Smad4后的小鼠,类固醇类激素调节被阻断,血浆孕酮水平上升,颗粒细胞发生早熟性黄体化,最终导致卵巢功能早衰[29]。所以,进一步认识BMP/Smad信号通路的调控机理对于解决哺乳动物卵泡发育机制及生殖不孕等关键问题具有深远意义。
2.3 NPPC/NPR2信号通路
钠尿肽家族广泛存在于动物大脑、心脏等各类组织和器官中,具有维持血压及血容量的稳定、促进脂肪代谢和软骨生长等多个功能[30]。该家族在哺乳动物中主要由3种配体和3种特异性受体组成。配体以前体肽形式存在,分别是心房钠尿肽(Atrial natriuretic peptide,ANP,又称为NPPA)、脑钠尿肽(Brain natriuretic peptide,BNP,又称为NPPB)、C-型钠肽(C-type natriuretic peptide,CNP,又称为 NPPC);特异性受体以二聚体形式存在,分别是钠尿肽受体A(Natriuretic peptide receptor A,NPRA,又称为 NPR1)、钠尿肽受体 B(Natriuretic peptide receptor B,NPRB,又称为 NPR2)、钠尿肽受体 C(Natriuretic peptide receptor C,NPRC,又称为 NPR3)[31]。NPPA 和 NPPB 在哺乳动物心脏内合成,主要是通过与血液中的肾素-血管紧张素-醛固酮系统拮抗来达到保持体内电解质的平衡,进而使心血管内环境稳定[32];NPPC是一种有生物活性的小分子多肽,由22个氨基酸构成,所编码的基因具有高度的保守性,在哺乳动物卵母细胞减数分裂方面具有重要作用,它与NPR2有较好的亲和性,可以与其特异性结合而发挥作用。3种钠尿肽受体在哺乳动物中均有1个细胞外配体结合域和1个单跨膜结构,其中,NPR1和NPR2同为鸟苷酸环化酶偶联受体,催化GTP生成细胞内第二信使,进而介导利钠肽发挥作用;NPR3为钠肽清除受体,与利钠肽的清除有关[33]。
NPPC/NPR2信号通路在抑制哺乳动物卵母细胞过早成熟方面发挥着巨大的作用。卵泡壁层颗粒细胞产生的 NPPC与 NPR2结合刺激产生了 cGMP,cGMP通过卵母细胞和颗粒细胞的间隙链接进入到卵母细胞中[34],使磷酸二酯酶(Phosphodiesterase,PDE3A)的活性被抑制,cAMP的水解程度也随之下降,从而稳定在一个较高的水平,cAMP依赖的蛋白激酶PKA通过磷酸酶细胞周期蛋白(Phosphatase cell division cycle 25,CDC25)、weel激酶和髓磷脂转录因子Myt1,调节促成熟因子(Maturation-promoting factor,MPF)的活性,CDC25脱磷酸化周期蛋白依赖性激酶 1(Cyclin-dependent kinase 1,CDK1),weel和Myt1磷酸化CDK1,磷酸化的CDK1以及相关复合体失活,最终抑制了卵母细胞的成熟(图2)。
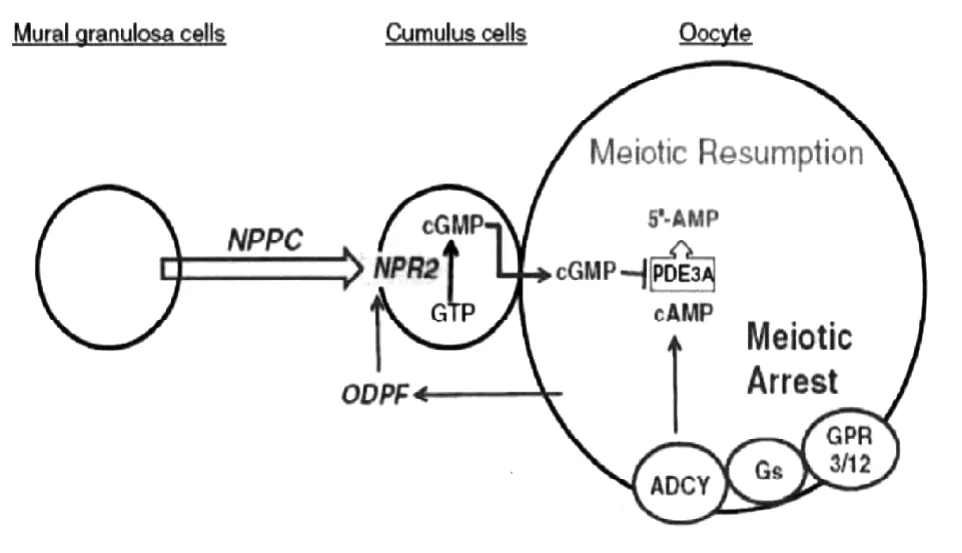
图2 NPPC及NPR2抑制卵母细胞成熟的分子机制[35]Figure 2 Mechanism of NPPC and NPR2 in the maintenance of oocyte meiotic arrest
3 展望
如今,哺乳动物卵泡发育分子机制的相关研究已取得了巨大的成果。目前,已经证明了许多信号通路在卵泡生长发育过程中起着非常重要的作用,并且其中一些信号通路的研究已经非常透彻。但是,还有一些信号通路的研究,如哺乳动物是如何启动原始卵泡发育以及在信号转导途径中又有哪些下游靶基因进行了参与等相关的问题仍未解决。因此,深入了解哺乳动物卵泡发育分子机制的调控仍然需要对卵泡发育相关生物学及有关基因表达继续研究,这对于治疗哺乳动物生殖不孕等疾病有重要意义。
